
Influence of Culture Media and Environmental Factors (Water Potential and Temperature) on Mycelial Growth of Phytopythium vexans (de Bary), the Causal Agent of Dieback Disease in Apple Trees



Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Oomycete Isolates
2.2. Influence of Culture Media on Phytopythium vexans Radial Growth
2.3. Effect of Temperature on Phytopythium vexans Radial Growth
2.4. Combined Effect of Temperature and Water Potential (Ψw) on Radial Growth and Lag Time
2.5. Radial Growth and Lag Time Assessment
2.6. Statistical Analysis
3. Results
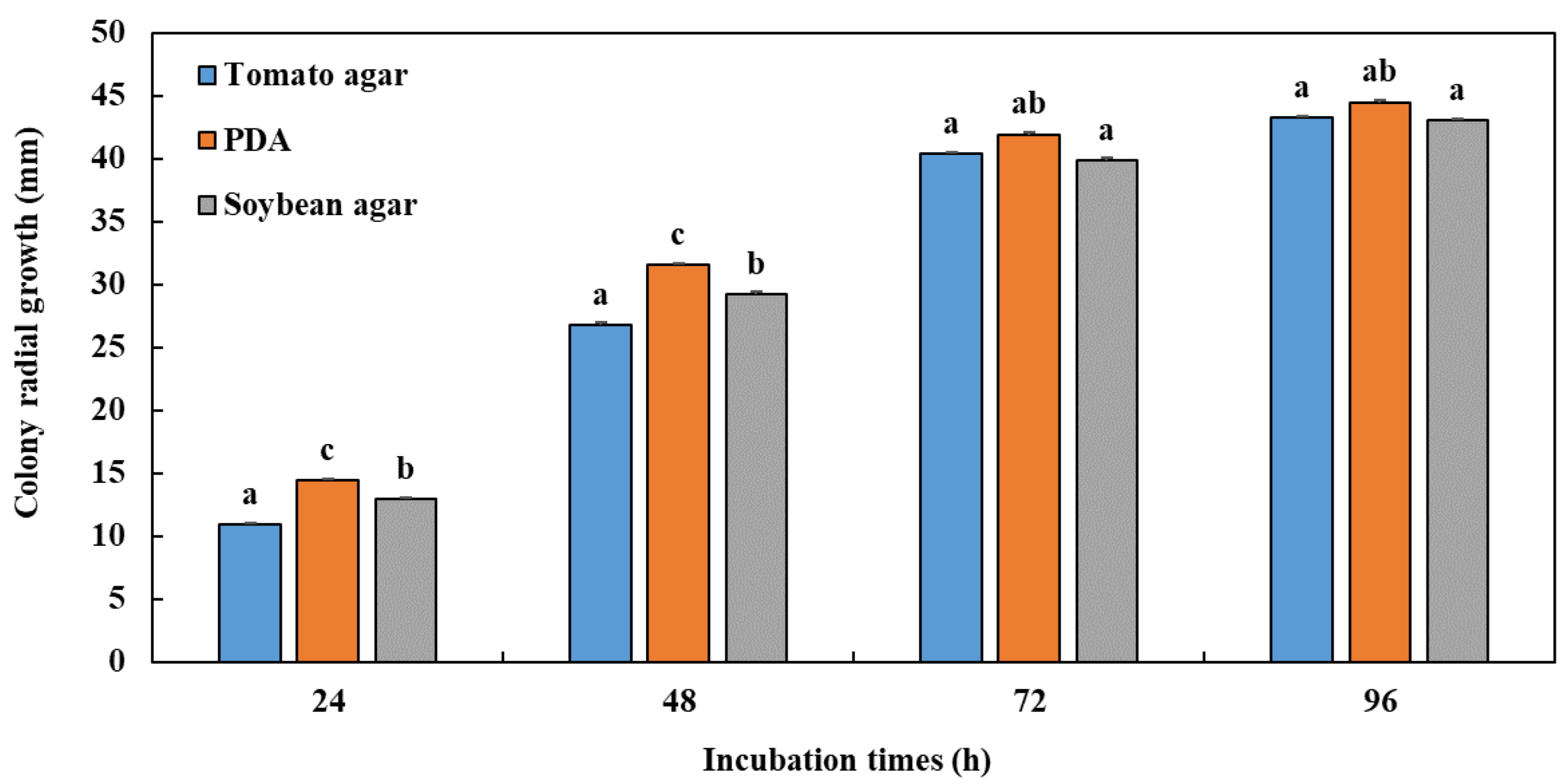
3.1. Influence of Culture Media on Radial Mycelial Growth
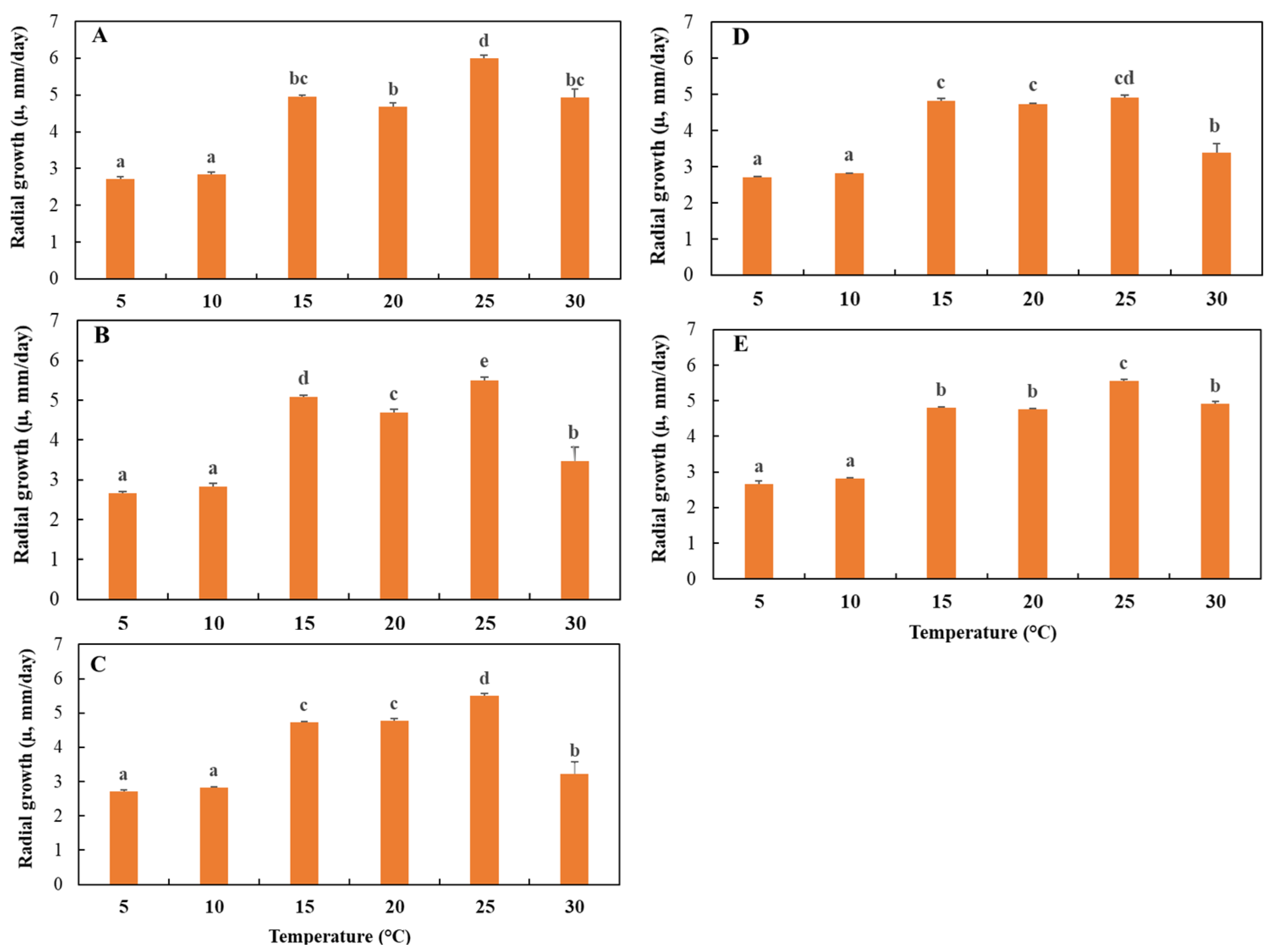
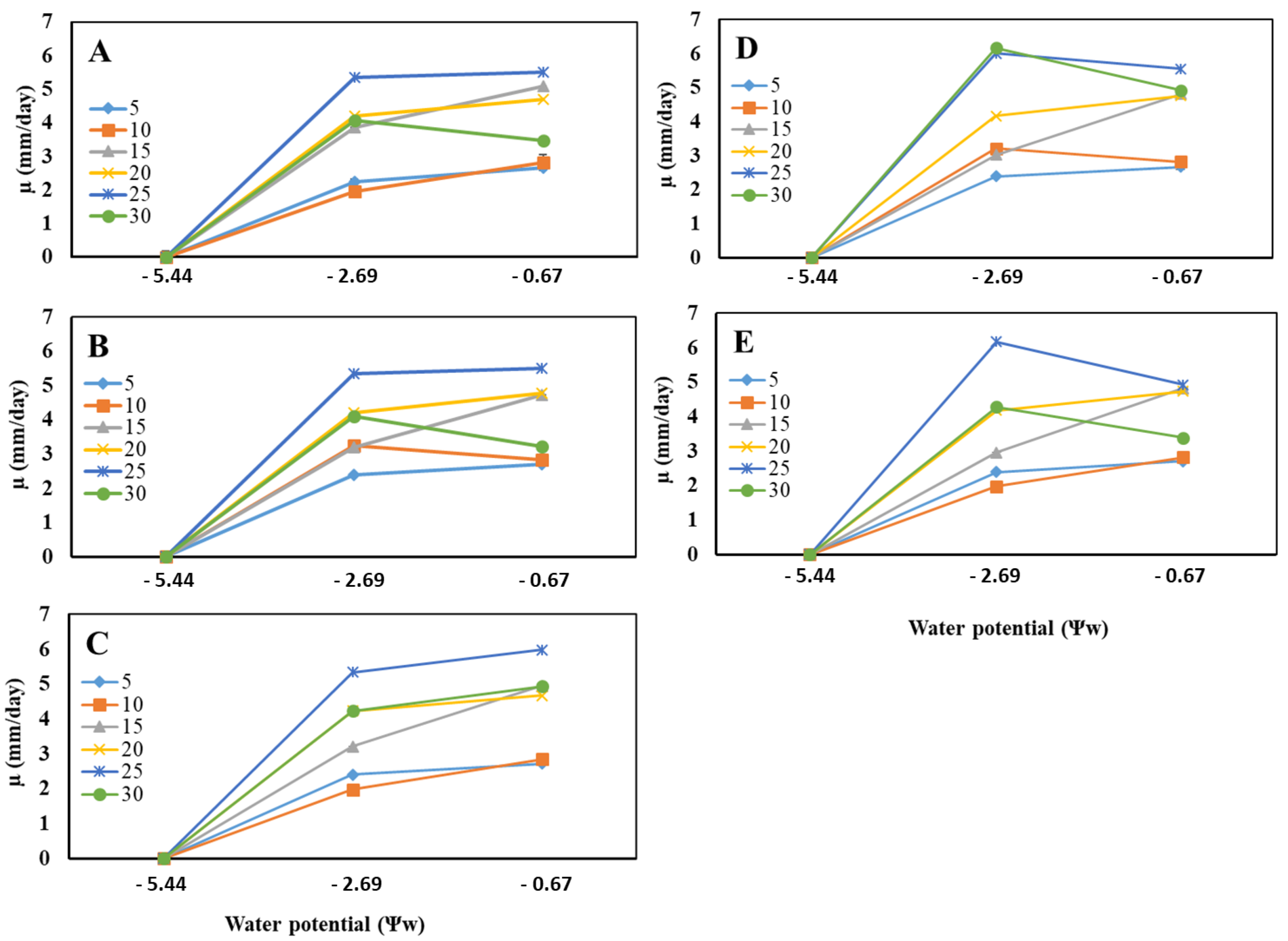
3.2. Effects of Temperature and Water Potential (Ψw) on Mycelial Radial Growth Rate
3.3. Effects of Temperature and Water Potential (Ψw) on Lag Times
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moinina, A.; Lahlali, R.; MacLean, D.; Boulif, M. Farmers’ Knowledge, Perception and Practices in Apple Pest Management and Climate Change in the Fes-Meknes Region, Morocco. Horticulturae 2018, 4, 42. [Google Scholar] [CrossRef]
- Mbovora, S.M.; Musvosvi, C.; Gasura, E. Morphological Diversity among Accessions of Apple Tree (Malus × Domestica Borkh). Adv. Agric. 2021, 2021, 7705856. [Google Scholar] [CrossRef]
- Gołębiewska, E.; Kalinowska, M.; Yildiz, G. Sustainable Use of Apple Pomace (AP) in Different Industrial Sectors. Materials 2022, 15, 1788. [Google Scholar] [CrossRef] [PubMed]
- Raada, S.; Mazouz, H.; Boulif, M. Phytosanitary practices of apple growers in the Ifrane province of the Middle Atlas of Morocco and perspectives of improvement. Pratiques phytosanitaires des pomiculteurs de la province d’Ifrane au Moyen Atlas du Maroc et perspectives d’amélioration. Rev. Maroc. Prot. Plantes 2019, 13, 19–33. [Google Scholar]
- Jabiri, S.; Bahra, C.; MacLean, D.; Radouane, N.; Barka, E.A.; Bendriss Amraoui, M.; Lahlali, R. Phytopythium vexans Associated with Apple and Pear Decline in the Saïss Plain of Morocco. Microorganisms 2021, 9, 1916. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, B.; Gold, C.; Kashaija, I.; Ssali, H.; Night, G.; Bwamiki, D. Effects of legume intercrops on soil-borne pests, biomass, nutrients and soil water in banana. Biol. Fertil. Soils 2001, 34, 342–348. [Google Scholar] [CrossRef]
- Lehtijärvi, A.; Aday Kaya, A.G.; Woodward, S.; Jung, T.; Doğmuş Lehtijärvi, H.T. Oomycota species associated with deciduous and coniferous seedlings in forest tree nurseries of Western Turkey. For. Pathol. 2017, 47, e12363. [Google Scholar] [CrossRef]
