
Degradation of Polycyclic Aromatic Hydrocarbons by Co-Culture of Pleurotus ostreatus Florida and Azospirillum brasilense



Abstract
1. Introduction
2. Materials and Methods
3. Results
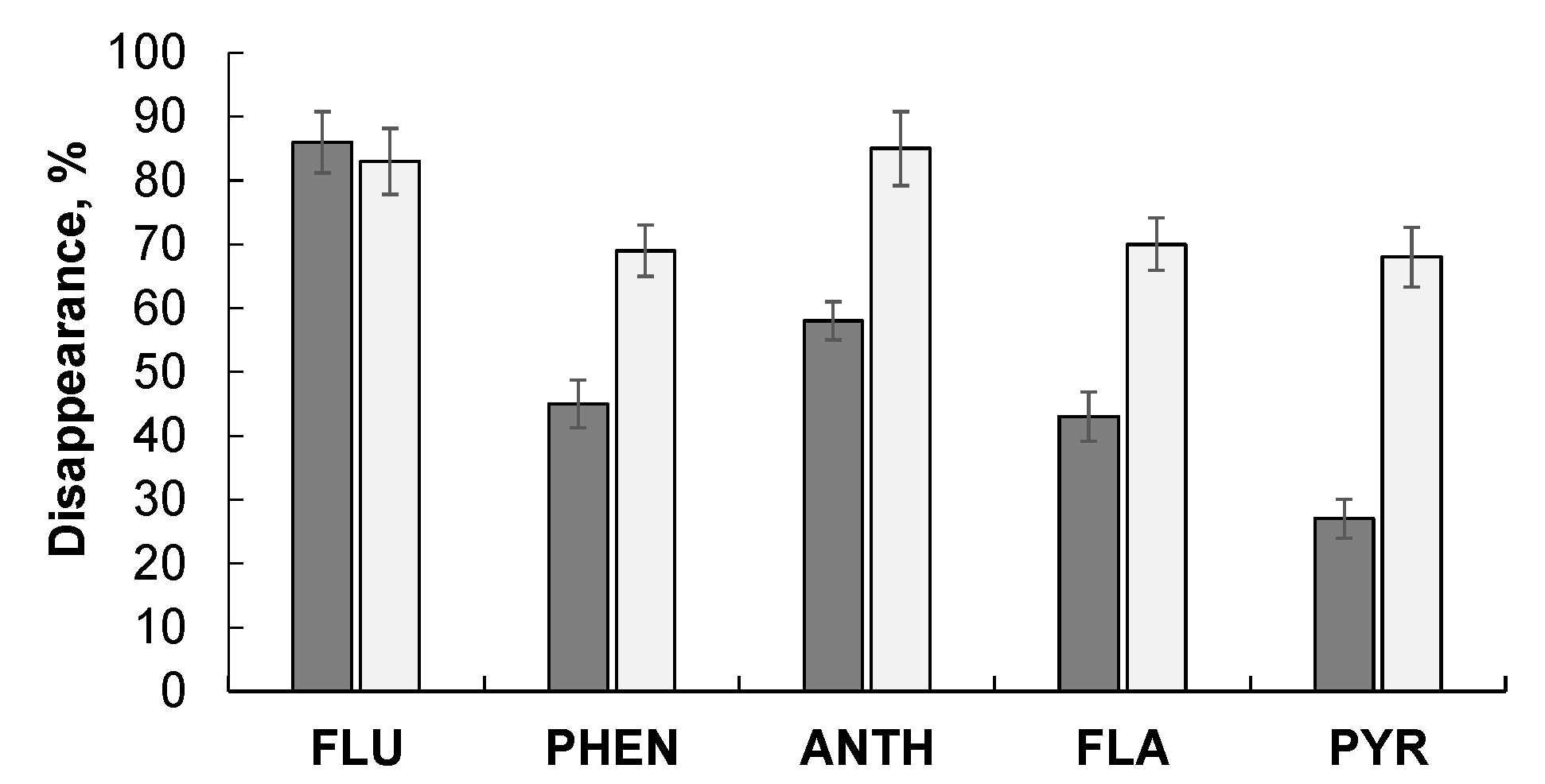
3.1. Degradation of the PAH Mixture
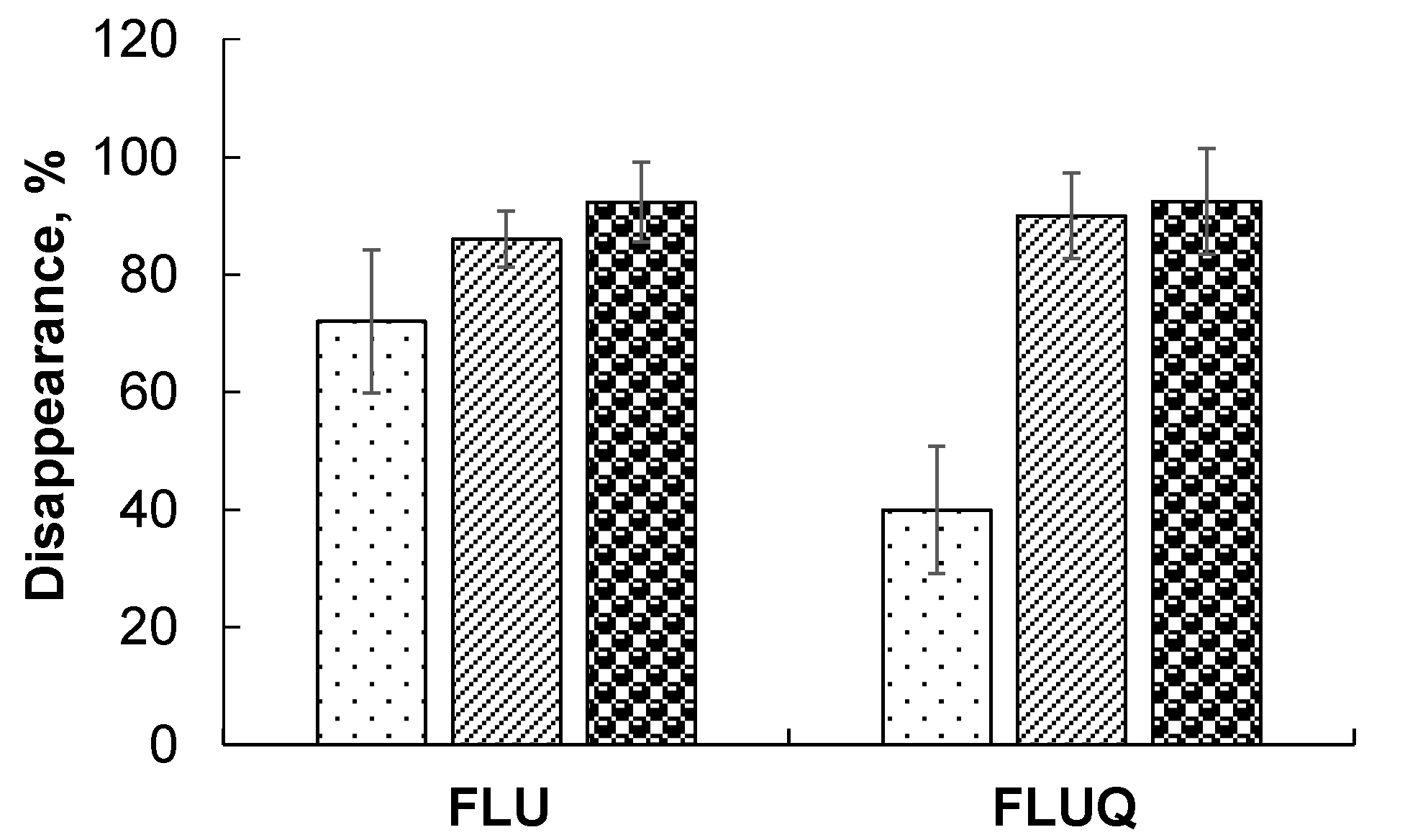
3.2. Degradation of Fluorene and Its Main Metabolite, 9-Fluorenone
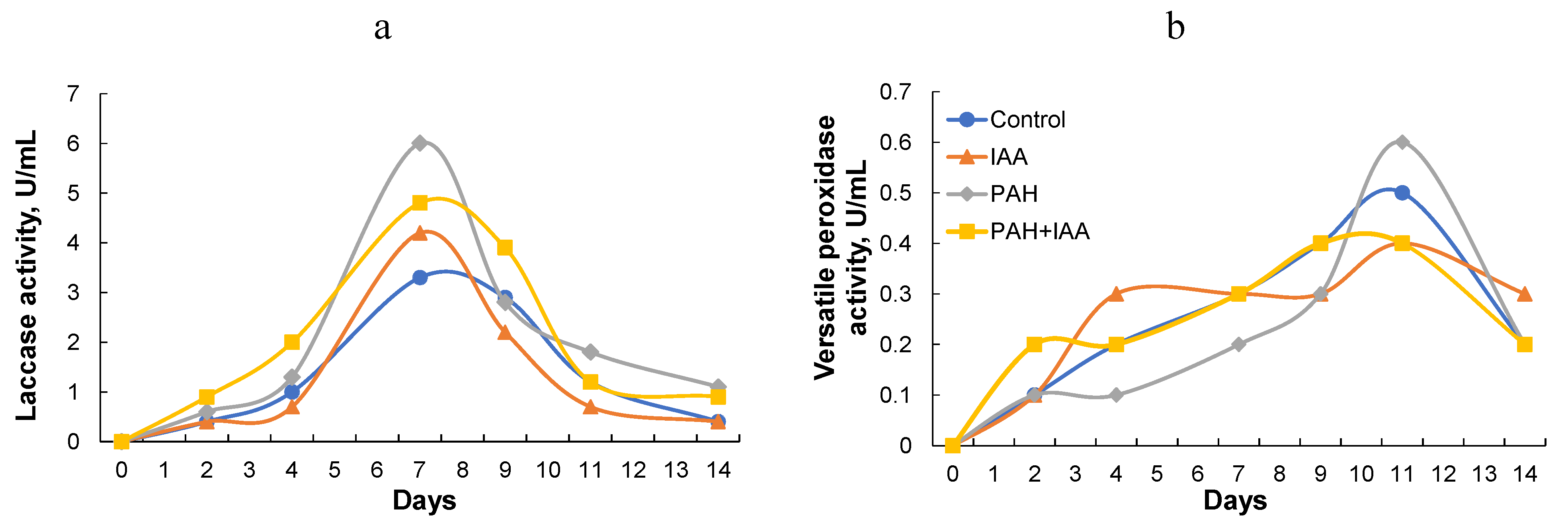
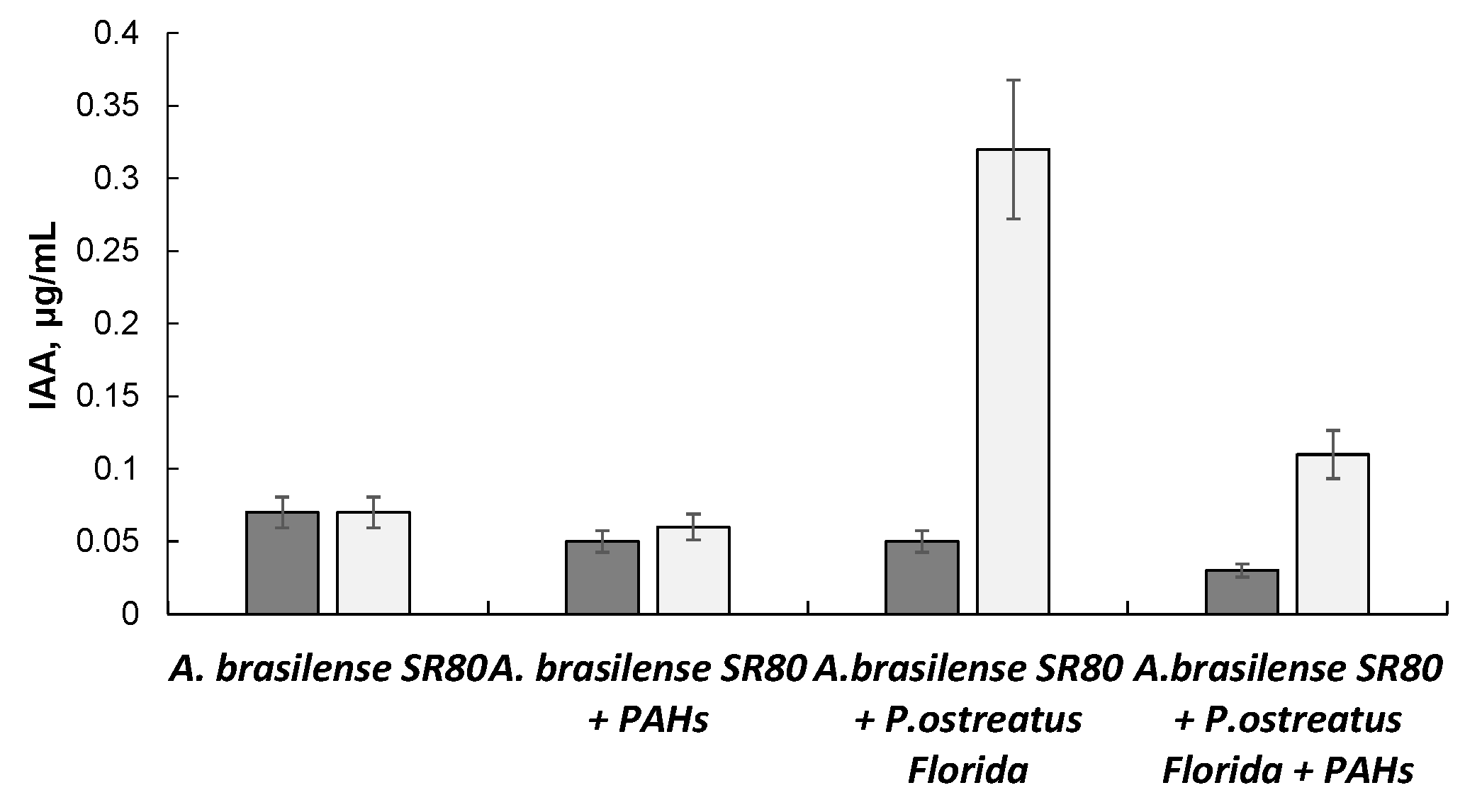
3.3. Effect of IAA on Fungal Growth, Enzyme Activity, and PAH Degradation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wargo, M.; Hogan, D. Fungal-bacterial interactions: A mixed bag of mingling microbes. Curr. Opin. Microbiol. 2006, 9, 359–364. [Google Scholar] [CrossRef]
- de Boer, W. Upscaling of fungal-bacterial interactions: From the lab to the field. Curr. Opin. Microbiol. 2017, 37, 35–41. [Google Scholar] [CrossRef]
- Kohlmeier, S.; Smits, T.H.M.; Ford, R.M.; Keel, C.; Harms, H.; Wick, L.Y. Taking the fungal highway: Mobilization of pollutant-degrading bacteria by fungi. Environ. Sci. Technol. 2005, 39, 4640–4646. [Google Scholar] [CrossRef]
- van Overbeek, L.S.; Saikkonen, K. Impact of bacterial-fungal interactions on the colonization of the endosphere. Trends Plant Sci. 2016, 21, 230–242. [Google Scholar] [CrossRef]
- Furuno, S.; Päzolt, K.; Rabe, K.; Neu, T.R.; Harms, H.; Wick, L.Y. Fungal mycelia allow chemotactic dispersal of polycyclic aromatic hydrocarbon-degrading bacteria in water-unsaturated systems. Environ. Microbiol. 2010, 12, 1391–1398. [Google Scholar] [CrossRef]
- Minerdi, D.; Bianciotto, V.; Bonfante, P. Endosymbiotic bacteria in mycorrhizal fungi: From their morphology to genomic sequences. Plant Soil 2002, 244, 211–219. [Google Scholar] [CrossRef]
- Jambon, I.; Thijs, S.; Weyens, N.; Vangronsveld, J. Harnessing plant-bacteria-fungi interactions to improve plant growth and degradation of organic pollutants. J. Plants Interact. 2018, 13, 119–130. [Google Scholar] [CrossRef]
- Kim, M.K.; Math, R.K.; Cho, K.M.; Shin, K.J.; Kim, J.O.; Ryu, J.S.; Lee, Y.H.; Yun, H.D. Effect of Pseudomonas sp. P7014 on the growth of edible mushroom Pleurotus eringii in bottle culture for commercial production. Bioresour. Technol. 2008, 99, 3306–3308. [Google Scholar] [CrossRef]
