1. Introduction
Ready-to-eat (RTE) refrigerated foods have emerged as notable vehicles for foodborne pathogens, especially when minimal processing and mild formulations allow microbial survival [
1]. Hummus, a popular chickpea-based dip, exemplifies this risk. In the United States,
Listeria monocytogenes contamination of hummus has been linked to multiple outbreaks and recalls in recent years [
2,
3]. Between 2002 and 2017, at least 17 hummus-associated outbreaks were reported nationally, resulting in approximately 1105 illnesses, 64 hospitalizations, and 4 deaths [
1]. Notably, two listeriosis outbreaks in 2013 accounted for all four fatalities, including one California incident that sickened 28 people [
1]. Even in cases without confirmed illnesses, contamination has prompted large-scale product recalls—for instance, Sabra recalled over 30,000 cases of hummus in 2015 after routine testing found
L. monocytogenes [
3]. These events underscore the food safety vulnerability of minimally processed dips, such as hummus, and the critical need for effective control measures.
Controlling pathogens in RTE dips is challenging because their intrinsic properties often permit bacterial growth under refrigeration [
1]. Hummus typically has water activity above 0.92 and pH above 4.4—conditions that support the growth of many foodborne pathogens. Unlike many cooked foods that undergo a final lethal heat treatment, hummus is typically consumed without such treatment after preparation, making it susceptible to post-processing contamination. While ingredients, such as chickpeas, are cooked during preparation, other components, such as tahini, may not be heat treated, and improper sanitation, poor water quality, or contaminated packaging materials can introduce pathogens.
L. monocytogenes, in particular, is a psychrotrophic pathogen capable of proliferating at refrigeration temperatures. Previous studies have shown that
L. monocytogenes not only survives but may increase in hummus during storage, particularly when mild acidification and salt levels are used [
2]. Under temperature-abuse conditions (e.g., 30 °C),
Listeria can grow rapidly, with levels exceeding 8 log CFU/g within 48 h in some formulations, highlighting the importance of additional safety barriers.
Modern consumer preferences further complicate formulation strategies. There is growing demand for “clean-label” foods—products with short ingredient lists free from synthetic additives or preservatives [
4]. In response, manufacturers are pursuing natural food safety interventions to meet regulatory and market expectations. These trends have increased interest in bio-preservation strategies that utilize naturally occurring microorganisms or their metabolites to inhibit pathogens [
5]. Among these, protective cultures and probiotic microorganisms have shown potential to maintain microbial stability in minimally processed foods without altering flavor or texture [
6,
7].
Probiotics are traditionally defined as “live microorganisms which, when administered in adequate amounts, confer a health benefit to the host” [
8,
9]. Beyond their health effects, certain strains also exert antimicrobial activity through nutrient competition, pH reduction, or secretion of bacteriocins. While lactic acid bacteria have been widely studied for this purpose, their viability and functional performance are often compromised in non-fermented, refrigerated, or high-pH foods [
5,
10]. In contrast, spore-forming
Bacillus species—including
Bacillus subtilis—are exceptionally resilient and able to withstand diverse processing and storage conditions [
11,
12,
13].
Bacillus subtilis produces endospores—highly resistant, dormant structures capable of surviving heat, desiccation, acid, and other harsh food environments [
11,
12]. Unlike vegetative probiotic cells, spores can remain stable for months, only germinating when triggered by favorable environmental conditions, such as elevated temperature or nutrient availability [
6,
13,
14]. This dormancy makes them ideal for use in RTE foods; they remain inactive under refrigerated storage, posing minimal risk to product quality, and become metabolically active only when the food is exposed to conditions that would also promote pathogen growth—such as temperature abuse [
4,
6,
15]
Once germinated,
B. subtilis can inhibit pathogens through several mechanisms: rapid nutrient uptake, production of siderophores (e.g., bacillibactin), competitive exclusion of colonization sites, and secretion of narrow-spectrum antimicrobials, such as subtilosin A and subtilin, which are particularly effective against Gram-positive bacteria, such as
Listeria monocytogenes [
15,
16,
17,
18]. Because Gram-negative organisms, such as
Salmonella and
E. coli, have protective outer membranes, they are less sensitive to these peptide-based antimicrobials, making Gram-positive pathogens more susceptible to inhibition by
Bacillus-based probiotics [
5].
