Abstract
Human granulocyte-macrophage colony-stimulating factor (GM-CSF) is an important therapeutic cytokine. Methylotrophic yeasts such as Komagataella phaffii and Ogataea parapolymorpha are attractive hosts for recombinant protein production. In this study, these yeasts were engineered to produce GM-CSF in both secreted and cell-surface-anchored forms. Secreted GM-CSF accumulated to tens of milligrams per liter in culture supernatants following induction. Fluorescent antibody labeling confirmed that yeast strains expressing surface-displayed GM-CSF exhibited markedly increased fluorescence compared to parental strains. The highest signal was observed in K. phaffii and further validated by immunofluorescence microscopy. Functional assays demonstrated that K. phaffii cells displaying GM-CSF stimulated TF-1 cell proliferation 1.41-fold compared to control yeast lacking GM-CSF. These results confirm both the successful surface display and preserved biological activity of the cytokine. This work demonstrates the feasibility of engineering K. phaffii to present active human GM-CSF on the cell surface. Surface display was efficient and yielded biologically functional cytokine, as shown by fluorescence-based quantification and TF-1 proliferation assays. The study highlights yeast surface display as a promising platform for delivering therapeutic cytokines without requiring purification steps.
1. Introduction
The development of innovative and effective strategies to enhance wound healing and tissue regeneration remains a pressing challenge in modern medicine. Wound healing is a complex, highly regulated process involving numerous cell types and molecular signals. Polypeptide growth factors are central to this process, modulating key pathways such as PI3K/AKT, RAS/MAPK, and JAK/STAT to promote cell proliferation, intercellular communication, extracellular matrix formation, angiogenesis, epithelialization, and granulation tissue development [1,2]. However, the bioavailability of these factors at wound sites is often limited due to reduced synthesis or rapid degradation, especially in chronic or severe wounds [3].
Granulocyte-macrophage colony-stimulating factor (GM-CSF) is a critical cytokine in wound healing and immune modulation, detected in wound beds following acute injury [4]. GM-CSF exerts diverse effects, including enhancing myofibroblast differentiation and wound contraction, recruiting inflammatory cells, promoting epidermal proliferation, and facilitating Langerhans cell migration [5,6,7,8]. Beyond wound repair, GM-CSF stimulates the proliferation and differentiation of hematopoietic stem cells, underscoring its role as a potent immunostimulant [9]. Studies in GM-CSF-deficient animal models reveal impaired healing, reduced immune cell recruitment, and diminished vascularization [10], while local GM-CSF application significantly accelerates recovery [11,12]. Despite its therapeutic promise, the high production costs and limitations of current expression systems restrict GM-CSF’s broader application, particularly for chronic or combat-related wounds.
Recombinant GM-CSF has been produced in various platforms, including mammalian cells, bacteria, yeasts, plant cells, and baculovirus systems [13,14,15,16]. However, these systems often yield GM-CSF with deviations from the native human protein, such as altered glycosylation, amino acid substitutions, or stability issues. Commercially available GM-CSF from Saccharomyces cerevisiae is glycosylated but features a leucine-to-proline substitution at position 23, while the Escherichia coli-derived version is non-glycosylated with an additional N-terminal methionine [17]. Bacterial systems face challenges like inclusion body formation, incomplete methionine cleavage, and misfolding, potentially increasing immunogenicity [18,19,20]. Mammalian systems, though capable of proper glycosylation, are hampered by GM-CSF cytotoxicity to host cells and contamination with endogenous factors [21].
Yeast-based expression systems offer a compelling alternative for cytokine production, combining eukaryotic post-translational modifications with rapid growth on cost-effective media and thereby reducing production costs. Promising results have been reported with Yarrowia lipolytica, where secretion of GM-CSF under the control of the POX2 promoter yielded up to 250 mg/L of the target cytokine in bioreactor cultures, and the purified protein demonstrated clear biological activity [22]. Comparative studies of secreted GM-CSF expression in Komagataella phaffii (formerly Pichia pastoris) and Saccharomyces cerevisiae showed markedly higher productivity in K. phaffii (285 mg/L versus 64 mg/L), and the protein purified from K. phaffii displayed substantially greater biological activity on the GM-CSF-dependent TF-1 cell line than that obtained from S. cerevisiae [23]. In addition, strains of K. phaffii have been engineered to secrete either recombinant human GM-CSF (100 mg/L) or a chimeric GM-CSF fused to human apolipoprotein A-I (60 mg/L) in flask cultures, with both forms retaining biological activity in cell-based assays [24]. Furthermore, in bioreactor cultivation, a K. phaffii strain expressing GM-CSF under the constitutive GAP promoter produced up to 250 mg/L of the target cytokine in the culture supernatant [25]. Methylotrophic yeasts are particularly attractive for GM-CSF expression because they possess the methanol-inducible AOX1 promoter, one of the strongest eukaryotic promoters, together with a well-established molecular toolkit including synthetic promoters, CRISPR/Cas9 systems, and efficient secretion pathways [26,27]. Moreover, K. phaffii in particular can achieve very high cell densities in bioreactors, often exceeding 100 g dry cell weight per liter, which provides a considerable advantage in terms of yield and scalability [28]. A limitation of yeast systems is hyperglycosylation, which may alter protein function or immunogenicity [29]. Glycoengineering has addressed this by humanizing glycosylation patterns, enhancing therapeutic protein production. In particular, humanized strains of S. cerevisiae [30], K. phaffii [https://pichia.com/try-pichia/try-pichia-glycoswitch/ (accessed on 14 September 2025)], and O. parapolymorpha [31] have been constructed.
We utilized methylotrophic yeasts K. phaffii and O. parapolymorpha, both engineered for human-like glycosylation, to produce recombinant GM-CSF. A key novelty of this work is the first reported use of O. parapolymorpha for GM-CSF expression, along with the development of yeast strains capable of displaying GM-CSF on the cell surface as a cost-effective alternative to purified protein formulations. Nucleotide sequences encoding GM-CSF were designed, and expression vectors were constructed for both secreted and surface-anchored forms. Transformants were evaluated for protein production and localization. Our results show that K. phaffii achieves a higher yield of secreted GM-CSF, while O. parapolymorpha produces a more biologically active form. Notably, this is the first demonstration that a K. phaffii strain displaying GM-CSF on its surface presents functionally active cytokine, as evidenced by TF-1 cell proliferation assays. The approach outlined here offers a promising platform for low-cost GM-CSF production to support the development of accessible wound-healing therapies.