- Fu, S.-F.; Wei, J.-Y.; Chen, H.-W.; Liu, Y.-Y.; Lu, H.-Y.; Chou, J.-Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal. Behav. 2015, 10, e1048052. [Google Scholar] [CrossRef]
- Pham, M.T.; Huang, C.-M.; Kirschner, R. The plant growth-promoting potential of the mesophilic wood-rot mushroom Pleurotus pulmonarius. J. Appl. Microbiol. 2019, 127, 1157–1171. [Google Scholar] [CrossRef]
- McErlean, C.; Marchant, R.; Banat, I. An evaluation of soil colonization potential of selected fungi and their production of ligninolytic enzymes for use in soil bioremediation applications. Antonie Van Leeuwenhoek 2006, 90, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, M.; Cajthaml, T.; Šašek, V. Mycoremediation of PAH-contaminated soil. Folia Microbiol. 2002, 47, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Thion, C.; Cebron, A.; Beguiristain, T.; Leyval, C. Inoculation of PAH-degrading strains of Fusarium solani and Arthrobacter oxydans in rhizospheric sand and soil microcosms: Microbial interactions and PAH dissipation. Biodegradation 2013, 24, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Nomura, N.; Nakajima, T.; Uchiyama, H. Case study of the relationship between fungi and bacteria associated with high-molecular-weight polycyclic aromatic hydrocarbon degradation. J. Biosci. Bioeng. 2012, 113, 624–630. [Google Scholar] [CrossRef]
- Zafra, G.; Absalon, A.; Cuevas, M.; Cortes-Espinosa, D. Isolation and selection of a highly tolerant microbial consortium with potential for PAH biodegradation from heavy crude oil-contaminated soil. Water Air Soil Pollut. 2014, 225, 1826. [Google Scholar] [CrossRef]
- Ma, X.; Ding, N.; Peterson, E.C.; Daugulis, A.J. Heavy metals species affect fungal-bacterial synergism during the bioremediation of fluoranthene. Appl. Microbiol. Biotechnol. 2016, 100, 7741–7750. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Das, A.; Palaniswamy, M.; Angayarkanni, J. Degradation of benzo[a]pyrene by Pleurotus ostreatus PO-3 in the presence of defined fungal and bacterial co-cultures. J. Basic Microbiol. 2017, 57, 95–103. [Google Scholar] [CrossRef]
