
Appropriate Human Serum Albumin Fluid Therapy and the Alleviation of COVID-19 Vulnerabilities: An Explanation of the HSA Lymphatic Nutrient Pump


Abstract
:1. Introduction

2. Long COVID-19
Evidence That Low HSA Is Responsible for COVID-19 and Long-COVID-19 Vulnerabilities
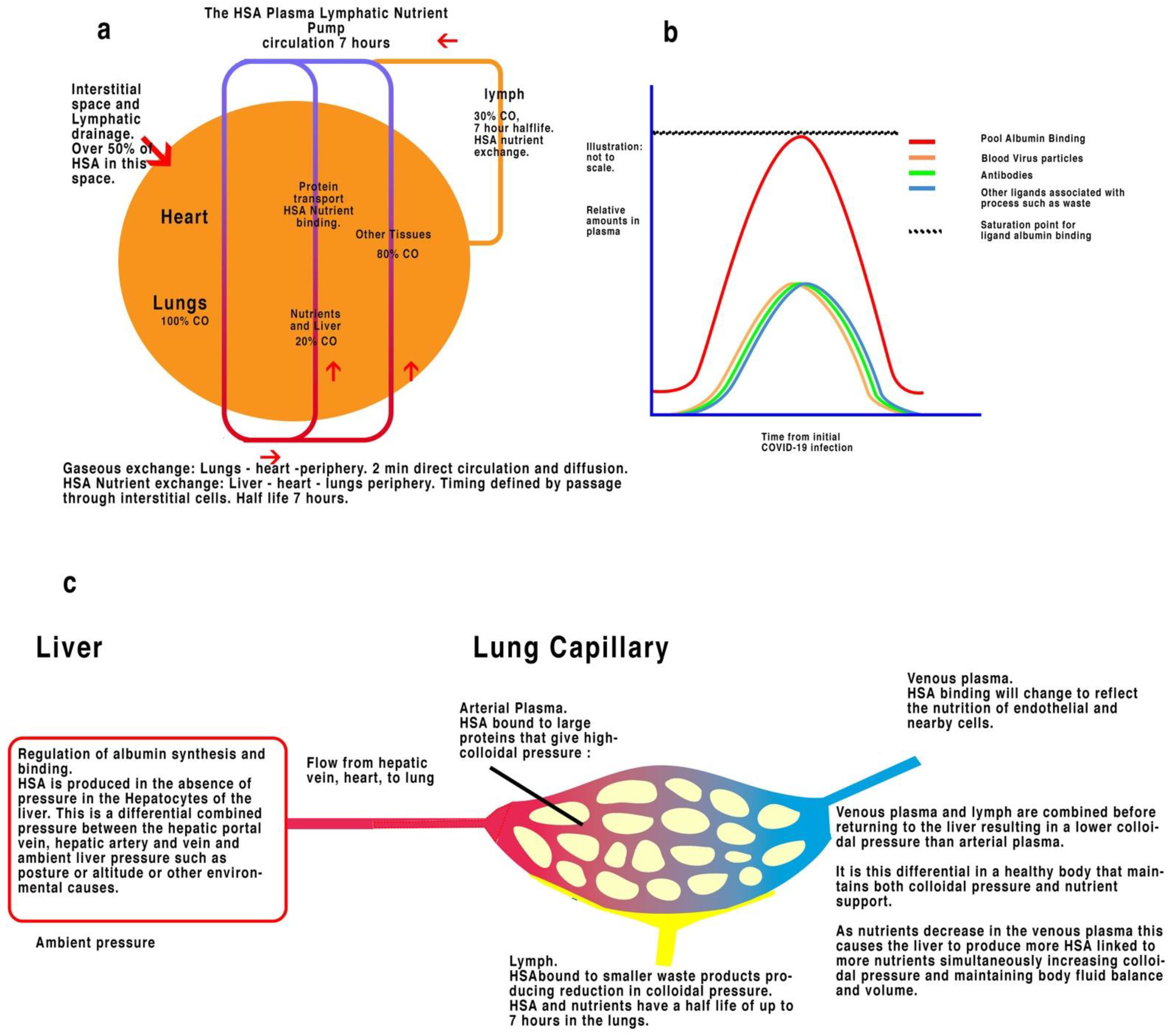
3. Vaccines Revisited
4. Generation of Vulnerabilities That Spread Systemically to Cause Complications
4.1. Albumin-Binding Deficiency
4.2. Common Damage to Epithelial Cells
4.3. The Lymphatic System and Plasma–Lymph Nutrient–Albumin Pump
4.4. Liver
4.5. Obesity
4.6. Diabetes
4.7. Arthritic Pain
4.8. Lungs
4.9. Heart
4.10. The Blood Brain, Placental Barriers, and Albumin Transport in the Kidney
4.11. The Central Nervous System and the Blood–Brain Barrier (BBB)
- (I)
- Virions attached to HSA may affect the transport of vital nutrients across the BBB. This nutrient deficit will depend upon the state of has-binding deficiency. This level of binding deficiency will alter the levels of transmitter and affect transmission of action potential affecting cognition.