- Brasier, C.; Scanu, B.; Cooke, D.; Jung, T. Phytophthora: An ancient, historic, biologically and structurally cohesive and evolutionarily successful generic concept in need of preservation. IMA Fungus 2022, 13, 12. [Google Scholar] [CrossRef]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A Molecular Phylogeny of Phytophthora and Related Oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- Richards, T.A.; Talbot, N.J. Plant Parasitic Oomycetes Such as Phytophthora Species Contain Genes Derived from Three Eukaryotic Lineages. Plant Signal. Behav. 2007, 2, 112–114. [Google Scholar] [CrossRef][Green Version]
- Lang-Yona, N.; Pickersgill, D.A.; Maurus, I.; Teschner, D.; Wehking, J.; Thines, E.; Pöschl, U.; Després, V.R.; Fröhlich-Nowoisky, J. Species Richness, rRNA Gene Abundance, and Seasonal Dynamics of Airborne Plant-Pathogenic Oomycetes. Front. Microbiol. 2018, 9, 2673. [Google Scholar] [CrossRef] [PubMed]
- Sewell, G.W.F. Effects of Pythium species on the growth of apple and their possible causal role in apple replant disease. Ann. Appl. Biol. 1981, 97, 31–42. [Google Scholar] [CrossRef]
- Harris, D.C. The Phytophthora diseases of apple. J. Hortic. Sci. 1991, 66, 513–544. [Google Scholar] [CrossRef]
- Bala, K.; Robideau, G.P.; Desaulniers, N.; de Cock, A.W.; Levesque, C.A. Taxonomy, DNA barcoding and phylogeny of three new species of Pythium from Canada. Persoonia 2010, 25, 22–31. [Google Scholar] [CrossRef]
- Nam, B.; Choi, Y.-J. Phytopythium and Pythium Species (Oomycota) Isolated from Freshwater Environments of Korea. Mycobiology 2019, 47, 261–272. [Google Scholar] [CrossRef]
- Jabiri, S.; Lahlali, R.; Bendriss Amraoui, M.; Tahiri, A.; Amiri, S. First report of Phytopythium vexans associated with dieback disease of apple trees in Morocco. J. Plant Pathol. 2020, 102, 1319. [Google Scholar] [CrossRef]
- Rodriguez-Padron, C.; Siverio, F.; Perez-Sierra, A.; Rodriguez, A. Isolation and pathogenicity of Phytophthora species and Phytopythiumvexans recovered from avocado orchards in the Canary Islands, including Phytophthoraniederhauserii as a new pathogen of avocado. Phytopathol. Mediterr. 2018, 57, 89–106. [Google Scholar]
- Polat, Z.; Awan, Q.N.; Hussain, M.; Akgül, D.S. First Report of Phytopythium vexans Causing Root and Collar Rot of Kiwifruit in Turkey. Plant Dis. 2017, 101, 1058. [Google Scholar] [CrossRef]
- Williamson-Benavides, B.A.; Dhingra, A. Understanding Root Rot Disease in Agricultural Crops. Horticulturae 2021, 7, 33. [Google Scholar] [CrossRef]