2. Materials and Methods
2.1. Molecular-Biology Techniques
Standard cloning techniques were used as described by [32]. Genomic DNA of yeasts was isolated using the NucleoSpin® Tissue Kit (Macherey-Nagel, Duren, Germany). Restriction endonucleases and DNA ligase (Thermo Fisher Scientific Baltics, Vilnius, Lithuania) were used. Plasmid isolation from E. coli involved the Zyppy™ Plasmid Miniprep (Irvine, CA, USA). PCR-amplification of the fragments of interest was achieved with Phusion High-Fidelity DNA Polymerase (Thermo Fisher Scientific Baltics, Vilnius, Lithuania). PCRs were obtained using a GeneAmp PCR System 9700 thermocycler (Applied Biosystems, Foster City, CA, USA).
2.2. Plasmids and Strains
The human GM-CSF gene sequence (GenBank accession number M11220.1; https://www.ncbi.nlm.nih.gov/nuccore/M11220 (accessed on 14 September 2025)) was used as the reference sequence. Codon optimization for K. phaffii was performed using the ATGme online tool (http://atgme.org/ (accessed on 14 September 2025)) to match the codon usage preferences of the yeast genome (Figure S1).
For GM-CSF expression in K. phaffii, the base plasmid pPICZαB (Thermo Fisher Scientific) was used. This vector contains the alcohol oxidase (AOX) promoter from K. phaffii, which is methanol-inducible, and an S. cerevisiae α-factor secretion signal to direct GM-CSF to the culture medium. A 6 × His tag was added to the N-terminus of the GM-CSF coding sequence to enable purification by metal affinity chromatography. The resulting construct, designated pKp-CSF, was synthesized by GenScript (www.genscript.com (accessed on 14 September 2025)) (Figure 1a).

Figure 1.
Linear scheme of plasmid (a) pKp-CSF, (b) pOp-CSF, (c) pKp-CSF-GS, (d) pOp-CSF-GS, (e) pKp-CSF-SAG1, (f) pOp-CSF-SAG1, (g) pKp-CSF-GS-SAG1, and (h) pOp-CSF-GS-SAG1. Promoter and terminator of the AOX K. phaffii are indicated as blue boxes; promoter of the AOX O. polymorpha—as pink box; human GM-CSF—as gray box; glycine- and serine-rich linkers—as dark gray boxes; S. cerevisiae α-factor secretion signal and 6 × His tag—as green boxes; 3′ SAG1 S. cerevisiae—red boxes; selective markers ZeoR and NatR—as dark blue and orange boxes, respectively.
To construct a vector for expression in O. parapolymorpha, the K. phaffii AOX promoter in pKp-CSF was replaced with the O. polymorpha AOX promoter. The O. polymorpha AOX promoter was amplified from genomic DNA of O. polymorpha NCYC 495 using primers Ko1112 and Ko1113 (sequences of all used primers are shown in Table S1). The vector backbone lacking the K. phaffii AOX promoter was amplified from pKp-CSF using primers Ko1109 and Ko1110. Both fragments were digested with BglII and AscI and ligated at the corresponding sites. In the next step, the zeocin resistance marker in the resulting plasmid was replaced with a nourseothricin (NTC) resistance marker. The NTC marker was amplified from pRS41N [33] using primers OK42 and OK43. The plasmid backbone lacking the selection marker was amplified using primers SMF and SMR. PCR products were digested with NdeI and ligated. The resulting construct, named pOp-CSF, carried the GM-CSF gene under control of the O. polymorpha AOX promoter and an NTC resistance marker (Figure 1b).
To improve secretion efficiency, the GM-CSF gene was flanked by flexible, glycine- and serine-rich linker sequences. The linker-flanked GM-CSF variant was amplified using primers Ko1428 and Ko1432 and cloned into EcoRI/XbaI sites of pKp-CSF and pOp-CSF, replacing the native GM-CSF gene. The resulting constructs were designated pKp-CSF-GS and pOp-CSF-GS (Figure 1c,d).
For cell-surface display, the GM-CSF coding sequence was fused to the GPI-anchor domain derived from the C-terminus of the S. cerevisiae SAG1 gene encoding α-agglutinin. Two GM-CSF variants, native and GS-flanked, both lacking stop codons, were amplified from pKp-CSF using primer pairs Ko1424/Ko1433 and Ko1428/Ko1429, respectively. The SAG1 anchor fragment was amplified from genomic DNA of S. cerevisiae S288C using primers Ko1152 and Ko1153. Insert fragments were digested with EcoRI/XbaI (GM-CSF) and XbaI/SalI (SAG1), and ligated into EcoRI/SalI-digested vectors pKp-CSF, pOp-CSF, pKp-CSF-GS, and pOp-CSF-GS. The resulting plasmids were designated pKp-CSF-SAG1, pOp-CSF-SAG1, pKp-CSF-GS-SAG1, and pOp-CSF-GS-SAG1 (Figure 1e–h). All constructed plasmids were verified by sequencing. The nucleotide sequences of the target expression cassettes from plasmids pKp-CSF, pOp-CSF, pKp-CSF-GS, pOp-CSF-GS, pKp-CSF-SAG1, pOp-CSF-SAG1, pKp-CSF-GS-SAG1, and pOp-CSF-GS-SAG1 are shown in Figure S2. E. coli DH5α [Φ80dlacZΔM15, recA1, endA1, gyrA96, thi-1, hsdR17(r− K m+ K), supE44, relA1, deoR, Δ(lacZYA-argF) U169] was used for routine plasmid DNA cloning and propagation. DH5α was grown at 37 °C in LB medium as described [32]. Transformed E. coli cells were maintained in rich medium containing 50 mg/L of zeocin or 50 mg/L of NTC.