- Boonchan, S.; Britz, M.L.; Stanley, G.A. Degradation and mineralization of high-molecular-weight polycyclic aromatic hydrocarbons by defined fungal-bacterial cocultures. Appl. Environ. Microbiol. 2000, 66, 1007–1019. [Google Scholar] [CrossRef]
- Kim, J.; Lee, C. Microbial degradation of polycyclic aromatic hydrocarbon in soil by bacterium-fungus co-cultures. Biotechnol. Bioprocess Eng. 2007, 12, 410–416. [Google Scholar] [CrossRef]
- Brodkorb, T.; Legge, R. Enhanced biodegradation of phenanthrene in oil tar-contaminated soils supplemented with Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1992, 58, 3117–3121. [Google Scholar] [CrossRef]
- Zeng, J.; Lin, X.; Zhang, J.; Zhu, H.; Chen, H.; Wong, M.H. Successive transformation of benzo[a]pyrene by laccase of Trametes versicolor and pyrene-degrading Mycobacterium strains. Appl. Microbiol. Biotechnol. 2013, 97, 3183–3194. [Google Scholar] [CrossRef] [PubMed]
- Kadri, T.; Rouissi, T.; Brar, S.K.; Cledon, M.; Sarma, S.; Verma, M. Biodegradation of polycyclic aromatic hydrocarbons (PAHs) by fungal enzymes: A review. J. Environ. Sci. 2017, 51, 52–74. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Kumar, M. Biodegradation of polycylic aromatic hydrocarbons (PAHs): A sustainable approach. In Sustainable Green Technologies for Environment; Shah, S., Venkatramanan, V., Prasad, R., Eds.; Springer: Singapore, 2019; pp. 111–139. [Google Scholar]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current state of knowledge in microbial degradation of polycyclic aromatic hydrocarbons (PAHs): A review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [PubMed]
- Pozdnyakova, N.; Balandina, S.; Turkovskaya, O. Degradative properties of Pleurotus fungi. In Pleurotus Mushrooms: Ecology, Cultivation and Uses; Mervyn, P., Gwynn, I., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2017; pp. 1–56. [Google Scholar]
- Muratova, A.Y.; Turkovskaya, O.V.; Antonyuk, L.P.; Makarov, O.E.; Pozdnyakova, L.I.; Ignatov, V.V. Oil-oxidizing potential of associative rhizobacteria of the genus Azospirillum. Microbiology 2005, 74, 210–215. [Google Scholar] [CrossRef]
- Gruz-Hernandez, M.A.; Mendoza-Herrera, A.; Bocanegra-Garcia, V.; Rivera, G. Azospirillum spp. From plant growth-promoting bacteria to their use in bioremediation. Microorganisms 2022, 10, 1057. [Google Scholar] [CrossRef]