- Rodríguez-Padrón, C.; Rodríguez, A.; Siverio, F. Survey in Nurseries and Irrigation Water Reservoirs as Sources of Oomycetes Found in Avocado Orchards in the Canary Islands. Plant Dis. 2019, 103, 1264–1274. [Google Scholar] [CrossRef]
- Jones, L.A.; Worobo, R.W.; Smart, C.D.; Elkins, C.A. Plant-Pathogenic Oomycetes, Escherichia coli Strains, and Salmonella spp. Frequently Found in Surface Water Used for Irrigation of Fruit and Vegetable Crops in New York State. Appl. Environ. Microbiol. 2014, 80, 4814–4820. [Google Scholar] [CrossRef] [PubMed]
- Redekar, N.R.; Bourret, T.B.; Eberhart, J.L.; Johnson, G.E.; Pitton, B.J.L.; Haver, D.L.; Oki, L.R.; Parke, J.L. The population of oomycetes in a recycled irrigation water system at a horticultural nursery in southern California. Water Res. 2020, 183, 116050. [Google Scholar] [CrossRef] [PubMed]
- Nam, B.; Lee, D.-J.; Choi, Y.-J. High-Temperature-Tolerant Fungus and Oomycetes in Korea, Including Saksenaea longicolla sp. nov. Mycobiology 2021, 49, 476–490. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, S.; Furzer, O.; Jones, J.D.G.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.D.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabières, F.; et al. The Top 10 oomycete pathogens in molecular plant pathology. Mol. Plant Pathol. 2015, 16, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Vannini, A.; Breccia, M.; Bruni, N.; Tomassini, A.; Vettraino, A.M. Behaviour and survival of Phytophthora cambivora inoculum in soil-like substrate under different water regimes. For. Pathol. 2012, 42, 362–370. [Google Scholar] [CrossRef]
- Halsall, D.M.; Williams, J.D. Effect of Root Temperature on the Development of Phytophthora cinnamomi Root Rot in Eucalyptus Seedlings. Aust. J. Bot. 1984, 32, 521–528. [Google Scholar] [CrossRef]
- Martin, F.N.; Loper, J.E. Soilborne Plant Diseases Caused by Pythium Spp.: Ecology, Epidemiology, and Prospects for Biological Control. Crit. Rev. Plant Sci. 1999, 18, 111–181. [Google Scholar] [CrossRef]
- de Cock, A.W.A.M.; Lodhi, A.M.; Rintoul, T.L.; Bala, K.; Robideau, G.P.; Abad, Z.G.; Coffey, M.D.; Shahzad, S.; Lévesque, C.A. Phytopythium: Molecular phylogeny and systematics. Persoonia 2015, 34, 25–39. [Google Scholar] [CrossRef]
- Baten, M.A.; Asano, T.; Motohashi, K.; Ishiguro, Y.; Rahman, M.Z.; Inaba, S.; Suga, H.; Kageyama, K. Phylogenetic relationships among Phytopythium species, and re-evaluation of Phytopythium fagopyri comb. nov., recovered from damped-off buckwheat seedlings in Japan. Mycol. Prog. 2014, 13, 1003. [Google Scholar] [CrossRef]