This selective activity has been supported in multiple studies and was also observed in the preliminary broth experiments reported here, where
Bacillus subtilis spores significantly inhibited
L. monocytogenes under nutrient-limited conditions, yet had a limited impact on
E. coli O157:H7 and
Salmonella Typhimurium. However, whether this efficacy translates to complex food matrices, such as hummus, remains unclear. Hummus presents a high-viscosity, particulate-rich environment that may hinder probiotic–pathogen interactions and create micro-niches in which
Listeria can persist undisturbed [
2,
19].
To address this knowledge gap, the present study was designed to investigate the suitability of Bacillus subtilis spores as biocontrol agents in both simplified and complex food systems. Specifically, the objectives were (1) to assess the ability of a B. subtilis spore cocktail to inhibit L. monocytogenes, E. coli O157:H7, and S. Typhimurium under varying nutrient conditions in tryptic soy broth, and (2) to evaluate the probiotic’s effectiveness in controlling L. monocytogenes in a temperature-abused hummus matrix. The following sections detail the experimental design, microbial enumeration methods, and observed results under both laboratory and food system conditions.
2. Materials and Methods
2.1. Preparation of Microbial Cultures
2.1.1. Listeria monocytogenes
For this study, four strains of L. monocytogenes were used and included: Scott A-2, V7-2, PMM39-2, and PMM383-2. Scott A and V7 are well-known strains; PMM383 and PMM39 were isolated from raw and ready-to-eat (RTE) meat products, respectively) These strains were previously adapted to streptomycin (100 μg/mL), through spontaneous mutation. The L. monocytogenes cultures were grown individually in tryptic soy broth (TSB; Difco, Becton Dickinson, Sparks, MD, USA), supplemented with 100 μg/mL streptomycin and 100 μg/mL rifamycin. After approximately 18 h of incubation at 30 °C, they were centrifuged (3000× g for 15 min), the supernatant was then disposed of, and the pellets were resuspended in saline solution (9 mg/mL sodium chloride solution). A four-strain cocktail was made for L. monocytogenes by combining 2 mL of each strain. The final concentration of the bacterial cocktail was adjusted to approximately 5.1 ± 0.2 log CFU/mL.
2.1.2. Escherichia coli O157:H7
Five strains of
E. coli O157:H7 used in this study: 1 (Beef isolate), 5 (human isolate), 932 (human isolate), E009 (Beef isolate) and E0122 (cattle isolate). Each
E. coli strain used had previously been adapted to 50 mg/L nalidixic acid [
20] through spontaneous mutation. These strains were individually grown in TSB supplemented with 50 mg/L nalidixic acid for approximately 18 h at 37 °C. At the end of the incubation period, bacterial strains were washed by centrifugation as described above. A five-strain cocktail was made for
E. coli by mixing an equal amount of each strain in a 15 mL capacity sterile centrifuge tube. The final concentration of the cocktail was adjusted to 5.2 ± 0.3 log CFU/mL.
2.1.3. Salmonella Typhimurium DT104
Three strains of S. Typhimurium DT 104 were used and included: H2662 (cattle isolate), 13068A (cattle isolate), and H3279 (human isolate). S. Typhimurium strains were grown individually in TSB, which had been supplemented with 32 mg/L ampicillin, 16 mg/L tetracycline, and 64 mg/L streptomycin. After the strains grew for approximately 18 h at 37 °C, they were washed by centrifugation and a five-strain bacterial cocktail containing a pathogen population of 5.1 ± 0.2 log CFU/mL was prepared using the method described above.
All bacterial cocktails were centrifuged at 4000× g for 10 min, and then the pellets were resuspended in phosphate-buffered saline solution (PBS) to create experiment-ready cultures.