- Diamantidis, G.; Eosse, A.; Potier, P.; Bally, R. Purifcation and characterization of the first bacterial laccase in the rhizospheric bacterium Azospirillum lipoferum. Soil Biol. Biochem. 2000, 32, 919–927. [Google Scholar] [CrossRef]
- Kupriashina, M.A.; Petrov, S.V.; Ponomareva, E.G.; Nikitina, V.E. Ligninolytic activity of bacteria of the genera Azospirillum and Niveispirillum. Microbiology 2015, 84, 791–795. [Google Scholar] [CrossRef]
- Bezalel, L.; Hadar, Y.; Cerniglia, C. Enzymatic mechanisms involved in phenanthrene degradation by the white rot fungus Pleurotus ostreatus. Appl. Environ. Microbiol. 1997, 63, 2495–2501. [Google Scholar] [CrossRef]
- Nikiforova, S.; Pozdnyakova, N.; Makarov, O.; Chernyshova, M.; Turkovskaya, O. Chrysene bioconversion by the white rot fungus Pleurotus ostreatus D1. Microbiology 2010, 79, 456–460. [Google Scholar] [CrossRef]
- Reasoner, D.J.; Geldreich, E.E. A new medium for the enumeration and subculture of bacteria from potable water. Appl. Environ. Microbiol. 1985, 49, 1–7. [Google Scholar] [CrossRef]
- Tsivileva, O.M.; Shaternikov, A.N.; Nikitina, B.E. Bakterii roda Azospirillum v optimizatsii iskusstvennogo kul’tivirovaniya vysshikh gribov-ksilotrofov (Bacteria of the Azospirillum genus for the optimization of the artificial culture of xylotrophic mushrooms). Biotekhnologiya (Biotechnology) 2020, 36, 16–25, (In Russian, abstract in English). [Google Scholar]
- Niku-Paavola, M.; Karhunen, E.; Salola, P.; Raunio, V. Ligninolytic enzymes of the white rot fungus Phlebia radiata. Biochem. J. 1988, 254, 877–884. [Google Scholar] [CrossRef]
- Martinez, M.; Ruiz-Duenas, F.; Guillen, F.; Martinez, A. Purification and catalytic properties of two manganese peroxidase isoenzymes from Pleurotus eryngii. Eur. J. Biochem. 1996, 237, 424–432. [Google Scholar] [CrossRef]
- Royse, D.J.; Ries, S.M. The influence of fungi isolated from peach twigs on the pathogenicity of Cytospora cincta. Phytopathology 1978, 68, 603–607. [Google Scholar] [CrossRef]
- Kalaiselvi, S.; Panneerselvam, A. In vitro assessment of antagonistic activity of Trichoderma sp. against Sarocladium oryzae causing sheath rot disease in paddy. Int. J. Appl. Biol. Pharm. Technol. 2011, 2, 179–183. [Google Scholar]