- Prencipe, S.; Savian, F.; Nari, L.; Ermacora, P.; Spadaro, D.; Martini, M. First Report of Phytopythium vexans causing decline syndrome of Actinidia deliciosa ‘Hayward’ in Italy. Plant Dis. 2020, 104, 2032. [Google Scholar] [CrossRef]
- Thao, L.D.; Hien, L.T.; Liem, N.V.; Thanh, H.M.; Khanh, T.N.; Binh, V.T.P.; Trang, T.T.T.; Anh, P.T.; Tu, T.T. First report of Phytopythium vexans causing root rot disease on durian in Vietnam. New Dis. Rep. 2020, 41, 2. [Google Scholar] [CrossRef]
- Boari, A.D.J.; Cunha, E.M.; Quadros, A.F.F.; Barreto, R.W.; Fernandes, A.F. First Report of Phytopythium sp. Causing Storage Root Rot and Foliage Blight of Cassava in Brazil. Plant Dis. 2018, 102, 1042–1104. [Google Scholar] [CrossRef]
- Wielgoss, A.; Nechwatal, J.; Bogs, C.; Mendgen, K. Host plant development, water level and water parameters shape Phragmites australis-associated oomycete communities and determine reed pathogen dynamics in a large lake. FEMS Microbiol. Ecol. 2009, 69, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Derevnina, L.; Petre, B.; Kellner, R.; Dagdas, Y.F.; Sarowar, M.N.; Giannakopoulou, A.; De la Concepcion, J.C.; Chaparro-Garcia, A.; Pennington, H.G.; van West, P.; et al. Emerging oomycete threats to plants and animals. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150459. [Google Scholar] [CrossRef] [PubMed]
- Velásquez, A.C.; Castroverde, C.D.M.; He, S.Y. Plant–Pathogen Warfare under Changing Climate Conditions. Curr. Biol. 2018, 28, R619–R634. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Serrhini, M.N.; Friel, D.; Jijakli, M.H. Predictive modelling of temperature and water activity (solutes) on the in vitro radial growth of Botrytis cinerea Pers. Int. J. Food Microbiol. 2007, 114, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Serrhini, M.N.; Friel, D.; Jijakli, M.H. In vitro effects of water activity, temperature and solutes on the growth rate of P. italicum Wehmer and P. digitatum Sacc. J. Appl. Microbiol. 2006, 101, 628–636. [Google Scholar] [CrossRef]
- Lahlali, R.; Bajji, M.; Serrhini, M.N.; Jijakli, M.H. Modelling the effect of temperature, Water activity and solute on the in vitro growth of the biocontrol yeast Pichia anomala strain K. Biotechnol. Agron. Soc. Environ. 2008, 12, 353–359. [Google Scholar]
- Lahlali, R.; Serrhini, M.N.; Jijakli, M.H. Studying and modelling the combined effect of temperature and water activity on the growth rate of P. expansum. Int. J. Food Microbiol. 2005, 103, 315–322. [Google Scholar] [CrossRef]