2.1.4. Probiotic Strains
Two Bacillus subtilis strains were used in this study: Bacillus subtilis 1 and Bacillus subtilis Novonesis HU58™. Both strains are commercially available spore-forming probiotics that have been designated as Generally Recognized as Safe (GRAS) by the United States Food and Drug Administration (FDA) as well as by an independent expert panel. The spore preparations were obtained from commercial suppliers in spray-dried powder form. The initial concentrations of the powdered products were 11.47 log CFU/g for B. subtilis 1 and 10.18 log CFU/g for HU58™. For all experimental assays, a mixed spore cocktail was prepared by combining 50 mg of B. subtilis 1 and 950 mg of HU58™ powder, thereby reflecting a formulation dominated by the HU58™ strain.
2.2. Impact of Bacillus spores on the Growth of Foodborne Pathogens
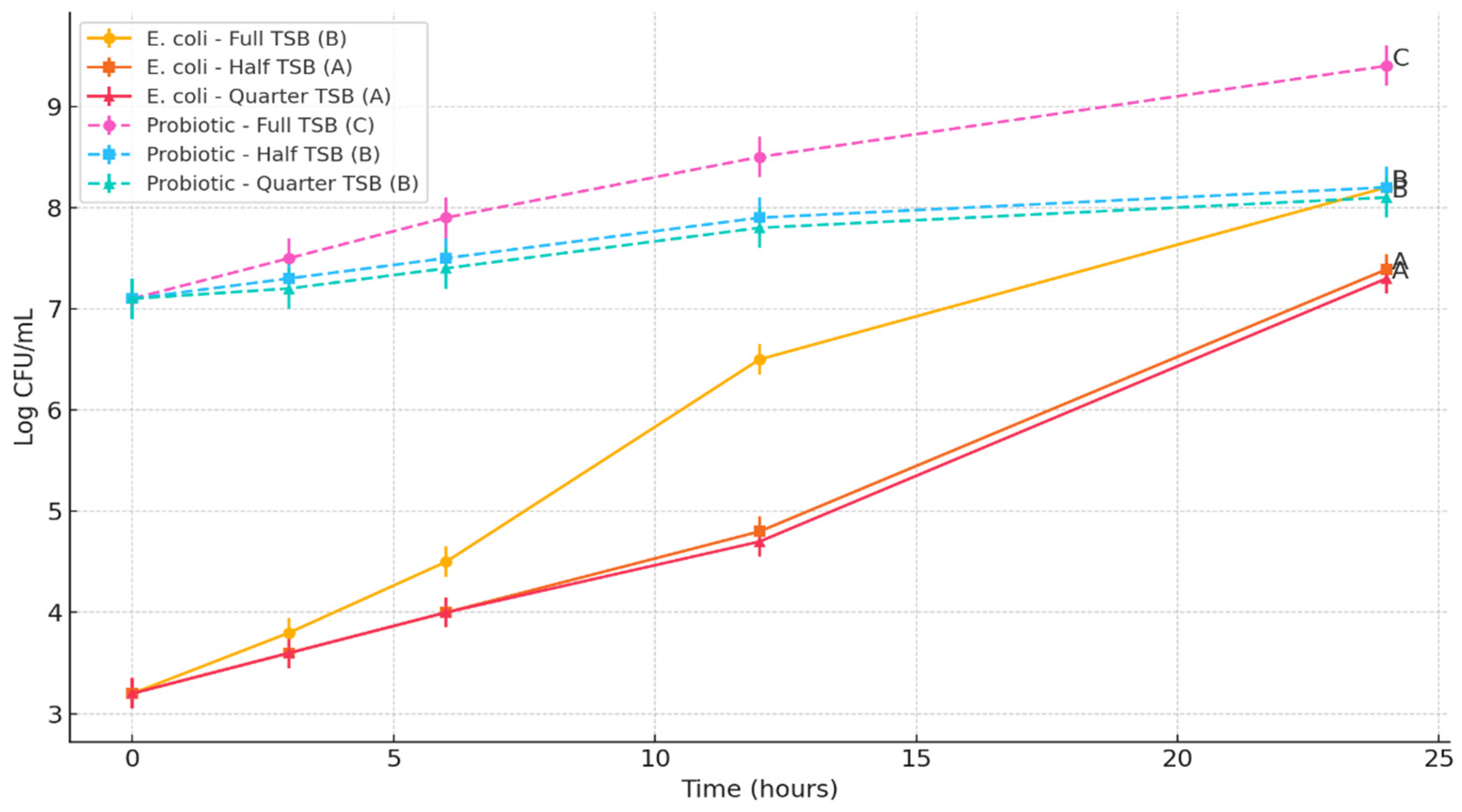