Plasmids for K. phaffii transformation (pKp-CSF, pKp-CSF-GS, pKp-CSF-SAG1, and pKp-CSF-GS-SAG1) were linearized using SacI and transformed into the K. phaffii GlycoSwitch SuperMan5 strain (https://pichia.com/try-pichia/try-pichia-glycoswitch/ (accessed on 14 September 2025)). Transformation of K. phaffii by electroporation was performed following the standard protocol described in the Pichia manuals from Invitrogen. Transformants were selected on YPD agar plates supplemented with 500 µg/mL zeocin. Plasmids for O. parapolymorpha transformation (pOp-CSF, pOp-CSF-GS, pOp-CSF-SAG1, and pOp-CSF-GS-SAG1) were linearized with BglII and transformed into the O. parapolymorpha (former Hansenula polymorpha) DL1-g24Sm strain [31]. Transformation of the yeast O. parapolymorpha by electroporation was carried out as described previously [34]. Transformants were selected on YPD agar plates containing 300 µg/mL NTC.
Transformant stabilization was achieved by alternating 15–20 generations of cultivation in non-selective and selective media. The resulting stable strains, including Kp-CSF, Kp-CSF-GS, Kp-CSF-SAG1, Kp-CSF-GS-SAG1, Op-CSF, Op-CSF-GS, Op-CSF-SAG1, and Op-CSF-GS-SAG1, were verified by colony PCR using primers Ko1424 and Ko1427 to confirm the presence of the GM-CSF gene.
2.3. Cultivation Condition of Engineered K. phaffii and O. parapolymorpha
To induce GM-CSF secretion, yeast strains K. phaffii GlycoSwitch SuperMan and its derivatives Kp-CSF and Kp-CSF-GS were pre-cultured from fresh agar plates in YPG medium (0.5% yeast extract, 1% peptone, 1% glycerol) for 24 h. Cultures were then inoculated at a starting OD600 of 5.0 into 100 mL of YPM medium (0.5% yeast extract, 1% peptone, 1% methanol) or mineral YNB medium (0.67% Yeast Nitrogen Base and 1% methanol) in 300 mL Erlenmeyer flasks and incubated at 30 °C for 120 h with shaking at 200 rpm. Methanol was supplemented every 24 h to a final concentration of 1%. O. parapolymorpha strain DL1-g24Sm and its derivatives Op-CSF and Op-CSF-GS were pre-cultured under similar conditions in YPD medium (0.5% yeast extract, 1% peptone, 2% glucose) and inoculated into YPM or YNB at a starting OD600 of 15. Cultures were grown in 100 mL YPM or YNB in 300 mL Erlenmeyer flasks at 37 °C with shaking for 120 h. Cells were harvested by centrifugation, washed twice with PBS, and pelleted. The culture medium was supplemented with 1 mM PMSF. Both cell pellets and culture supernatants were stored at −80 °C for further analysis. To induce cell surface immobilization of GM-CSF, strains Kp-CSF-SAG1, Kp-CSF-GS-SAG1 (K. phaffii) and Op-CSF-SAG1, Op-CSF-GS-SAG1 (O. parapolymorpha) were cultivated in YPM or YNB with 1% methanol for 72 h at 30 °C and 37 °C with starting OD600 of 0.3. Cells were washed three times with sterile PBS buffer. Aliquots of 100 μL of cells at OD600 of 100 were heat-inactivated by incubation at 60 °C for 10 min (K. phaffii) or twice for 15 min (O. parapolymorpha). Complete inactivation was confirmed by plating 30 μL of treated cells onto YPD agar to ensure the absence of growth. Inactivated cells were stored at −80 °C for further analysis. Cell biomass of yeast was determined with a Helios Gamma spectrophotometer (OD, 600 nm; cuvette, 10 mm) turbidimetrically with gravimetric calibration. A conversion factor of 1 OD600 = 5 × 107 cells/mL was used to calculate yeast cell numbers as described in Pichia pastoris expression kit manual from Invitrogen.
2.4. Biochemical Methods
Total cell extracts were prepared for immunoblotting as previously described [35]. For detection of secreted GM-CSF in the culture medium, proteins from 1.5 mL of culture supernatant were precipitated by the addition of 200 μL of 100% trichloroacetic acid, followed by incubation at −80 °C for 30 min. Precipitated proteins were pelleted by centrifugation (10 min at 16,000× g, room temperature), washed twice with ice-cold 80% acetone, air-dried, and resuspended in 50 μL of 1× SDS sample buffer. For analysis of concentrated GM-CSF in the medium, 50 μL of culture medium was mixed with 50 μL of 2× SDS sample buffer. All samples were boiled for 5 min. SDS-polyacrylamide gel electrophoresis was performed as described [36], and Western blotting was carried out according to established methods [37]. Proteins were detected using an enhanced chemiluminescence kit (LumiGO ECL Substrate Kit, Cleaver Scientific, Rugby, UK) following incubation with a polyclonal anti-GM-CSF antibody (Invitrogen). For deglycosylation, 10 µg of GM-CSF purified from the culture medium was treated with 2 µL of PNGase F (New England Biolabs, Ipswich, MA, USA) and incubated at 37 °C for 4 h, following the manufacturer’s instructions.
2.5. Purification
Proteins secreted into the culture medium and harvested after 120 h of cultivation were concentrated 50-fold using a Vivaspin 20 centrifugal concentrator (10,000 MWCO, PES membrane). Protein purification was performed by metal affinity chromatography. The concentrated medium was mixed with NEBExpress Ni Resin (1 mL of medium with 300 μL of 50% Ni Resin suspension), and binding was carried out at 4 °C for 18 h with gentle agitation. Purification was performed using NEBExpress Ni Spin Columns according to the manufacturer’s instructions. Bound proteins were eluted using 300 mM imidazole. The eluate was dialyzed for 18 h against 50 mM potassium phosphate buffer (pH 7.5) using a dialysis membrane with gentle stirring at 4 °C. The purified protein was sterilized by filtration through a 0.2 μm Minisart RC 4 membrane filter and stored at −80 °C for subsequent Western blot analysis and bioactivity assay. The concentrations of GM-CSF were measured spectrophotometrically by UV absorption at a wavelength of 280 nm, considering their molar extinction coefficients.