- Haritash, A.; Kaushik, C. Biodegradation aspects of polycyclic aromatic hydrocarbons (PAHs). J. Hazard. Mater. 2009, 169, 1–15. [Google Scholar] [CrossRef]
- Lambert, M.; Kremer, S.; Sterner, O.; Anke, H. Metabolism of pyrene by the basidiomycete Crinipellis stipitaria and identification of pyrene quinones and their hydroxylated precursors in strain JK375. Appl. Environ. Microbiol. 1994, 60, 3597–3601. [Google Scholar] [CrossRef]
- Pozdnyakova, N.; Balandina, S.; Turkovskaya, O. Degradativnaya aktivnost’ gribov po otnosheniyu k uglevodorodam nefti v usloviyakh povyshennoy temperatury (Degradative activity of fungi towards oil hydrocarbons under high temperature). Teor. I Prikl. Ekologiya. (Theor. Appl. Ecol.) 2019, 4, 30–36, (In Russian, abstract in English). [Google Scholar]
- Pozdnyakova, N.N.; Dubrovskaya, E.V.; Grinev, V.S.; Turkovskaya, O.V. Perspektivy ispol’zovaniya ksilotrofnykh gribov Pleurotus ostreatus Florida i Schizophyllum commune dlya mikoremediatsii pochv, zagryaznennykh neftyanymi uglevodorodami i poverkhnostno-aktivnymi veshchestvami (Prospects for the use of xylotrophic fungi Pleurotus ostreatus Florida and Schizophyllum commune for mycoremediation of soils contaminated with oil hydrocarbons and surfactants). Biotekhnologiya (Biotechnology) 2021, 37, 108–116, (In Russian, abstract in English). [Google Scholar]
- Adamski, M.; Pietr, S. Biodiversity of bacteria associated with eight Pleurotus ostreatus (Fr.) P. Kumm. strains from Poland, Japan and the USA. Pol. J. Microbiol. 2019, 68, 71–81. [Google Scholar] [CrossRef]
- Bashan, Y.; De-Bashan, L.E. How the plant growth-promoting bacterium Azospirillum promotes plant growth—A critical assessment. Adv. Agron. 2010, 108, 77–136. [Google Scholar]
- Fukami, J.; Cerezini, P.; Hungria, M. Azospirillum: Benefits that go far beyond biological nitrogen fixation. AMB Express 2018, 8, 73. [Google Scholar] [CrossRef]
- Gałązka, A.; Król, M.; Perzyński, A. The efficiency of rhizosphere bioremediation with Azospirillum sp. and Pseudomonas stutzeri in soils freshly contaminated with PAHs and diesel fuel. Pol. J. Environ. Stud. 2012, 21, 345–353. [Google Scholar]