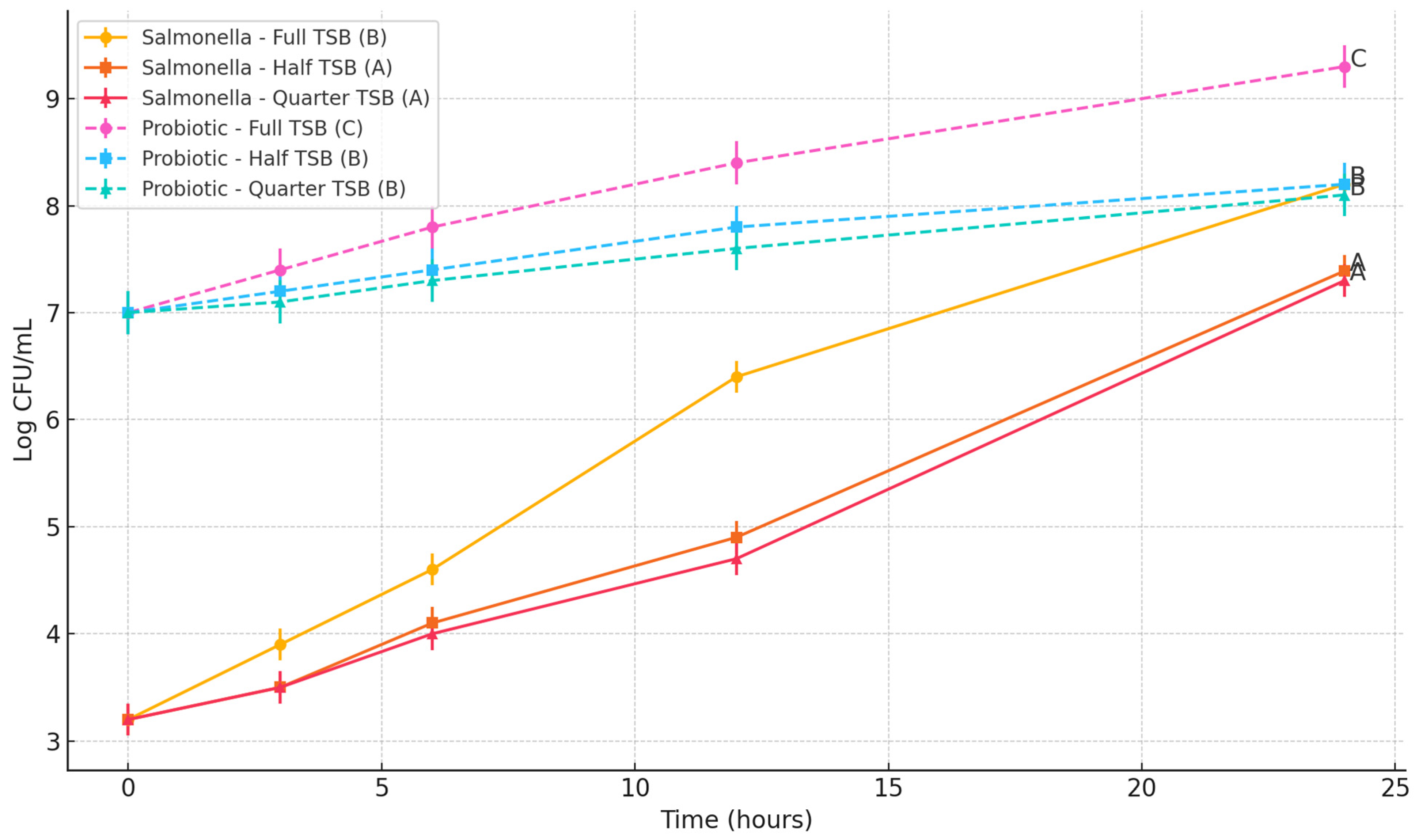
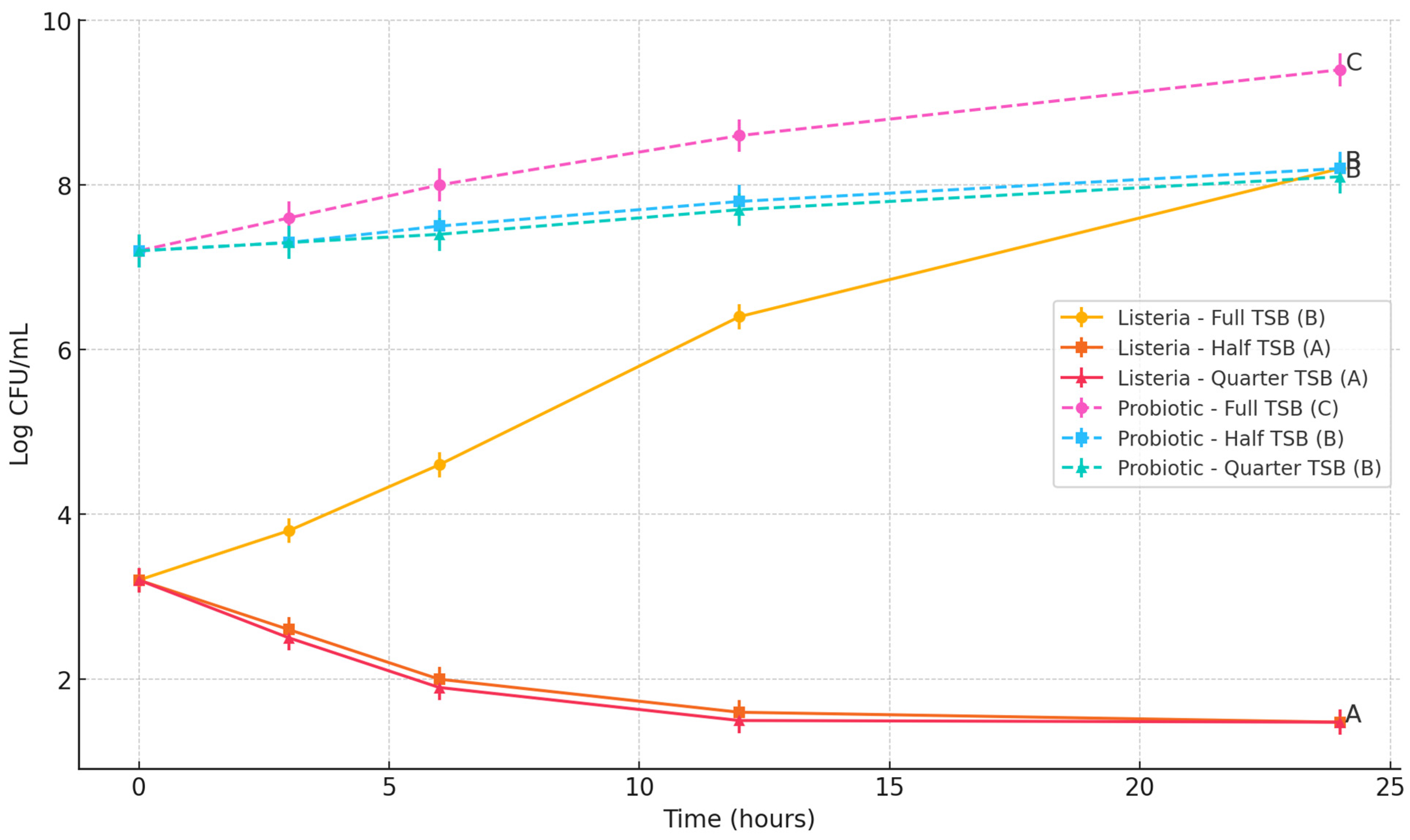
This study assessed the influence of the Bacillus subtilis spore cocktail on the growth of three foodborne pathogens: Escherichia coli O157:H7, Salmonella Typhimurium, and Listeria monocytogenes. The objective was to determine whether the probiotic spores could inhibit the proliferation of these pathogens in nutrient-rich and nutrient-limited environments.