2.6. Immunofluorescent Assay
For immunofluorescence labeling, 200 μL of cells at OD600 of 1.0 were incubated with primary antibodies against GM-CSF (GM-CSF Polyclonal Antibody, Invitrogen, Carlsbad, CA, USA) at a 1:400 dilution for 3 h. The cell suspension was washed three times with PBS and then incubated with fluorescently labeled secondary antibodies (Anti-Rabbit IgG (H+L), CF488A, Sigma-Aldrich, St. Louis, MO, USA) at a 1:400 dilution for 2 h. Following incubation, cells were washed three more times with PBS. Fluorescence imaging was performed using a fluorescence microscope (Axio Imager A1, Carl Zeiss MicroImaging, Jena, Germany) equipped with a monochrome digital camera (AxioCam MRm, Carl Zeiss MicroImaging, Jena, Germany). Images were processed using AxioVision 4.5 software (Carl Zeiss MicroImaging) and ImageJ 1.53. For quantitative assessment of GM-CSF immobilization on the cell surface, fluorescence intensity was measured using a spectrofluorometer (RF-6000, Shimadzu, Kyoto, Japan) at excitation and emission wavelengths of 490 nm and 515 nm, respectively. Fluorescence was measured in an aqueous suspension of labeled cells adjusted to OD600 of 0.2.
2.7. Bioactivity of GM-CSF
Purified rhGM-CSF from each yeast species was added to 7.5 × 104 cells/mL of TF-1 (a GM-CSF dependent cell line) in RPMI plus 10% FBS and 2 mM L-glutamine to obtain a final GM-CSF concentration 20 ng/mL. Final concentration of yeast cell was 1.5 × 107 cells/mL. 200 µL of each combination was aliquoted into a 96-well plate and cultured for 72 h in a 5% CO2 incubator. The proliferation of TF-1 cells was measured by using a CCK-8 kit (Sigma-Aldrich, St. Louis, MO, USA) at OD 450 nm according to the manufacturer’s instructions. Negative controls were performed with TF-1 cells cultured without GM-CSF. Positive control was performed with TF-1 cells cultured with 3.5 ng/mL of rhGM-CSF (Sigma-Aldrich, St. Louis, MO, USA). All measurements were performed in triplicate and average values were reported.
2.8. Statistical Analysis
The results were analyzed with GraphPad Prism 8 and 9 and are presented as a mean (M) ± standard deviation (SD) of three independent experiments (n = 3) carried out in triplicate. A statistical evaluation was performed using two-way ANOVA analysis. A p-value of <0.05 was considered statistically significant.
3. Results and Discussion
3.1. Design of GM-CSF Expression Vectors and Generation of Recombinant Yeast Strains
To enable the expression of human GM-CSF in methylotrophic yeasts, we designed codon-optimized nucleotide sequence and constructed vectors for both secreted and surface-displayed forms of the protein. Expression in K. phaffii was driven by the methanol-inducible AOX promoter, and vectors included an S. cerevisiae α-factor secretion signal at the N-terminus, along with a 6 × His tag for purification (Figure 1a). For O. parapolymorpha, the AOX promoter from closely related yeast species O. polymorpha and a nourseothricin resistance marker were used (Figure 1b). To enhance secretion efficiency, flexible glycine–serine linkers were incorporated at both ends of the GM-CSF coding sequence, yielding GS-tagged constructs for both yeast species (Figure 1c,d). For surface immobilization, the GM-CSF variants were fused in-frame with the C-terminal domain of the S. cerevisiae cell wall protein α-agglutinin (Sag1), enabling GPI-anchor-mediated localization (Figure 1e–h). All plasmids were sequence-verified and transformed into glycoengineered yeast strains with humanized N-glycosylation profiles: K. phaffii SuperMan5 and O. parapolymorpha DL1-g24Sm. Stable integrants were selected and verified via colony PCR. Between 10 and 20 colonies of each type of transformant were analyzed to identify those with the highest levels of target protein secretion or surface immobilization. The selected strains were designated as Kp-CSF, Kp-CSF-GS, Op-CSF, and Op-CSF-GS for protein secretion, and as Kp-CSF-SAG1, Kp-CSF-GS-SAG1, Op-CSF-SAG1, and Op-CSF-GS-SAG1 for surface display of GM-CSF. These strains were used for subsequent studies.
3.2. GM-CSF Secretion by Engineered Yeast Strains
The Kp-CSF and Op-CSF transformants were cultivated under methanol induction in YPM or YNB medium, and GM-CSF expression was analyzed in both culture supernatants and cell extracts. In initial experiments, the recombinant GM-CSF remained intracellular, with no secretion observed in both yeast species (Figure 2a,b). Incorporation of glycine-serine linkers led to a marked improvement in secretion, likely due to reduced steric hindrance and improved folding of the recombinant protein. The GS-tagged variants of the secreted protein in strains Kp-CSF-GS and Op-CSF-GS yielded 15 mg/L and 3 mg/L, respectively. Comparatively, K. phaffii strain Kp-CSF-GS secreted noticeably more GM-CSF than O. parapolymorpha strain Op-CSF-GS under identical induction conditions. This finding is consistent with the well-documented higher secretion capacity of K. phaffii, which benefits from a robust secretory pathway and efficient ER-to-Golgi trafficking mechanisms [38]. Despite the secretion of the target protein into the culture medium, a significant portion of GM-CSF remained inside the cells. Further work is needed to improve GM-CSF secretion; however, the amount of secreted protein was sufficient for us to perform bioactivity assays and compare it with yeast cells displaying the cytokine on their surface.

Figure 2.
Western-blot analysis of cell extracts Kp-CSF (1), Kp-CSF-GS (3), and culture supernatants Kp-CSF (2), Kp-CSF-GS (4) of K. phaffii (a); cell extracts Op-CSF (1), Op-CSF-GS (3), and culture supernatants Op-CSF (2), Op-CSF-GS (4) of O. parapolymorpha (b); purified recombinant GM-CSF secreted into the culture: rhGM-CSF (Sigma) (1), from Kp-CSF-GS (2), and from Op-CSF-GS (3) (c); GM-CSF samples purified from the culture media of K. phaffii Kp-CSF-GS (1) and O. parapolymorpha Op-CSF-GS (2) before PNGase F treatment, and after treatment (3 and 4, respectively) (d). Minor differences in banding patterns between Figure 2c (lanes 2 and 3) and Figure 2d (lanes 1 and 2) can be attributed to variation in the amount of protein loaded and differences in SDS-PAGE running time. The molecular weight marker is shown on the left in kDa. Anti-GM-CSF polyclonal antibody (Invitrogen) was used for Western-blot. Glycosylated forms of GM-CSF are indicated by blue arrows, whereas the nonglycosylated form is indicated by a black arrow.