- (II)
- Depletion of nutrients will also affect the capillaries of the brain, for example, the endothelial glycocalyx layer (EGL) already described [3,5]. A reduction in the EGL will eventually cause leakage, thrombosis [40], and rupture. Rupture may promote secondary infection, leading to symptoms of meningitis [41].
4.12. Kidney
4.13. Pregnancy
4.14. Skin: Distribution of Albumin in the Adult and Child and Infant Body
4.15. Fluid Therapy
5. Discussion
5.1. Physiology
5.2. Why Is Present Fluid Therapy Inappropriate in COVID-19?
5.3. Saline
5.4. HSA Infusion
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Moman, R.N.; Gupta, N.; Varacallo, M. Physiology, Albumin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK459198/ (accessed on 4 January 2022).
- Sherwood, L. Human Physiology: From Cells to Systems, 8th ed.; Brooks/Cole, Cengage Learning: Boston, MA, USA, 2013; ISBN 9781111577438. (In English) [Google Scholar]
- Johnson, A.S.; Winlow, W. COVID-19 vulnerabilities are intensified by declining human serum albumin levels. Exp. Physiol. 2022, 107, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Swartz, M.A. The physiology of the lymphatic system. Adv. Drug Deliv. Rev. 2001, 50, 3–20. [Google Scholar] [CrossRef]
- Johnson, A.S.; Fatemi, R.; Winlow, W. SARS-CoV-2 Bound Human Serum Albumin and Systemic Septic Shock. Front. Cardiovasc. Med. 2020, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Paar, M.; Rossmann, C.; Nusshold, C.; Wagner, T.; Schlagenhauf, A.; Leschnik, B.; Oettl, K.; Koestenberger, M.; Cvirn, G.; Hallström, S. Anticoagulant action of low, physiologic, and high albumin levels in whole blood. PLoS ONE 2017, 12, e0182997. [Google Scholar] [CrossRef]
- Gyamlani, G.; Molnar, M.; Lu, J.; Sumida, K.; Kalantar-Zadeh, K.; Kovesdy, C. Association of serum albumin level and venous thromboembolic events in a large cohort of patients with nephrotic syndrome. Nephrol. Dial. Transplant. 2017, 32, 157–164. [Google Scholar] [CrossRef]
- Goncharov, N.V.; Belinskaya, D.A.; Razygraev, A. On the enzymatic activity of albumin. Russ. J. Bioorg. Chem. 2015, 41, 113–124. [Google Scholar] [CrossRef]
- Weaving, G.; Batstone, G.F.; Jones, R.G. Age and sex variation in serum albumin concentration: An observational study. Ann. Clin. Biochem. 2016, 53, 106–111. [Google Scholar] [CrossRef]
- Islam, N.; Shkolnikov, V.M.; Acosta, R.J.; Klimkin, I.; Kawachi, I.; Irizarry, R.A.; Alicandro, G.; Khunti, K.; Yates, T.; Jdanov, D.A.; et al. Excess deaths associated with COVID-19 pandemic in 2020: Age and sex disaggregated time series analysis in 29 high income countries. BMJ 2021, 373, n1137. [Google Scholar] [CrossRef]
- Egesten, A.; Frick, I.M.; Mörgelin, M.; Olin, A.I.; Björck, L. Binding of albumin promotes bacterial survival at the epithelial surface. J. Biol. Chem. 2011, 286, 2469–2476. [Google Scholar] [CrossRef]
- De, R.; Dutta, S. Role of the Microbiome in the Pathogenesis of COVID-19. Front. Cell. Infect. Microbiol. 2022, 12, 736397. [Google Scholar] [CrossRef]
- Altmann, D.M.; Boyton, R.J. COVID-19 vaccination: The road ahead. Science 2022, 375, 1127–1132. [Google Scholar] [CrossRef]
- Yamaoka-Tojo, M. Endothelial glycocalyx damage as a systemic inflammatory microvascular endotheliopathy in COVID-19. Biomed. J. 2020, 43, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Klammt, S.; Wojak, H.J.; Mitzner, A.; Koball, S.; Rychly, J.; Reisinger, E.; Mitzner, S. Albumin-binding capacity (ABiC) is reduced in patients with chronic kidney disease along with an accumulation of protein-bound uraemic toxins. Nephrol. Dial. Transplant. 2012, 27, 2377–2383. [Google Scholar] [CrossRef]