- Romero, S.M.; Patriarca, A.; Fernández Pinto, V.; Vaamonde, G. Effect of water activity and temperature on growth of ochratoxigenic strains of Aspergillus carbonarius isolated from Argentinean dried vine fruits. Int. J. Food Microbiol. 2007, 115, 140–143. [Google Scholar] [CrossRef]
- Patriarca, A.; Vaamonde, G.; Fernández Pinto, V.; Comerio, R. Influence of water activity and temperature on the growth of Wallemia sebi: Application of a predictive model. Int. J. Food Microbiol. 2001, 68, 61–67. [Google Scholar] [CrossRef]
- Weiland, J.E.; Santamaria, L.; Grünwald, N.J. Sensitivity of Pythium irregulare, P. sylvaticum, and P. ultimum from Forest Nurseries to Mefenoxam and Fosetyl-Al, and Control of Pythium Damping-off. Plant Dis. 2014, 98, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Hord, M.J.; Ristaino, J.B. Effects of Physical and Chemical Factors on the Germination of Oospores of Phytophthora capsici in vitro. Phytopathology 1991, 81, 1541–1546. [Google Scholar] [CrossRef]
- Delmas, C.E.L.; Mazet, I.D.; Jolivet, J.; Delière, L.; Delmotte, F. Simultaneous quantification of sporangia and zoospores in a biotrophic oomycete with an automatic particle analyzer: Disentangling dispersal and infection potentials. J. Microbiol. Methods 2014, 107, 169–175. [Google Scholar] [CrossRef]
- Shivakumar, K.S.; Somasekhara, Y.M. Influence of Culture Media on Growth and Sporulation of Phytophthora capsici the Cause of Quick Wilt of Black Pepper. Mysore J. Agric. Sci. 2018, 52, 369–374. [Google Scholar]
- Medina, M.V.; Platt, H.W. Comparison of different culture media on the mycelial growth, sporangia and oospore production ofPhytophthora infestans. Am. J. Potato Res. 1999, 76, 121–125. [Google Scholar] [CrossRef]
- de Sousa Silva, J.L.; Ishida, A.K.N.; Cunha, R.L.; Lima, A.M.; Moura, E.F. Culture medium and inoculation methodology for the study of soft root rot caused by Phytopythium sp. Cienc. Rural 2019, 49, 1–5. [Google Scholar] [CrossRef]
- Cantrell, H.F.; Dowler, W.M. Effects of Temperature and ph on Growth and Composition of Pythium Irregulare and Pythium Vexans. Mycologia 1971, 63, 31–37. [Google Scholar] [CrossRef]
- Agustí-Brisach, C.; Armengol, J. Effects of temperature, pH and water potential on mycelial growth, sporulation and chlamydospore production in culture of Cylindrocarpon spp. associated with black foot of grapevines. Phytopathol. Mediterr. 2012, 51, 37–50. [Google Scholar] [CrossRef]
- Krajaejun, T.; Chongtrakool, P.; Angkananukul, K.; Brandhorst, T. Effect of temperature on growth of the pathogenic oomycete Pythium insidiosum. Southeast Asian J. Trop. Med. Public Health 2010, 41, 1462–1466. [Google Scholar]