- Kulkarni, K.; Bhogale, G.M.; Nalawade, R. Adsorptive removal of fluoride from water samples using Azospirillum biofertilizer and lignite. Korean J. Chem. Eng. 2017, 35, 153–163. [Google Scholar] [CrossRef]
- Bondarenkova, A.D.; Muratova, A.Y.; Turkovskaya, O.V. Influence of oil on the associative rhizobacterium Azospirillum brasilense. Bull. Saratov State Agrar. Univ. N.I. Vavilov 2009, 9, 5–10, (In Russian, abstract in English). [Google Scholar]
- Obase, K. Bacterial community on ectomycorrhizal roots of Laccaria laccata in a chestnut plantation. Mycoscience 2019, 60, 40–44. [Google Scholar] [CrossRef]
- Loshchinina, E.A.; Nikitina, V.E.; Tsivileva, O.M.; Stepanova, L.V.; Ponomareva, E.G.; Sheludko, A.V. Morphological and cultural characteristics of the basidiomycete Lentinus edodes in co-cultivation with bacteria of the genus Azospirillum. Bull. Saratov State Agrar. Univ. N.I. Vavilov 2006, 6, 24–26, (In Russian, abstract in English). [Google Scholar]
- Valkova, H.; Novotny, C.; Malachova, K.; Slovarcikova, P.; Fojtik, J. Effect of bacteria on the degradation ability of Pleurotus ostreatus. Sci. Total Environ. 2017, 584–585, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski-Dyé, F.; Lozano, L.; Acosta-Cruz, E.; Borland, S.; Drogue, B.; Prigent-Combaret, C.; Rouy, Z.; Barbe, V.; Mendoza Herrera, A.; González, V.; et al. Genome sequence of Azospirillum brasilense CBG497 and comparative analyses of Azospirillum core and accessory genomes provide insight into niche adaptation. Genes 2012, 3, 576–602. [Google Scholar] [CrossRef]
- Chen, J.P.; Lopez-de-Victoria, G.; Lowell, C.R. Utilization of aromatic compounds as carbon and energy sources during growth and N2-fixation by free-living nitroxen fixing bacteria. Arch. Microbiol. 1993, 159, 207–212. [Google Scholar] [CrossRef]
- Krol, M.J.; Perzynski, A. The utilization of polycyclic aromatic hydrocarbons (PAHs) as the sole carbon source in fixation of nitrogen-free by Azospirillum spp. strains of bacteria. Pol. J. Agron. 2002, 131, 69–80. [Google Scholar]
- Cruz-Hernandez, M.A.; Jimenez-Andrade, J.M.; Mendoza-Herrera, A. Characterization of the degradation potential of xenobiotic compounds by the rhizobacteria Azospirillum brasilense. Mex. J. Biotechnol. 2019, 4, 10–22. [Google Scholar] [CrossRef]