- Oettl, K.; Birner-Gruenberger, R.; Spindelboeck, W.; Stueger, H.P.; Dorn, L.; Stadlbauer, V.; Putz-Bankuti, C.; Krisper, P.; Graziadei, I.; Vogel, W.; et al. Oxidative albumin damage in chronic liver failure: Relation to albumin binding capacity, liver dysfunction and survival. J. Hepatol. 2013, 59, 978–983. [Google Scholar] [CrossRef] [PubMed]
- Belmin, J.; Corman, B.; Merval, R.; Tedgui, A. Age-related changes in endothelial permeability and distribution volume of albumin in rat aorta. Am. J. Physiol. 1993, 264 Pt 2, H679–H685. [Google Scholar] [CrossRef] [PubMed]
- Shehadeh, L.A.; Webster, K.A.; Hare, J.M.; Vazquez-Padron, R.I. Dynamic regulation of vascular myosin light chain (MYL9) with injury and aging. PLoS ONE 2011, 6, e25855. [Google Scholar] [CrossRef]
- Du Preez, H.N.; Aldous, C.; Kruger, H.G.; Lin, J. N-acetylcysteine and other sulfur-donors as a preventative and adjunct therapy for COVID-19. Adv. Pharmacol. Pharm. Sci. J. 2022; preprint. [Google Scholar] [CrossRef]
- Du Preez, H.N.; Aldous, C.; Hayden, M.R.; Kruger, H.G.; Lin, J. Pathogenesis of COVID-19 described through the lens of an undersulfated and degraded epithelial and endothelial glycocalyx. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2022, 36, e22052. [Google Scholar] [CrossRef]
- Curry, F.E.; Michel, C.C. The Colloid Osmotic Pressure Across the Glycocalyx: Role of Interstitial Fluid Sub-Compartments in Trans-Vascular Fluid Exchange in Skeletal Muscle. Front. Cell Dev. Biol. 2021, 9, 729873. [Google Scholar] [CrossRef]
- Ridruejo, E.; Soza, A. The liver in times of COVID-19: What hepatologists should know. Ann. Hepatol. 2020, 19, 353–358. [Google Scholar] [CrossRef]
- Hirt, P.A.; Castillo, D.E.; Yosipovitch, G.; Keri, J.E. Skin changes in the obese patient. J. Am. Acad. Dermatol. 2019, 81, 1037–1057. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.C.; Xu, X.; Ferrante, A.W.; Krakoff, J. Reduced plasma albumin predicts type 2 diabetes and is associated with greater adipose tissue macrophage content and activation. Diabetol. Metab. Syndr. 2019, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Anguizola, J.; Matsuda, R.; Barnaby, O.S.; Hoy, K.S.; Wa, C.; DeBolt, E.; Koke, M.; Hage, D.S. Glycation of human serum albumin. Clin. Chim. Acta Int. J. Clin. Chem. 2013, 425, 64–76. [Google Scholar] [CrossRef]
- Shiraki, T.; Miura, Y.; Sawada, T. Glycated albumin suppresses glucose-induced insulin secretion by impairing glucose metabolism in rat pancreatic β-cells. Nutr. Metab. 2011, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.M.; Song, S.O.; You, Y.-H.; Yoon, K.-H.; Kang, E.S.; Cha, B.S.; Lee, H.C.; Kim, J.-W.; Lee, B.-W. Glycated albumin causes pancreatic β-cells dysfunction through autophagy dysfunction. Endocrinology 2013, 154, 2626–2639. [Google Scholar] [CrossRef] [PubMed]
- Iles, J.K.; Zmuidinaite, R.; Sadee, C.; Gardiner, A.E.; Lacey, J.C.; Harding, S.; Ule, J.; Roblett, D.; Heeney, J.L.; Baxendale, H.; et al. SARS-CoV-2 Spike Protein Binding of Glycated Serum Albumin—Its Potential Role in the Pathogenesis of the COVID-19 Clinical Syndromes and Bias towards Individuals with Pre-Diabetes/Type 2 Diabetes and Metabolic Diseases. Int. J. Mol. Sci. 2022, 23, 4126. [Google Scholar] [CrossRef]