- Bolton, A.T. Effects of temperature and pH of soilless media on root rot of poinsettia caused by Pythium aphanidermatum. Can. J. Plant Pathol. 1980, 2, 83–85. [Google Scholar] [CrossRef]
- Wong, D.; Sivasithamparam, K.; Barbetti, M. Influence of environmental factors on the growth and survival of Phytophthora clandestina. Can. J. Microbiol. 2011, 32, 553–556. [Google Scholar] [CrossRef]
- Cook, R.J.; Duniway, J.M. Water Relations in the Life-cycles of Soilborne Plant Pathogens. Water Potential Relat. Soil Microbiol. 1981, 9, 119–139. [Google Scholar]
- Duniway, J.M. Water Relations of Water Molds. Annu. Rev. Phytopathol. 1979, 17, 431–460. [Google Scholar] [CrossRef]
- Gill, J.S.; Sivasithamparam, K.; Smettem, K.R.J. Effect of soil moisture at different temperatures on Rhizoctonia root rot of wheat seedlings. Plant Soil 2001, 231, 91–96. [Google Scholar] [CrossRef]
- Malajczuk, N.; Theodorou, C. Influence of water potential on growth and cultural characteristics of Phytophthora cinnamomi. Trans. Br. Mycol. Soc. 1979, 72, 15–18. [Google Scholar] [CrossRef]
- Harris, R.F.; Dalton, F.N.; Gardner, W.R. Water potential relations of three root-infecting Phytophthora species. Phytopathology 1970, 60, 932–934. [Google Scholar]
- Shew, H.D. Effects of Soil Matric Potential on Infection of Tobacco by Phytophthora parasitica var. nicotianae. Phytopathology 1983, 73, 1160. [Google Scholar] [CrossRef]
- Hord, M.J. Effect of the Matric Component of Soil Water Potential on Infection of Pepper Seedlings in Soil Infested with Oospores of Phytophthora capsici. Phytopathology 1992, 82, 792. [Google Scholar] [CrossRef]
- Begoude, B.A.D.; Lahlali, R.; Friel, D.; Tondje, P.R.; Jijakli, M.H. Response surface methodology study of the combined effects of temperature, pH, and aw on the growth rate of Trichoderma asperellum. J. Appl. Microbiol. 2007, 103, 845–854. [Google Scholar] [CrossRef]
- Judelson, H.S.; Ah-Fong, A.M. Exchanges at the Plant-Oomycete Interface That Influence Disease. Plant Physiol. 2019, 179, 1198–1211. [Google Scholar] [CrossRef] [PubMed]
- Bassani, I.; Larousse, M.; Tran, Q.D.; Attard, A.; Galiana, E. Phytophthora zoospores: From perception of environmental signals to inoculum formation on the host-root surface. Comput. Struct. Biotechnol. J. 2020, 18, 3766–3773. [Google Scholar] [CrossRef] [PubMed]
- Pratt, R.G. Interrelationships of Seedling Age, Inoculum, Soil Moisture Level, Temperature, and Host and Pathogen Genotype in Phytophthora Root Rot of Alfalfa. Phytopathology 1976, 66, 81. [Google Scholar] [CrossRef]