3.3. Purification and Analysis of Secreted GM-CSF
For protein purification, we chose to cultivate the yeast strains in YPM medium. Concentration of the mineral-based YNB medium resulted in the precipitation of mineral salts, which interfered with further concentration of the culture supernatant. Recombinant GM-CSF secreted into the culture YPM medium was purified using nickel-affinity chromatography under native conditions via its N-terminal His-tag. Western-blot analysis of GM-CSF from K. phaffii revealed a dominant band at ~19 kDa, corresponding to the expected molecular weight of GS-tagged variants of GM-CSF (Figure 2c). Additionally, we observed a smear resulting from heterogeneous glycosylation. The spots pattern of GM-CSF on the PVDF membrane shows that GM-CSF was glycosylated in a wide extent of 19 to 50 kDa. A similar pattern of glycosylated rhGM-CSF (28 kDa to >43 kDa) was previously reported in heterologous expression using P. pastoris by Tran et al. [23]. Recombinant GM-CSF from O. parapolymorpha exhibited smeared forms ranging from ~30 to ~50 kDa (Figure 2c). There are two N-linked and four O-linked glycosylation sites in hGM-CSF which are known as the reason of heterogeneous glycosylation in the expression of this protein by yeasts [23]. After treatment of purified GM-CSF from the culture media of K. phaffii and O. parapolymorpha with PNGase F, the high-molecular-weight smear shifted to a distinct band at approximately 19 kDa, as shown in Figure 2d. The heterogeneous glycosylation in our glycoengineered K. phaffii SuperMan5 and O. parapolymorpha DL1-g24Sm derivative strains, designed for humanized N-glycosylation profiles, likely enhances GM-CSF stability and solubility, as reported for other humanized yeast-expressed proteins [29]. However, residual heterogeneity in glycosylation may still impact receptor-binding affinity, potentially modulating GM-CSF bioactivity in these strains [22]. These results highlight the potential of glycoengineered yeasts for producing therapeutically relevant GM-CSF while indicating areas for further optimization of glycosylation profiles. Comparative analysis of wild-type and humanized yeast strains, together with deglycosylation assays, will be important to clarify the glycosylation status of recombinant GM-CSF.
3.4. Surface Display of GM-CSF
Fluorescence analysis was conducted for the K. phaffii and O. parapolymorpha strains Kp-CSF-SAG1 and Op-CSF-SAG1 expressing GM-CSF-SAG1. The fluorescence intensity of these strains was indistinguishable from that of the corresponding parental strains (Figure 3a). In contrast, the strains Kp-CSF-GS-SAG1 and Op-CSF-GS-SAG1 exhibited 2.2- and 1.3-fold increases in fluorescence, respectively, compared to their parental counterparts (Figure 3a). In immunofluorescence analysis, the parental strains displayed fluorescence levels comparable to autofluorescence (cells without antibody treatment or with secondary antibody only), confirming the specificity of antibody binding to the target epitope. Strains Kp-CSF-GS-SAG1 and Op-CSF-GS-SAG1 showed 1585 and 204 fluorescence units (FU), respectively, after subtraction of autofluorescence (Figure 3a). Due to the relatively modest increase observed for Op-CSF-GS-SAG1, further analysis was focused on Kp-CSF-GS-SAG1. Surface localization of GM-CSF was confirmed by indirect immunofluorescence microscopy, with a detectable fluorescence signal in approximately 40–50% of the cells, indicating successful display via the GPI-anchor (Figure 3b–e). The K. phaffii strain expressing GM-CSF-GS-SAG1 in YNB medium exhibited the highest fluorescence signal, reaching 1585 FU (Figure 3a). When cultivated in YPM medium, the same strain showed 947 FU, suggesting that defined media such as YNB may enhance surface anchoring or improve accessibility of the displayed cytokine (Figure 3a). This surface display approach offers a purification-free alternative for GM-CSF delivery and may serve as a platform for direct interaction with target immune cells.
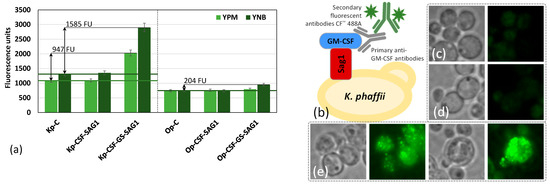
Figure 3.
Fluorescence of K. phaffii Kp-CSF-SAG1, Kp-CSF-GS-SAG1, and O. parapolymorpha Op-CSF-SAG1 and Op-CSF-GS-SAG1 strains, together with corresponding controls Kp-C (K. phaffii SuperMan5) and Op-C (O. parapolymorpha DL1-g24Sm), grown on methanol-supplemented rich (YPM) and synthetic (YNB) media and analyzed using an RF-6000 spectrofluorometer (Shimadzu) at excitation/emission wavelengths of 490/515 nm (a). Autofluorescence values (cells without antibody treatment or treated with secondary antibody only) are indicated as horizontal lines for strains of both species. Schematic representation of GM-CSF surface display in yeast (b), and fluorescence microscopy images of K. phaffii strains SuperMan5 (c), Kp-CSF-GS (d), and Kp-CSF-GS-SAG1 (e) grown on methanol-supplemented YNB medium. All samples were labeled with primary anti-GM-CSF antibodies followed by CF™ 488A-conjugated secondary antibodies.
3.5. Biological Activity of Secreted and Surface-Displayed GM-CSF
To assess the biological activity of recombinant GM-CSF, we performed proliferation assays using the human erythroleukemic TF-1 cell line, which is dependent on GM-CSF for survival and growth. Purified GM-CSF from O. parapolymorpha induced a 172% increase in TF-1 proliferation at a concentration of 20 ng/mL, whereas GM-CSF from K. phaffii induced a more modest 113% increase (Figure 4). The difference in biological activity likely stems from a combination of factors, including host-specific glycosylation patterns, protein folding efficiency, proteolytic processing, and polymerization tendencies. O. parapolymorpha may produce GM-CSF with glycosylation closer to human-like structures, better preserving receptor-binding sites, while K. phaffii’s higher secretion may lead to misfolding or aggregation, reducing bioactivity.