- Taldaev, A.; Rudnev, V.; Kulikova, L.; Nikolsky, K.; Efimov, A.; Malsagova, K.; Kaysheva, A. Molecular Dynamics Study of Citrullinated Proteins Associated with the Development of Rheumatoid Arthritis. Proteomes 2022, 10, 8. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, L.; He, H.; Wei, L.; Lai, W.; Yuan, J.; Hong, X.; Liu, L.; Wang, B.; Nandakumar, K.S.; et al. Albumin/Globulin Ratio as Yin-Yang in Rheumatoid Arthritis and Its Correlation to Inflamm-Aging Cytokines. J. Inflamm. Res. 2021, 14, 5501–5511. [Google Scholar] [CrossRef]
- Katsoularis, I.; Fonseca-RodrÃguez, O.; Farrington, P.; Jerndal, H.; Lundevaller, E.H.; Sund, M. Risks of deep vein thrombosis, pulmonary embolism, and bleeding after covid-19: Nationwide self-controlled cases series and matched cohort study. BMJ 2022, 377, e069590. [Google Scholar] [CrossRef]
- Asakura, H.; Ogawa, H. COVID-19-associated coagulopathy and disseminated intravascular coagulation. Int. J. Hematol. 2021, 113, 45–57. [Google Scholar] [CrossRef]
- Sidik, S.M. Heart Disease after COVID: What the Data Say. Nature 2022, 608, 26–28. [Google Scholar] [CrossRef] [PubMed]
- Lambot, N.; Lybaert, P.; Boom, A.; Delogne-Desnoeck, J.; Vanbellinghen, A.M.; Graff, G.; Lebrun, P.; Meuris, S. Evidence for a clathrin-mediated recycling of albumin in human term placenta. Biol. Reprod. 2006, 75, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Kaksonen, M.; Roux, A. Mechanisms of clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 2018, 19, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Hampshire, A.; Chatfield, D.A.; MPhil, A.M.; Jolly, A.; Trender, W.; Hellyer, P.J.; Giovane, M.D.; Newcombe, V.; Outtrim, J.G.; Warne, B.; et al. Cambridge NeuroCOVID Group, the NIHR COVID-19 BioResource, and Cambridge NIHR Clinical Research Facility (2022). Multivariate profile and acute-phase correlates of cognitive deficits in a COVID-19 hospitalised cohort. EClinicalMedicine 2022, 47, 101417. [Google Scholar] [CrossRef]
- Stanyon, H.F.; Viles, J.H. Human serum albumin can regulate amyloid-β peptide fiber growth in the brain interstitium: Implications for Alzheimer disease. J. Biol. Chem. 2012, 287, 28163–28168. [Google Scholar] [CrossRef]
- Zhao, M.; Guo, C. Multipronged Regulatory Functions of Serum Albumin in Early Stages of Amyloid-β Aggregation. ACS Chem. Neurosci. 2021, 12, 2409–2420. [Google Scholar] [CrossRef]
- Ziff, O.J.; Ashton, N.J.; Mehta, P.R.; Brown, R.; Athauda, D.; Heaney, J.; Heslegrave, A.J.; Benedet, A.L.; Blennow, K.; Checkley, A.M.; et al. Amyloid processing in COVID-19-associated neurological syndromes. J. Neurochem. 2022, 161, 146–157. [Google Scholar] [CrossRef]
- Gorog, D.A.; Storey, R.F.; Gurbel, P.A. Current and novel biomarkers of thrombotic risk in COVID-19: A Consensus Statement from the International COVID-19 Thrombosis Biomarkers Colloquium. Nat. Rev. Cardiol. 2022, 19, 475–495. [Google Scholar] [CrossRef] [PubMed]
- Mondal, R.; Ganguly, U.; Deb, S.; Shome, G.; Pramanik, S.; Bandyopadhyay, D.; Lahiri, D. Meningoencephalitis associated with COVID-19: A systematic review. J. Neurovirology 2021, 27, 12–25. [Google Scholar] [CrossRef]
- Wu, H.L.; Shenoy, M.; Kalra, P.A.; Chinnadurai, R. Intrinsic Kidney Pathology Following COVID-19 Infection in Children and Adolescents: A Systematic Review. Children 2022, 9, 3. [Google Scholar] [CrossRef]
- Birn, H.; Christensen, E.I. Renal albumin absorption in physiology and pathology. Kidney Int. 2006, 69, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Castrop, H.; Schießl, I.M. Novel routes of albumin passage across the glomerular filtration barrier. Acta Physiol. 2017, 219, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Pollak, M.R.; Quaggin, S.E.; Hoenig, M.P.; Dworkin, L.D. The glomerulus: The sphere of influence. Clin. J. Am. Soc. Nephrol. CJASN 2014, 9, 1461–1469. [Google Scholar] [CrossRef] [PubMed]