To simulate varying nutrient availability, three formulations of tryptic soy broth (TSB) were prepared: full-strength (100%), half-strength (50%), and quarter-strength (25%). Each TSB formulation (10 mL per tube) was dispensed into sterile 15 mL polypropylene centrifuge tubes. All media were autoclaved at 121 °C for 15 min and cooled to room temperature prior to inoculation.
Each tube was individually inoculated with one of the target pathogens to achieve an initial concentration of approximately 3 log CFU/mL. The probiotic spore cocktail was then added to achieve a final concentration of approximately 7 log CFU/mL, in accordance with WHO recommendations suggesting a probiotic threshold between 6 to 9 log CFU/mL to confer health benefits [
8,
9]. The higher probiotic dose was selected to mimic typical probiotic supplementation levels used in commercial and therapeutic applications.
The tubes were incubated aerobically at 37 °C for E. coli O157:H7 and S. Typhimurium, and at 30 °C for L. monocytogenes, which better reflects its psychrotrophic growth characteristics. At 0, 3, 6, 12, and 24 h time points, 2 mL of each culture was sampled. One milliliter was used for the enumeration of pathogens and total viable counts, and the remaining 1 mL was used for spore germination analysis.
For microbial enumeration, 1 mL of the samples wereserially diluted in 0.1% buffered peptone water and surface-plated onto selective and non-selective media. E. coli O157:H7 was enumerated on MacConkey agar (BD Difco, Sparks, MD, USA) supplemented with 50 mg/L nalidixic acid. S. Typhimurium was plated on Xylose Lysine Deoxycholate (XLD) agar (BD Difco, Sparks, MD, USA) supplemented with 32 mg/L ampicillin, 16 mg/L tetracycline, and 64 mg/L streptomycin. L. monocytogenes was enumerated using Oxford agar base (Oxoid, Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 100 μg/mL streptomycin and 100 μg/mL rifamycin. For total bacterial counts, samples were plated on tryptic soy agar (TSA; BD Difco, Sparks, MD, USA). The survival of probiotics was analyzed by subtracting the colony counts received on selective media from total bacterial counts on TSA.
The second portion of the sample (1 mL) was used to assess spore germination over the incubation period. Samples were transferred to sterile 15 mL borosilicate glass tubes and subjected to a thermal shock by immersion in a pre-heated water bath (Thermo Scientific Precision GP, Waltham, MA, USA) at 80 °C for 10 min using a previously published method [
13]. This treatment selectively inactivated vegetative cells while preserving heat-resistant spores. The thermally treated samples were then plated on TSA to enumerate remaining spore populations.
2.3. Effectiveness of Bacillus spores to Control L. monocytogenes in Hummus
2.3.1. Hummus Preparation
Hummus was prepared using a standardized protocol to ensure batch-to-batch consistency and reproducibility for experimental use. Chickpeas (
Cicer arietinum) served as the primary ingredient. For each batch, 453 g of dry chickpeas (approximately one 16 oz packet) were weighed and soaked overnight in potable water supplemented with 0.5 g of baking soda per 450 g of chickpeas [
19]. Sodium bicarbonate has been shown to enhance textural properties by promoting seed coat breakdown and reducing cooking time (
Figure 1).