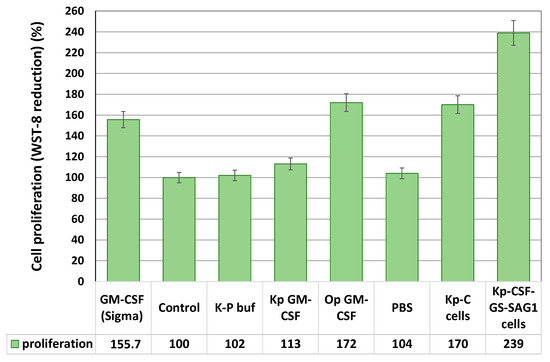
Figure 4.
Cell proliferation of TF-1 cells under the effect of the studied samples treated for 72 h. The negative control consisted of TF-1 cells cultured without supplementation and was set as 100%. The positive control included cells cultured with 3.5 ng/mL GM-CSF (Sigma). The proliferation of TF-1 cells was measured using a CCK-8 kit, in which the water-soluble tetrazolium salt (WST-8) is reduced by cellular dehydrogenases to produce a soluble colored formazan. The CCK-8 assay data are expressed as a percentage of the control (non-treated) group and presented as the mean ± SD (n = 4–6).
Interestingly, heat-inactivated K. phaffii cells displaying GM-CSF on their surface stimulated TF-1 cell proliferation by 239% when applied at a ratio of 200 yeast cells per one TF-1 cell (Figure 4). This response exceeded that observed with purified GM-CSF, likely due to the high local density and multivalent presentation of cytokine on the cell surface enhancing receptor clustering and downstream signaling. Notably, control yeast cells lacking GM-CSF also induced TF-1 proliferation (170%). Thus, K. phaffii cells displaying GM-CSF increased TF-1 cell proliferation by 1.41-fold compared to control yeast lacking GM-CSF. The induction of TF-1 cell proliferation by control yeast cells may result from nonspecific interactions between yeast cell wall components, particularly mannoproteins and β-glucans, and TF-1 cells. Yeast mannoproteins can mimic pathogen-associated patterns and engage pattern recognition receptors, potentially activating signaling cascades that support cell survival or proliferation [39]. A minor turbidity of the medium may have originated from heat-inactivated yeast cells; however, comparison with the control strain demonstrated that the effect was specifically due to surface-displayed GM-CSF.
3.6. Implications and Future Directions
These results demonstrate that glycoengineered K. phaffii and O. parapolymorpha strains can be used to produce both secreted and surface-anchored forms of human GM-CSF with measurable biological activity. Among the tested configurations, only constructs flanked by glycine-serine (GS) linkers enabled both efficient secretion and successful surface display of GM-CSF. In contrast, non-flanked constructs failed to support detectable secretion or anchoring of the cytokine on the yeast cell surface. The flexible GS linkers likely reduce steric hindrance and promote proper folding and trafficking of the recombinant protein, facilitating its translocation through the secretory pathway and presentation on the cell membrane [40].
To further improve performance, we propose several future directions. Since the majority of the target protein remained intracellular, cultivating the yeast in the presence of non-ionic detergents such as Tween 20 or Tween 80 may enhance GM-CSF secretion [41]. Increasing the gene copy number of GM-CSF in yeast strains may raise expression levels, particularly for surface-anchored forms. Replacing the inducible AOX promoter with the constitutive GAP promoter may simplify cultivation and yield more stable expression. Additionally, co-expression of protein folding chaperones such as Pdi1 and Kar2 could reduce aggregation within the endoplasmic reticulum and enhance secretion [42]. Beyond bioproduction, it would be of interest to test the immunomodulatory and regenerative effects of surface-displayed GM-CSF in animal models of wound healing, such as mouse skin injury systems. Such studies would help to validate the therapeutic potential of the yeast-based GM-CSF delivery system in vivo.
4. Conclusions
Yeast GM-CSF producers were constructed based on K. phaffii and O. parapolymorpha strains engineered for humanized protein glycosylation patterns. A K. phaffii strain was developed in which GM-CSF was displayed on the cell surface. The biological activity of this strain was confirmed, as its addition stimulated the proliferation of GM-CSF-dependent erythroblasts of the TF-1 line by 239%.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/applmicrobiol5030098/s1, Figure S1. Codon optimization of GM-CSF nucleotide sequence for enhanced expression in methylotrophic yeasts, Figure S2. The nucleotide sequences of the target expression cassettes from the plasmids pKp-CSF, pOp-CSF, pKp-CSF-GS, pOp-CSF-GS, pKp-CSF-SAG1, pOp-CSF-SAG1, pKp-CSF-GS-SAG1, and pOp-CSF-GS-SAG1. Promoter and terminator of the AOX K. phaffii are indicated as blue type; promoter of the AOX O. polymorpha—as orange type; human GM-CSF—as grey type; glycine- and serine-rich linkers—as violet italic type; S. cerevisiae α-factor secretion signal—as green type; 6 × His tag—as green bold type; 3’ SAG1 S. cerevisiae—red boxes; start ATG and stop TAA codons—bold, Table S1. List of primers used in this study (restriction sites are underlined).
Author Contributions
Conceptualization: A.S., K.D.; Methodology: O.D., N.F.; Investigation: O.D., N.F., R.S., K.D.; Writing—original draft preparation: O.D., K.D.; Writing—review and editing: R.S., A.S., K.D.; Project administration: A.S., K.D.; Funding acquisition: A.S., K.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a grant from the National Research Foundation of Ukraine (Project No. 2023.04/0032).