- Muner, M.B.; Velez, J. Proteinuria in COVID-19. Clin. Kidney J. 2021, 14 (Suppl. 1), i40–i47. [Google Scholar] [CrossRef]
- Li, F.; Guo, H.; Zou, J.; Chen, W.; Lu, Y.; Zhang, X.; Fu, C.; Xiao, J.; Ye, Z. Urinary excretion of uric acid is negatively associated with albuminuria in patients with chronic kidney disease: A cross-sectional study. BMC Nephrol. 2018, 19, 95. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.; Winlow, W. Pre-Eclampsia, Hypoalbuminaemia and Albumin Therapy. Eur. J. Biomed. Pharm. Sci. 2021, 8, 75–78. [Google Scholar]
- Agnihothri, R.; Fox, L.P. Clinical Patterns and Morphology of COVID-19 Dermatology. Dermatol. Clin. 2021, 39, 487–503. [Google Scholar] [CrossRef]
- Baeck, M.; Hoton, D.; Marot, L.; Herman, A. Chilblains and COVID-19: Why SARS-CoV-2 endothelial infection is questioned. Br. J. Dermatol. 2020, 183, 1152–1153. [Google Scholar] [CrossRef]
- Colmenero, I.; Santonja, C.; Alonso-Riaño, M.; Noguera-Morel, L.; Hernández-Martín, A.; Andina, D.; Wiesner, T.; Rodríguez-Peralto, J.L.; Requena, L.; Torrelo, A. SARS-CoV-2 endothelial infection causes COVID-19 chilblains: Histopathological, immunohistochemical and ultrastructural study of seven paediatric cases. Br. J. Dermatol. 2021, 183, 729–737. [Google Scholar] [CrossRef]
- Genovese, G.; Moltrasio, C.; Berti, E.; Marzano, A.V. Skin Manifestations Associated with COVID-19: Current Knowledge and Future Perspectives. Dermatology 2021, 237, 1–12. [Google Scholar] [CrossRef]
- Rekhtman, S.; Tannenbaum, R.; Strunk, A.; Birabaharan, M.; Wright, S.; Garg, A. Mucocutaneous disease and related clinical characteristics in hospitalized children and adolescents with COVID-19 and multisystem inflammatory syndrome in children. J. Am. Acad. Dermatol. 2021, 84, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.A.; Karmakar, S.; Chakraborty, U.; Sil, A.; Chandra, A. Purpura fulminans as the presenting manifestation of COVID-19. Postgrad. Med. J. 2021, 97, 473. [Google Scholar] [CrossRef] [PubMed]
- Hirschler, V. Skin and obesity in childhood: An update. AIMS Med. Sci. 2021, 8, 311–323. [Google Scholar] [CrossRef]
- Hoste, L.; Van Paemel, R.; Haerynck, F. Multisystem inflammatory syndrome in children related to COVID-19: A systematic review. Eur. J. Pediatr. 2021, 180, 2019–2034. [Google Scholar] [CrossRef]
- Fareau, G.G.; Vassilopoulou-Sellin, R. Hypercortisolemia and infection. Infect. Dis. Clin. N. Am. 2007, 21, 639–657. [Google Scholar] [CrossRef]
- Dziedzic, T.; Pera, J.; Wnuk, M.; Szczudlik, A.; Slowik, A. Serum albumin as a determinant of cortisol release in patients with acute ischemic stroke. Atherosclerosis 2012, 221, 212–214. [Google Scholar] [CrossRef]
- Tan, T.; Khoo, B.; Mills, E.G.; Phylactou, M.; Patel, B.; Eng, P.C.; Thurston, L.; Muzi, B.; Meeran, K.; Prevost, A.T.; et al. Association between high serum total cortisol concentrations and mortality from COVID-19. Lancet Diabetes Endocrinol. 2020, 8, 659–660. [Google Scholar] [CrossRef]
- Sakthivadivel, V.; Bohra, G.K.; Maithilikarpagaselvi, N.; Khichar, S.; Meena, M.; Palanisamy, N.; Gaur, A.; Garg, M.K. Association of Inflammatory Markers with COVID-19 Outcome among Hospitalized Patients: Experience from a Tertiary Healthcare Center in Western India. Maedica 2021, 16, 620–627. [Google Scholar] [CrossRef]