- Zeng, J.; Lin, X.; Zhang, J.; Li, X.; Wong, M.H. Oxidation of polycyclic aromatic hydrocarbons by the bacterial laccase CueO from E. coli. Appl. Microbiol. Biotechnol. 2011, 89, 1841–1849. [Google Scholar] [CrossRef]
- Flores, C.; Vidal, C.; Trejo-Hernandez, M.R.; Galino, E.; Serrano-Carreon, L. Selection of Trichoderma strains capable of increasing laccase production by Pleurotus ostreatus and Agaricus bisporus in dual cultures. J. Appl. Microbiol. 2009, 106, 249–257. [Google Scholar] [CrossRef]
- Kang, Y.M.; Cho, K.M. Identification of auxin from Pseudomonas sp. P7014 for the rapid growth of Pleurotus eryngii mycelium. Korean J. Microbiol. 2014, 50, 15–21. [Google Scholar] [CrossRef]
- Febriansyah, E.; Saskiawan, I.; Mangunwardoyo, W.; Sulistiyani, T.R.; Widhiya, E.W. Potency of growth promoting bacteria on mycelial growth of edible mushroom Pleurotus ostreatus and its identification based on 16S rDNA analysis. AIP Conf. Proc. 2018, 2002, 020023. [Google Scholar]
- Young, L.-S.; Chu, J.-N.; Hameed, A.; Young, C.-C. Cultivable mushroom growth-promoting bacteria and their impact on Agaricus blazei productivity. Pesq. Agropecu. Bras. 2013, 48, 636–644. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–430. [Google Scholar] [CrossRef] [PubMed]
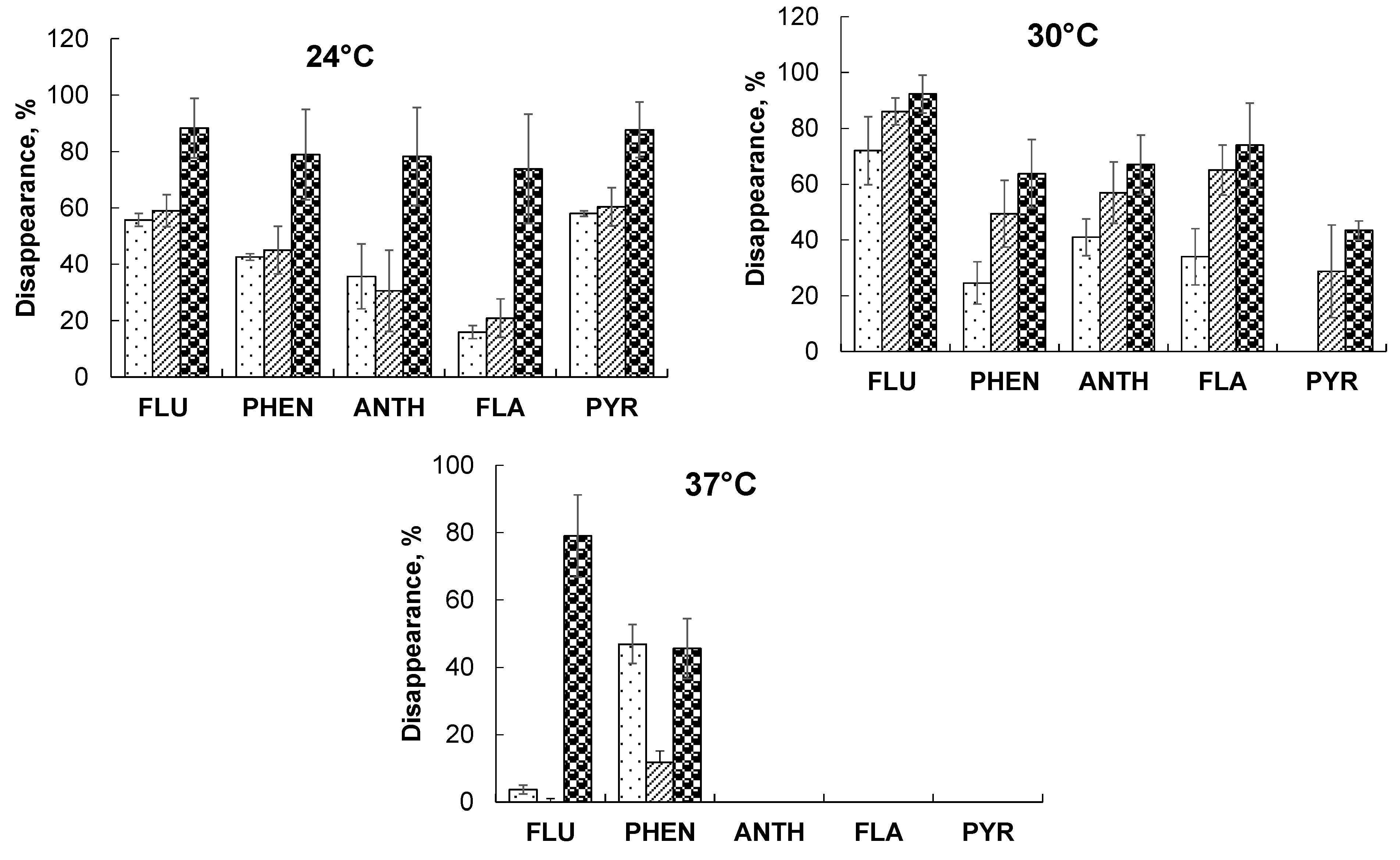
 ), P. ostreatus Florida (
), P. ostreatus Florida ( ), and A. brasilense + P. ostreatus (
), and A. brasilense + P. ostreatus ( ) at different temperatures: FLU—fluorene, PHEN—phenanthrene, ANTH—anthracene, FLA—fluoranthene, PYR—pyrene.
), P. ostreatus Florida (), and A. brasilense + P. ostreatus () at different temperatures: FLU—fluorene, PHEN—phenanthrene, ANTH—anthracene, FLA—fluoranthene, PYR—pyrene.
) at different temperatures: FLU—fluorene, PHEN—phenanthrene, ANTH—anthracene, FLA—fluoranthene, PYR—pyrene.
), P. ostreatus Florida (), and A. brasilense + P. ostreatus () at different temperatures: FLU—fluorene, PHEN—phenanthrene, ANTH—anthracene, FLA—fluoranthene, PYR—pyrene. ), P. ostreatus Florida (), and A. brasilense + P. ostreatus ().
), P. ostreatus Florida (), and A. brasilense + P. ostreatus ().