- Sterne, R.E. The Effect of Matric and Osmotic Potential of Soil on Phytophthora Root Disease of Persea indica. Phytopathology 1977, 77, 1491. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source | DF | Mean Square | F-Value | Pr > F |
|---|---|---|---|---|
| Species | 4 | 0.698 | 50.554 | 0.000 |
| Temperature | 5 | 26.855 | 1944.162 | 0.000 |
| aw | 2 | 465.160 | 33,675.175 | 0.000 |
| Species × Temperature | 20 | 0.530 | 38.377 | 0.000 |
| Species × aw | 8 | 0.507 | 36.670 | 0.000 |
| Temperature × aw | 10 | 8.816 | 638.267 | 0.000 |
| Species × Temperature × aw | 40 | 0.316 | 22.856 | 0.000 |
| T | Ψw | I5 | E4 | M2 | S2 | A7 |
|---|---|---|---|---|---|---|
| 5 | 0.995 | 2.72 ± 0.05 a | 2.66 ± 0.04 b | 2.70 ± 0.06 c | 2.70 ± 0.03 c | 2.66 ± 0.08 b |
| 0.98 | 2.40 ± 0.02 c | 2.25 ± 0.24 c | 2.39 ± 0.06 b | 2.39 ± 0.03 cd | 2.40 ± 0.02 bc | |
| 0.96 | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | |
| 10 | 0.995 | 2.84 ± 0.05 d | 2.83 ± 0.07 b | 2.83 ± 0.03 c | 2.87 ± 0.01 d | 2.81 ± 0.02 bcd |
| 0.98 | 1.98 ± 0.07 b | 1.97 ± 0.05 c | 3.26 ± 0.04 d | 1.98 ± 0.03 b | 3.21 ± 0.01 cd | |
| 0.96 | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | |
| 15 | 0.995 | 4.95 ± 0.04 e | 5.09 ± 0.04 e | 4.73 ± 0.02 f | 4.83 ± 0.05 d | 4.81 ± 0.01 f |
| 0.98 | 3.21 ± 0.07 h | 3.87 ± 0.19 g | 3.19 ± 0.10 d | 2.96 ± 0.03 g | 3.02 ± 0.05 cd | |
| 0.96 | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | |
| 20 | 0.995 | 4.67 ± 0.12 g | 4.69 ± 0.08 f | 4.77 ± 0.06 f | 4.74 ± 0.01 g | 4.76 ± 0.02 f |
| 0.98 | 4.23 ± 0.06 f | 4.21 ± 0.05 e | 4.19 ± 0.03 e | 4.18 ± 0.02 f | 4.17 ± 0.01 e | |
| 0.96 | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | |
| 25 | 0.995 | 5.99 ± 0.09 i | 5.51 ± 0.07 h | 5.51 ± 0.07 h | 4.92 ± 0.07 g | 5.55 ± 0.08 g |
| 0.98 | 5.34 ± 0.13 j | 5.35 ± 0.09 gh | 5.35 ± 0.09 h | 6.17 ± 0.37 f | 6.01 ± 0.54 gh | |
| 0.96 | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | |
| 30 | 0.995 | 4.93 ± 0.23 h | 3.47 ± 0.35 e | 3.22 ± 0.36 d | 3.38 ± 0.25 e | 4.92 ± 0.07 f |
| 0.98 | 4.23 ± 0.06 h | 4.08 ± 0.13 d | 4.10 ± 0.11 e | 4.28 ± 0.12 f | 6.17 ± 0.37 h | |
| 0.96 | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Temperature | Ψw | I5 λ ± SD | E4 λ ± SD | M2 λ ± SD | S2 λ ± SD | A7 λ ± SD |
|---|---|---|---|---|---|---|
| 5 | 0.995 | 5.158 ± 0.17 h | 4.99 ± 0.09 g | 5.01 ± 0.14 f | 5.10 ± 0.077 i | 5.01 ± 0.20 g |
| 0.98 | 5.56 ± 0.04 i | 5.44 ± 0.13 h | 5.53 ± 0.05 f | 5.53 ± 0.04 j | 5.54 ± 0.04 h | |
| 0.96 | >20 a | >20 a | >20 a | >20 a | >20 a | |