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors express our sincere gratitude to Hyun Ah Kang from the Department of Life Science, Chung-Ang University, Seoul 156-756, Korea, for kindly providing the DL1-g24Sm strain for this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Behm, B.; Babilas, P.; Landthaler, M.; Schreml, S. Cytokines, chemokines and growth factors in wound healing. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 812–820. [Google Scholar] [CrossRef]
- Bonnici, L.; Suleiman, S.; Schembri-Wismayer, P.; Cassar, A. Targeting signalling pathways in chronic wound healing. Int. J. Mol. Sci. 2023, 25, 50. [Google Scholar] [CrossRef]
- Demidova-Rice, T.N.; Hamblin, M.R.; Herman, I.M. Acute and impaired wound healing: Pathophysiology and current methods for drug delivery, part 2: Role of growth factors in normal and pathological wound healing: Therapeutic potential and methods of delivery. Adv. Skin Wound Care 2012, 25, 349–370. [Google Scholar] [CrossRef] [PubMed]
- Finnerty, C.C.; Jeschke, M.G.; Herndon, D.N.; Gamelli, R.; Gibran, N.; Klein, M.; Silver, G.; Arnoldo, B.; Remick, D.; Tompkins, R.G.; et al. Temporal cytokine profiles in severely burned patients: A comparison of adults and children. Mol. Med. 2008, 14, 553–560. [Google Scholar] [CrossRef]
- Rubbia-Brandt, L.; Sappino, A.P.; Gabbiani, G. Locally applied GM-CSF induces the accumulation of alpha-smooth muscle actin containing myofibroblasts. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1991, 60, 73–82. [Google Scholar] [CrossRef]
- Smith, C.H.; Allen, M.H.; Groves, R.W.; Barker, J.N. Effect of granulocyte macrophage-colony stimulating factor on Langerhans cells in normal and healthy atopic subjects. Br. J. Dermatol. 1998, 139, 239–246. [Google Scholar] [CrossRef]
- Mann, A.; Breuhahn, K.; Schirmacher, P.; Blessing, M. Keratinocyte-derived granulocyte-macrophage colony stimulating factor accelerates wound healing: Stimulation of keratinocyte proliferation, granulation tissue formation, and vascularization. J. Investig. Dermatol. 2001, 117, 1382–1390. [Google Scholar] [CrossRef]
- Kaplan, G.; Walsh, G.; Guido, L.S.; Meyn, P.; Burkhardt, R.A.; Abalos, R.M.; Barker, J.; Frindt, P.A.; Fajardo, T.T.; Celona, R. Novel responses of human skin to intradermal recombinant granulocyte/macrophage-colony-stimulating factor: Langerhans cell recruitment, keratinocyte growth, and enhanced wound healing. J. Exp. Med. 1992, 175, 1717–1728. [Google Scholar] [CrossRef]
- Carral, A.; de la Rubia, J.; Martín, G.; Martínez, J.; Sanz, G.; Jarque, I.; Sempere, A.; Soler, M.A.; Marty, M.L.; Sanz, M.A. Factors influencing hematopoietic recovery after autologous blood stem cell transplantation in patients with acute myeloblastic leukemia and with non-myeloid malignancies. Bone Marrow Transplant. 2002, 29, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Gong, S.J.; Xu, Y.H.; Hambly, B.D.; Bao, S. Impaired cutaneous wound healing in granulocyte/macrophage colony-stimulating factor knockout mice. Br. J. Dermatol. 2007, 157, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, K.; Ishii, G.; Ochiai, A.; Esumi, H. Improvement of the breaking strength of wound by combined treatment with recombinant human G-CSF, recombinant human M-CSF, and a TGF-beta1 receptor kinase inhibitor in rat skin. Cancer Sci. 2008, 99, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Shen, J.; Yao, M.; Beagley, K.W.; Hambly, B.D.; Bao, S. Granulocyte-macrophage colony-stimulating factor enhances wound healing in diabetes via upregulation of proinflammatory cytokines. Br. J. Dermatol. 2010, 162, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Forno, G.; Bollati Fogolin, M.; Oggero, M.; Kratje, R.; Etcheverrigaray, M.; Conradt, H.S.; Nimtz, M. N- and O-linked carbohydrates and glycosylation site occupancy in recombinant human granulocyte-macrophage colony-stimulating factor secreted by a Chinese hamster ovary cell line. Eur. J. Biochem. 2004, 271, 907–919. [Google Scholar] [CrossRef]
- Srinivasa Babu, K.; Muthukumaran, T.; Antony, A.; Prem Singh Samuel, S.D.; Balamurali, M.; Murugan, V.; Meenakshisundaram, S. Single step intein-mediated purification of hGMCSF expressed in salt-inducible E. coli. Biotechnol. Lett. 2009, 31, 659–664. [Google Scholar] [CrossRef] [PubMed]
- James, E.A.; Wang, C.; Wang, Z.; Reeves, R.; Shin, J.H.; Magnuson, N.S.; Lee, J.M. Production and characterization of biologically active human GM-CSF secreted by genetically modified plant cells. Protein Expr. Purif. 2000, 19, 131–138. [Google Scholar] [CrossRef]
- Chiou, C.J.; Wu, M.C. Expression of human granulocyte-macrophage colony-stimulating factor gene in insect cells by a baculovirus vector. FEBS Lett. 1990, 259, 249–253. [Google Scholar] [CrossRef]
- Dorr, R.T. Clinical properties of yeast-derived versus Escherichia coli-derived granulocyte-macrophage colony-stimulating factor. Clin. Ther. 1993, 15, 19–28. [Google Scholar]
- Bergès, H.; Joseph-Liauzun, E.; Fayet, O. Combined effects of the signal sequence and the major chaperone proteins on the export of human cytokines in Escherichia coli. Appl. Environ. Microbiol. 1996, 62, 55–60. [Google Scholar] [CrossRef]
- Khasa, Y.P.; Khushoo, A.; Tapryal, S.; Mukherjee, K.J. Optimization of human granulocyte macrophage-colony stimulating factor (hGM-CSF) expression using asparaginase and xylanase gene’s signal sequences in Escherichia coli. Appl. Biochem. Biotechnol. 2011, 165, 523–537. [Google Scholar] [CrossRef]