- Chen, C.-H.; Lin, S.-W.; Shen, C.-F.; Hsieh, K.-S.; Cheng, C.-M. Biomarkers during COVID-19: Mechanisms of Change and Implications for Patient Outcomes. Diagnostics 2022, 12, 509. [Google Scholar] [CrossRef]
- Finfer, S.; Myburgh, J.; Bellomo, R. Intravenous fluid therapy in critically ill adults. Nat. Rev. Nephrol. 2018, 14, 541–557. [Google Scholar] [CrossRef]
- Dubois, M.J.; Orellana-Jimenez, C.; Melot, C.; De Backer, D.; Berre, J.; Leeman, M.; Brimioulle, S.; Appoloni, O.; Creteur, J.; Vincent, J.L. Albumin administration improves organ function in critically ill hypoalbuminemic patients: A prospective, randomized, controlled, pilot study. Crit. Care Med. 2006, 34, 2536–2540. [Google Scholar] [CrossRef] [PubMed]
- Caironi, P.; Tognoni, G.; Masson, S.; Fumagalli, R.; Pesenti, A.; Romero, M.; Fanizza, C.; Caspani, L.; Faenza, S.; Grasselli, G.; et al. Albumin replacement in patients with severe sepsis or septic shock. N. Engl. J. Med. 2014, 370, 1412–1421. [Google Scholar] [CrossRef] [PubMed]
- Ramadori, G. Albumin Infusion in Critically Ill COVID-19 Patients: Hemodilution and Anticoagulation. Int. J. Mol. Sci. 2021, 22, 7126. [Google Scholar] [CrossRef]
- Imoberdorf, R.; Garlick, P.J.; McNurlan, M.A.; Casella, G.A.; Peheim, E.; Turgay, M.; Bärtsch, P.; Ballmer, P.E. Enhanced synthesis of albumin and fibrinogen at high altitude. J. Appl. Physiol. 2001, 2, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Ramadori, G. Hypoalbuminemia: An underestimated, vital characteristic of hospitalized COVID-19 positive patients? Hepatoma Res. 2020, 6, 28. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| There is repeated evidence of a connection between hypoalbuminemia and COVID-19 for each symptom of and vulnerability to COVID-19. HSA binding deficiency is a common factor. |
| There is evidence that raising HSA concentration in the liver may alleviate some of the vulnerabilities to COVID-19 by reducing any HSA binding deficiencies. |
| A mechanism for albumin involvement in long COVID-19 also exists and could be removed by appropriate HSA therapy, given that the liver precisely modulates nutrients in the plasma and maintains HSA levels. |
| Present fluid therapy, either saline or colloid, applied to a peripheral vein, results in a destabilisation of nutrient transport, leading to nutrient deficits in cells and cellular components because of albumin-binding deficiency. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, A.S.; Polese, G.; Johnson, M.; Winlow, W. Appropriate Human Serum Albumin Fluid Therapy and the Alleviation of COVID-19 Vulnerabilities: An Explanation of the HSA Lymphatic Nutrient Pump. COVID 2022, 2, 1379-1395. https://doi.org/10.3390/covid2100099
Johnson AS, Polese G, Johnson M, Winlow W. Appropriate Human Serum Albumin Fluid Therapy and the Alleviation of COVID-19 Vulnerabilities: An Explanation of the HSA Lymphatic Nutrient Pump. COVID. 2022; 2(10):1379-1395. https://doi.org/10.3390/covid2100099
Chicago/Turabian StyleJohnson, Andrew S., Gianluca Polese, Max Johnson, and William Winlow. 2022. "Appropriate Human Serum Albumin Fluid Therapy and the Alleviation of COVID-19 Vulnerabilities: An Explanation of the HSA Lymphatic Nutrient Pump" COVID 2, no. 10: 1379-1395. https://doi.org/10.3390/covid2100099
APA StyleJohnson, A. S., Polese, G., Johnson, M., & Winlow, W. (2022). Appropriate Human Serum Albumin Fluid Therapy and the Alleviation of COVID-19 Vulnerabilities: An Explanation of the HSA Lymphatic Nutrient Pump. COVID, 2(10), 1379-1395. https://doi.org/10.3390/covid2100099