| 10 | 0.995 | 2.55 ± 0.17 f | 2.54 ± 0.19 e | 2.45 ± 0.11 d | 2.48 ± 0.10 g | 2.40 ± 0.06 e |
| 0.98 | 3.13 ± 0.23 g | 3.02 ± 0.13 f | 3.17 ± 0.05 e | 3.11 ± 0.16 h | 3.10 ± 0.03 f | |
| 0.96 | >20 a | >20 a | >20 a | >20 a | >20 a | |
| 15 | 0.995 | 1.23 ± 0.06 d | 1.08 ± 0.04 c | 1.66 ± 0.01 c | 1.52 ± 0.06 e | 1.53 ± 0.02 d |
| 0.98 | 1.86 ± 0.27 e | 1.07 ± 0.03 c | 1.59 ± 0.45 c | 1.98 ± 0.23 f | 1.53 ± 0.07 d | |
| 0.96 | >20 a | >20 a | >20 a | >20 a | >20 a | |
| 20 | 0.995 | 1.23 ± 0.01 d | 1.22 ± 0.06 c | 1.22 ± 0.17 c | 1.04 ± 0.03 c | 1.01 ± 0.02 c |
| 0.98 | 1.80 ± 0.51 e | 1.83 ± 0.08 d | 1.58 ± 0.40 c | 1.50 ± 0.43 d | 1.69 ± 0.03 d | |
| 0.96 | >20 a | >20 a | >20 a | >20 a | >20 a | |
| 25 | 0.995 | 0.90 ± 0.03 cd | 1.08 ± 0.14 c | 1.11 ± 0.03 bc | 1.51 ± 0.18 c | 1.05 ± 0.06 c |
| 0.98 | 1.15 ± 0.02 d | 1.32 ± 0.32 d | 1.55 ± 0.32 c | 1.10 ± 0.06 cd | 1.14 ± 0.03 c | |
| 0.96 | >20 a | >20 a | >20 a | >20 a | >20 a | |
| 30 | 0.995 | 0.23 ± 0.03 ab | 0.18 ±0.021 b | 0.11 ± 0.025 a | 0.12 ± 0.02 ab | 0.38 ± 0.39 b |
| 0.98 | 0.60 ± 0.29 bc | 0.57 ± 0.14 c | 0.66 ± 0.02 b | 0.50 ± 0.17 b | 0.42 ± 0.14 b | |
| 0.96 | >20 a | >20 a | >20 a | >20 a | >20 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jabiri, S.; El Hamss, H.; Amraoui, M.B.; Lahlali, R. Influence of Culture Media and Environmental Factors (Water Potential and Temperature) on Mycelial Growth of Phytopythium vexans (de Bary), the Causal Agent of Dieback Disease in Apple Trees. Appl. Microbiol. 2022, 2, 861-872. https://doi.org/10.3390/applmicrobiol2040066
Jabiri S, El Hamss H, Amraoui MB, Lahlali R. Influence of Culture Media and Environmental Factors (Water Potential and Temperature) on Mycelial Growth of Phytopythium vexans (de Bary), the Causal Agent of Dieback Disease in Apple Trees. Applied Microbiology. 2022; 2(4):861-872. https://doi.org/10.3390/applmicrobiol2040066
Chicago/Turabian StyleJabiri, Salma, Hajar El Hamss, Mohammed Bendriss Amraoui, and Rachid Lahlali. 2022. "Influence of Culture Media and Environmental Factors (Water Potential and Temperature) on Mycelial Growth of Phytopythium vexans (de Bary), the Causal Agent of Dieback Disease in Apple Trees" Applied Microbiology 2, no. 4: 861-872. https://doi.org/10.3390/applmicrobiol2040066
APA StyleJabiri, S., El Hamss, H., Amraoui, M. B., & Lahlali, R. (2022). Influence of Culture Media and Environmental Factors (Water Potential and Temperature) on Mycelial Growth of Phytopythium vexans (de Bary), the Causal Agent of Dieback Disease in Apple Trees. Applied Microbiology, 2(4), 861-872. https://doi.org/10.3390/applmicrobiol2040066