- Sahdev, S.; Khattar, S.K.; Saini, K.S. Production of active eukaryotic proteins through bacterial expression systems: A review of the existing biotechnology strategies. Mol. Cell. Biochem. 2008, 307, 249–264. [Google Scholar] [CrossRef]
- DeLamarter, J.F.; Mermod, J.J.; Liang, C.M.; Eliason, J.F.; Thatcher, D.R. Recombinant murine GM-CSF from E. coli has biological activity and is neutralized by a specific antiserum. EMBO J. 1985, 4, 2575–2581. [Google Scholar] [CrossRef]
- Gasmi, N.; Lassoued, R.; Ayed, A.; Tréton, B.; Chevret, D.; Nicaud, J.M.; Kallel, H. Production and characterization of human granulocyte-macrophage colony-stimulating factor (hGM-CSF) expressed in the oleaginous yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2012, 96, 89–101. [Google Scholar] [CrossRef]
- Tran, A.M.; Nguyen, T.T.; Nguyen, C.T.; Huynh-Thi, X.M.; Nguyen, C.T.; Trinh, M.T.; Tran, L.T.; Cartwright, S.P.; Bill, R.M.; Tran-Van, H. Pichia pastoris versus Saccharomyces cerevisiae: A case study on the recombinant production of human granulocyte-macrophage colony-stimulating factor. BMC Res. Notes 2017, 10, 148. [Google Scholar] [CrossRef]
- Pykhtina, M.; Miroshnichenko, S.; Romanov, V.; Grazhdantseva, A.; Kochneva, G.; Beklemishev, A. Construction of recombinant human GM-CSF and GM-CSF-ApoA-I fusion protein and evaluation of their biological activity. Pharmaceuticals 2021, 14, 459. [Google Scholar] [CrossRef] [PubMed]
- Pal, Y.; Khushoo, A.; Mukherjee, K.J. Process optimization of constitutive human granulocyte-macrophage colony-stimulating factor (hGM-CSF) expression in Pichia pastoris fed-batch culture. Appl. Microbiol. Biotechnol. 2006, 69, 650–657. [Google Scholar] [CrossRef]
- Vogl, T.; Fischer, J.E.; Hyden, P.; Wasmayer, R.; Sturmberger, L.; Glieder, A. Orthologous promoters from related methylotrophic yeasts surpass expression of endogenous promoters of Pichia pastoris. AMB Express 2020, 10, 38. [Google Scholar] [CrossRef]
- Lv, W.; Cai, M. Advancing recombinant protein expression in Komagataella phaffii: Opportunities and challenges. FEMS Yeast Res. 2025, 25, foaf010. [Google Scholar] [CrossRef] [PubMed]
- Besleaga, M.; Ebner, K.; Glieder, A.; Spadiut, O.; Kopp, J. Chances and drawbacks of derepressed recombinant enzyme production in continuous cultivations with Komagataella phaffii. Front. Bioeng. Biotechnol. 2025, 13, 1523037. [Google Scholar] [CrossRef]
- Hamilton, S.R.; Gerngross, T.U. Glycosylation engineering in yeast: The advent of fully humanized yeast. Curr. Opin. Biotechnol. 2007, 18, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Boonekamp, F.J.; Knibbe, E.; Vieira-Lara, M.A.; Wijsman, M.; Luttik, M.A.H.; van Eunen, K.; Ridder, M.D.; Bron, R.; Almonacid Suarez, A.M.; van Rijn, P.; et al. Full humanization of the glycolytic pathway in Saccharomyces cerevisiae. Cell Rep. 2022, 39, 111010. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.A.; Kim, H.; Oh, D.B.; Kwon, O.; Kang, H.A. Remodeling of the glycosylation pathway in the methylotrophic yeast Hansenula polymorpha to produce human hybrid-type N-glycans. J. Microbiol. 2012, 50, 341–348. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory: New York, NY, USA, 2012. [Google Scholar]
- Taxis, C.; Knop, M. System of centromeric, episomal, and integrative vectors based on drug resistance markers for Saccharomyces cerevisiae. Biotechniques 2006, 40, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Faber, K.N.; Haima, P.; Harder, W.; Veenhuis, M.; Ab, G. Highly-efficient electrotransformation of the yeast Hansenula polymorpha. Curr. Genet. 1994, 25, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Baerends, R.J.; Faber, K.N.; Kram, A.M.; Kiel, J.A.; van der Klei, I.J.; Veenhuis, M. A stretch of positively charged amino acids at the N terminus of Hansenula polymorpha Pex3p is involved in incorporation of the protein into the peroxisomal membrane. J. Biol. Chem. 2000, 275, 9986–9995. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Kyhse-Andersen, J. Electroblotting of multiple gels: A simple apparatus without buffer tank for rapid transfer of proteins from polyacrylamide to nitrocellulose. J. Biochem. Biophys. Methods 1984, 10, 203–209. [Google Scholar] [CrossRef]
- Delic, M.; Valli, M.; Graf, A.B.; Pfeffer, M.; Mattanovich, D.; Gasser, B. The secretory pathway: Exploring yeast diversity. FEMS Microbiol. Rev. 2013, 37, 872–914. [Google Scholar] [CrossRef]
- Baek, K.R.; Lee, H.J.; Jeong, K.H.; Park, B.R.; Kim, S.J.; Seo, S.O. Parabiotic immunomodulatory activity of yeast cell wall polysaccharides from Saccharomyces cerevisiae and S. boulardii. J. Funct. Foods 2024, 123, 106577. [Google Scholar] [CrossRef]
- Cho, J.S.; Oh, H.J.; Jang, Y.E.; Kim, H.J.; Kim, A.; Song, J.A.; Lee, E.J.; Lee, J. Synthetic pro-peptide design to enhance the secretion of heterologous proteins by Saccharomyces cerevisiae. Microbiologyopen 2022, 11, e1300. [Google Scholar] [CrossRef]
- Bahrami, A.; Shojaosadati, S.A.; Khalilzadeh, R.; Mohammadian, J.; Farahani, E.V.; Masoumian, M.R. Prevention of human granulocyte colony-stimulating factor protein aggregation in recombinant Pichia pastoris fed-batch fermentation using additives. Biotechnol. Appl. Biochem. 2009, 52, 141–148. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, H.L.; Xue, C.; Xiong, X.H.; Yao, X.Q.; Li, X.Y.; Chen, H.P.; Liu, Z.M. Enhanced secretion of heterologous proteins in Pichia pastoris following overexpression of Saccharomyces cerevisiae chaperone proteins. Biotechnol. Prog. 2006, 22, 1090–1095. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).