), P. ostreatus Florida (), and A. brasilense + P. ostreatus ().
), P. ostreatus Florida (), and A. brasilense + P. ostreatus ().
 —control (without IAA);
—control (without IAA);  —with IAA.
—control (without IAA); —with IAA.
—with IAA.
—control (without IAA); —with IAA.
 —7 days; —14 days.
—7 days; —14 days.
—7 days; —14 days.
—7 days; —14 days.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture | Bacterial Count (CFU × 107/mL) | Mycelium Dry Weight (mg) | ||||
|---|---|---|---|---|---|---|
| 24 °C | 30 °C | 37 °C | 24 °C | 30 °C | 37 °C | |
| A. brasilense | 1.4 ± 0.4 | 2.0 ± 0.4 | 14.6 ± 1.9 | - | - | - |
| A. brasilense + PAHs | 1.0 ± 0.4 | 0.2 ± 0.1 | 10.8 ± 0.9 | - | - | - |
| P. ostreatus | - | - | - | 212.4 ± 12.9 | 353.5 ± 18.3 | 9.2 ± 2.7 |
| P. ostreatus + PAHs | - | - | - | 154.5 ± 8.8 | 295.0 ± 30.1 | 22.7 ± 4.3 |
| P. ostreatus + A. brasilense | 0.5 ± 0.0 | 3.2 ± 0.2 | 109.0 ± 8.8 | 270.5 ± 12.1 | 641.4 ± 27.0 | 40.3 ± 7.5 |
| P. ostreatus + A. brasilense + PAHs | 6.1 ± 0.9 | 40.5 ± 7.4 | 83.0 ± 2.8 | 153.8 ± 15.0 | 73.4 ± 10.1 | 13.4 ± 2.7 |
| Culture | Laccase Activity (U/mL) | Versatile Peroxidase Activity (U/mL) | ||||
|---|---|---|---|---|---|---|
| 24 °C | 30 °C | 37 °C | 24 °C | 30 °C | 37 °C | |
| P. ostreatus | 2.4 ± 0.6 | 0.7 ± 0.1 | 0.2 ± 0.03 | 1.8 ± 0.4 | 0.1 ± 0.02 | 0.1 ± 0.01 |
| P. ostreatus + PAHs | 3.3 ± 0.8 | 0.2 ± 0.05 | 0.06 ± 0.01 | 4.1 ± 0.5 | 3.7 ± 0.8 | 0.6 ± 0.02 |
| P. ostreatus + A. brasilense | 3.1 ± 0.7 | 1.8 ± 0.3 | 0.12 ± 0.03 | 3.0 ± 0.8 | 0.1 ± 0.01 | 0.06 ± 0.01 |
| P. ostreatus + A. brasilense + PAHs | 8.9 ± 1.0 | 2.2 ± 0.9 | 0.36 ± 0.08 | 7.1 ± 1.2 | 2.5 ± 0.5 | 0.3 ± 0.04 |
| Culture | Growth | Enzymatic Activity (U/mL) | ||||||
|---|---|---|---|---|---|---|---|---|
| Bacterial Count (CFU × 107/mL) | Mycelium Dry Weight (mg) | Laccase | Versatile Peroxidase | |||||
| FLU 1 | FLUQ 2 | FLU | FLUQ | FLU | FLUQ | FLU | FLUQ | |
| A. brasilense | 0.06 ± 0.01 | 0.02 ± 0.004 | - | - | 0 | 0 | 0 | 0 |
| P. ostreatus | - | - | 309.9 ± 12.1 | 303.8 ± 17.0 | 1.90 ± 0.4 | 1.12 ± 0.3 | 0.83 ± 0.01 | 0.67 ± 0.07 |
| P. ostreatus + A. brasilense | 1.8 ± 0.03 | 1.3 ± 0.04 | 318.0 ± 26.0 | 350.6 ± 15.8 | 2.61 ± 0.7 | 1.35 ± 0.3 | 1.13 ± 0.3 | 0.92 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pozdnyakova, N.; Muratova, A.; Turkovskaya, O. Degradation of Polycyclic Aromatic Hydrocarbons by Co-Culture of Pleurotus ostreatus Florida and Azospirillum brasilense. Appl. Microbiol. 2022, 2, 735-748. https://doi.org/10.3390/applmicrobiol2040056
Pozdnyakova N, Muratova A, Turkovskaya O. Degradation of Polycyclic Aromatic Hydrocarbons by Co-Culture of Pleurotus ostreatus Florida and Azospirillum brasilense. Applied Microbiology. 2022; 2(4):735-748. https://doi.org/10.3390/applmicrobiol2040056
Chicago/Turabian StylePozdnyakova, Natalia, Anna Muratova, and Olga Turkovskaya. 2022. "Degradation of Polycyclic Aromatic Hydrocarbons by Co-Culture of Pleurotus ostreatus Florida and Azospirillum brasilense" Applied Microbiology 2, no. 4: 735-748. https://doi.org/10.3390/applmicrobiol2040056
APA StylePozdnyakova, N., Muratova, A., & Turkovskaya, O. (2022). Degradation of Polycyclic Aromatic Hydrocarbons by Co-Culture of Pleurotus ostreatus Florida and Azospirillum brasilense. Applied Microbiology, 2(4), 735-748. https://doi.org/10.3390/applmicrobiol2040056