Following hydration, the chickpeas were drained and cooked in a domestic pressure cooker until fully softened—typically for six whistles, equivalent to approximately 25 min of cooking time. This step ensured complete gelatinization of starches and optimal softening. After cooking, chickpeas were drained and weighed, yielding a final cooked mass of approximately 800 g. Cooked chickpeas not used immediately were stored in sterile, airtight containers at −20 °C to maintain product integrity.
Tahini was prepared in-house by blending toasted white sesame seeds and extra virgin olive oil in a 70:30 (w/w) ratio. A total of 700 g of sesame seeds were lightly toasted to enhance flavor development and reduce moisture content. The seeds were then ground with olive oil using a high-speed blender until a smooth, uniform paste was obtained. Prepared tahini was aliquoted into 65 g portions and stored at 4 °C in sterile containers to ensure consistency across trials.
The final hummus formulation consisted of 70% (w/w) mashed chickpeas, 10% tahini, 7% water, 7% olive oil, 5% freshly squeezed lemon juice, and 1% salt. All ingredients were sourced from a commercial online retailer (Amazon.com, Seattle, WA, USA) to ensure consistency across batches. The components were combined and homogenized using a laboratory-grade blender (Waring Commercial Laboratory Blender, Model 7011HS, Waring Products, Stamford, CT, USA) until a smooth, creamy consistency was achieved. To ensure uniformity in sample texture across replicates, exactly 200.61 g of blended hummus was portioned for each experiment, and 15.1 mL of sterile deionized water was added to standardize moisture content and product viscosity.
All prepared hummus samples were stored at 4 °C in sterile, sealed containers and used within 24 h of preparation. This approach ensured microbial stability and maintained the physicochemical properties essential for downstream experimental analyses.
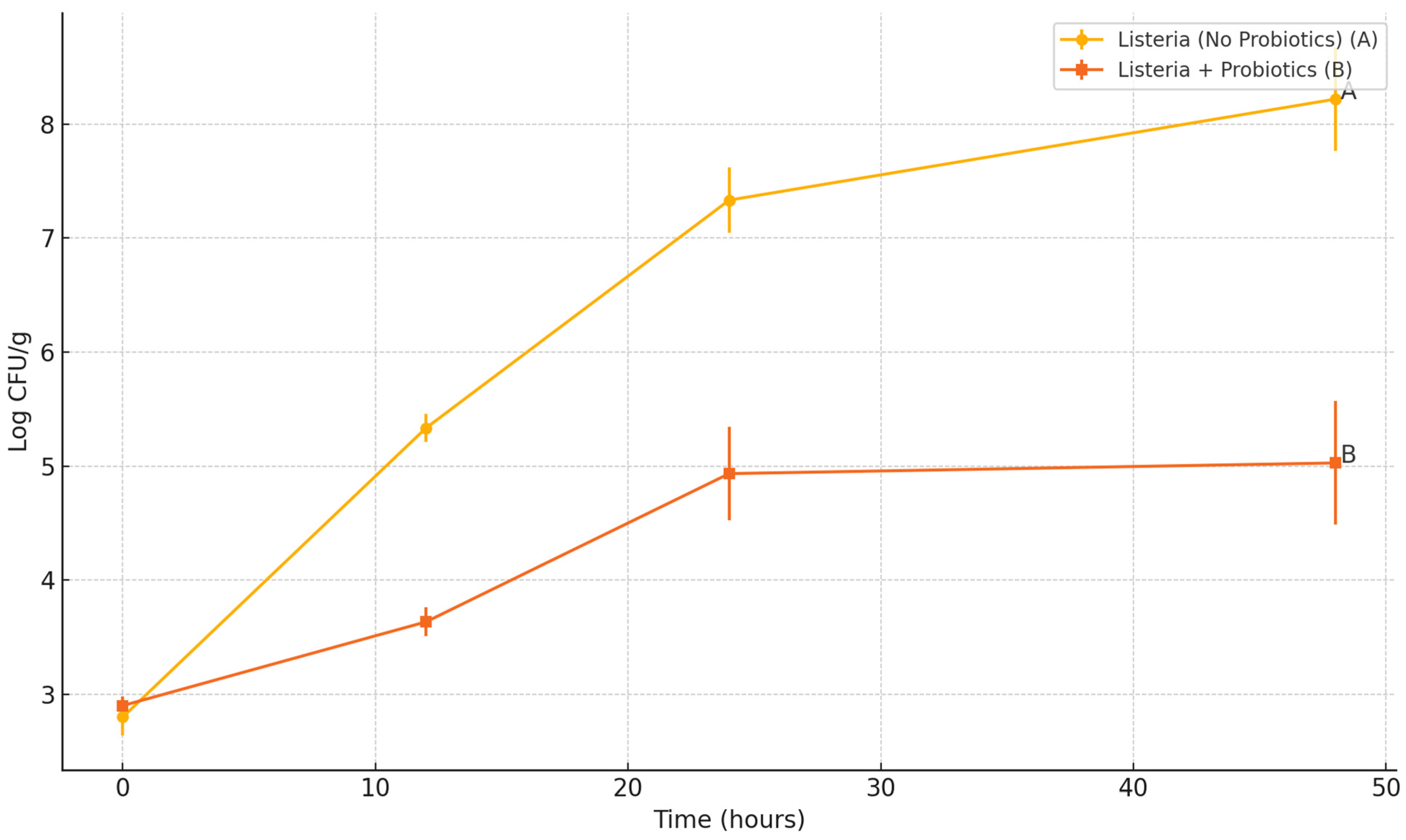
2.3.2. Inoculation of Hummus with L. monocytogenes and Bacillus spores
To assess the survival dynamics of L. monocytogenes and probiotic Bacillus subtilis spores in a refrigerated hummus matrix, the prepared hummus (as described above) was aseptically portioned into sterile polypropylene containers, each containing 25 g of product. The hummus samples were then inoculated with a cocktail of L. monocytogenes strains to achieve a final concentration of approximately 3 log CFU/g. Simultaneously, a mixed probiotic spore cocktail (comprising B. subtilis 1 and HU58™ strains) was added to reach a final concentration of 7 log CFU/g.
Following inoculation, all samples were mixed thoroughly within their respective containers to ensure homogeneous distribution of both target organisms. The containers were then stored at 30 ± 1 °C to simulate temperature-abuse conditions. Sampling was conducted at 0, 12, 24, and 48 h post-inoculation to monitor changes in microbial populations over time.
For microbial enumeration, one full 25 g sample container was removed at each sampling point. The entire hummus sample was transferred to a sterile stomacher bag and diluted in phosphate-buffered saline (PBS; pH 7.2) at a 1:3 ratio (w/v). The samples were homogenized using a stomacher (Seward Stomacher 400 Circulator, Model 0400/CLR, Seward Laboratory Systems Inc., Bohemia, NY, USA) for 1 min.
Following homogenization, aliquots were serially diluted in 0.1% buffered peptone water. L. monocytogenes was enumerated by surface plating on Oxford agar base with antibiotics as described above to inhibit background flora. Total viable bacterial counts (including probiotics and pathogens) were determined by plating on TSA.
To assess the persistence and germination of probiotic spores in the hummus matrix, a portion of each homogenized sample (1 mL) was subjected to thermal treatment. The sample was transferred to a sterile 15 mL glass tube and heated in a water bath at 80 °C for 15 min to eliminate vegetative cells. Surviving spores were enumerated by surface plating on TSA.
2.4. Statistical Analysis
All experiments were performed in triplicate. Pathogen and probiotic population data (log CFU/mL or log CFU/g) were analyzed using one-way analysis of variance (ANOVA) followed by Tukey’s Honest Significant Difference (HSD) post-hoc test using JMP Pro version 14 to determine significant differences between treatment groups. Mean values were reported with standard deviations, and statistical significance was set at p < 0.05. Data analysis and graphical presentation were performed using GraphPad Prism (version 10.0, GraphPad Software, San Diego, CA, USA). Letter groupings (A, B, C) were used on graphs to denote statistically distinct groups at the final time point.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}