1. Introduction
The winged bean (
Psophocarpus tetragonolobus (L.) DC.) is a neglected and underutilized legume that thrives well in hot, humid, equatorial countries [
1]. It belongs to the family
Fabaceae. It is predominantly grown in Papua New Guinea, Malaysia, Thailand, Bangladesh, Indonesia, Ghana, and to a lesser extent, Nigeria [
2]. It is nutritious, with an exceptionally high number of proteins, vitamins, and fatty oils that could play a significant role in the diet improvement of people in sub-Saharan Africa (SSA) countries [
3]. It is a multipurpose legume grown for its edible seeds, vegetable, and root tuber. The dried tubers and mature seeds contain about 17–19% and 28–45% proteins, respectively, and a high oil content of about 14–20.4%, similar to soybean [
4,
5,
6,
7]. Winged bean flour has the potential to reduce dietary deficiency in children and adults of SSA when blended and prepared into different meals [
4,
8]. Due to the increasing global food demand, the winged bean is a prospective crop that, when cultivated and utilized, could enhance nutrition and food security, especially in the developing countries of SSA [
2,
9].
Despite the nutritional potentials of the winged bean, very little attention is paid to exploiting it due to its long maturation period, indeterminate growth habit, low seed yield, and the need for a stake to support the vigorously growing vines, as well as the presence of anti-nutritive factors, including chymotrypsin inhibitors, hemagglutinins, and trypsin inhibitors [
5]. In addition, there is limited information about the genetic diversity among the available winged bean germplasm [
10]. Understanding the genetic diversity of existing landraces from various geographical regions is critical for their conservation and, subsequently, their utilization in breeding efforts [
2]. Information on genetic diversity is needed by plant breeders to develop effective approaches, as well as provide information on heterotic combinations in crop improvement programs [
11]. Researchers have made several efforts to assess the genetic diversity of winged beans using agro-morphological traits [
12,
13,
14,
15]. For example, Yulianah et al. [
12] assessed the genetic diversity of 21 Indonesian winged bean accessions based on morphological characters. They reported that phenotypic traits, including pod length, number of pods per plant, and total pod weight per plant, are useful in discriminating among the winged bean accessions. Similarly, Wong et al. [
15] evaluated 24-winged bean accessions sourced from Nigeria, Malaysia, Bangladesh, Papua New Guinea, Indonesia, Sri Lanka, and Liberia based on their yield-related traits. Apart from agro-morphological traits, molecular tools have also been employed to assess the genetic diversity of winged bean accessions using molecular markers. Mohanty et al. [
7] used Inter Simple Sequence Repeat (ISSR) and Random Amplified Polymorphic DNA (RAPD) markers to assess the genetic diversity of 24-winged bean accessions. In another study, Laosatit et al. [
10] employed 14 simple sequence repeats (SSR) markers to assess the genetic diversity and the population structure in 457 accessions of winged beans collected from six geographical regions in Thailand. However, it is important to assess the genetic diversity of newly collected winged accessions and already existing accessions in the IITA-Gene bank.
The multi–Spectral Imaging (MSI) system is an integrated computer-aided image analysis system for assessing morphometry and distinguishing seed variation [
16]. It measures a variety of seed morphometric features and provides a more accurate assessment of the seed features such as size, color, shape, and texture to discriminate among different accessions quantitatively [
17]. Additionally, it replaces subjective and perception-based evaluation with a more accurate objective quantification in the analysis of seed morphometric variation [
16,
17]. Venora et al. [
18] used an image analysis system for varietal identification of 15 Italian common bean landraces based on the seed size and color descriptors. Similarly, Kilic et al. [
19] developed a computer vision system and artificial neural network based on seed size and color for the classification of 69 common bean samples. In addition, Geetha et al. [
20] used a seed image analyzer to characterize the shape descriptors of nine mustard genotypes. They found most seed geometric features, including area, perimeter, circularity, and length, useful in discriminating the genotypes. However, the application and effectiveness of the MSI system in assessing the genetic diversity among winged bean accessions are yet to be reported. The MSI system offers to be an invaluable tool in the phenotypic characterization of winged bean accessions in the IITA-Gene bank. Hence, this study aimed to assess the genetic diversity of thirty-winged bean accessions using agro-morphological traits and seed morphometric traits derived from a MSI system.
4. Discussion
The significant variations observed among the accessions for seed yield per plant and all other agro-morphological traits revealed that adequate variation existed among the accessions. This implied that the accessions expressed variation for each trait, which could enable effective selection among the accessions. Additionally, the observed variation among the accessions could be used in possible combinations in crop improvement programs. Yulianah et al. and Wong et al. [
12,
15] reported similar findings in their study on winged bean accessions characterized based on their phenotypic traits. The observed significant mean squares for the environment for all traits suggest the distinctiveness of the environments in discriminating among the accessions. It, however, still suggests the need for more evaluation in different environments to ascertain the genotypic integrity of the accessions. The non-significant differences observed for the interaction between the accessions and the environments implied that the performance of the winged bean accessions was consistent across the study environments.
In this study, higher genotypic correlation coefficients were observed than phenotypic correlation. This denotes that genes largely influence trait expressions. The significant and positive phenotypic and genotypic correlation observed between seed yield per plant and hundred seed weight, pod weight per plant, number of locules per pod, and the number of seeds per pod implied that these traits could be substituted for seed yield during selection in crop improvement. In addition, the correlation recorded between these desirable traits implied that the improvement of one trait simultaneously improves the other trait [
29,
30]. These results agree with the findings of previous authors on the association between seed yield per plant and hundred seed weight in winged beans [
29] and AYB [
31]. In addition, seed yield per plant and pod weight per plant in AYB [
31]; seed yield per plant and number of seeds per pod in common bean [
32]; days to flowering and days to maturity in winged bean [
29]. The positive phenotypic and genotypic significant correlations among the yield-related traits suggested that these traits could be considered for selection to improve seed yield in the winged bean. The non-significant correlation observed between the number of pods per plant and seed yield per plant indicates that the seed yield is independent of the number of pods produced by the plant.
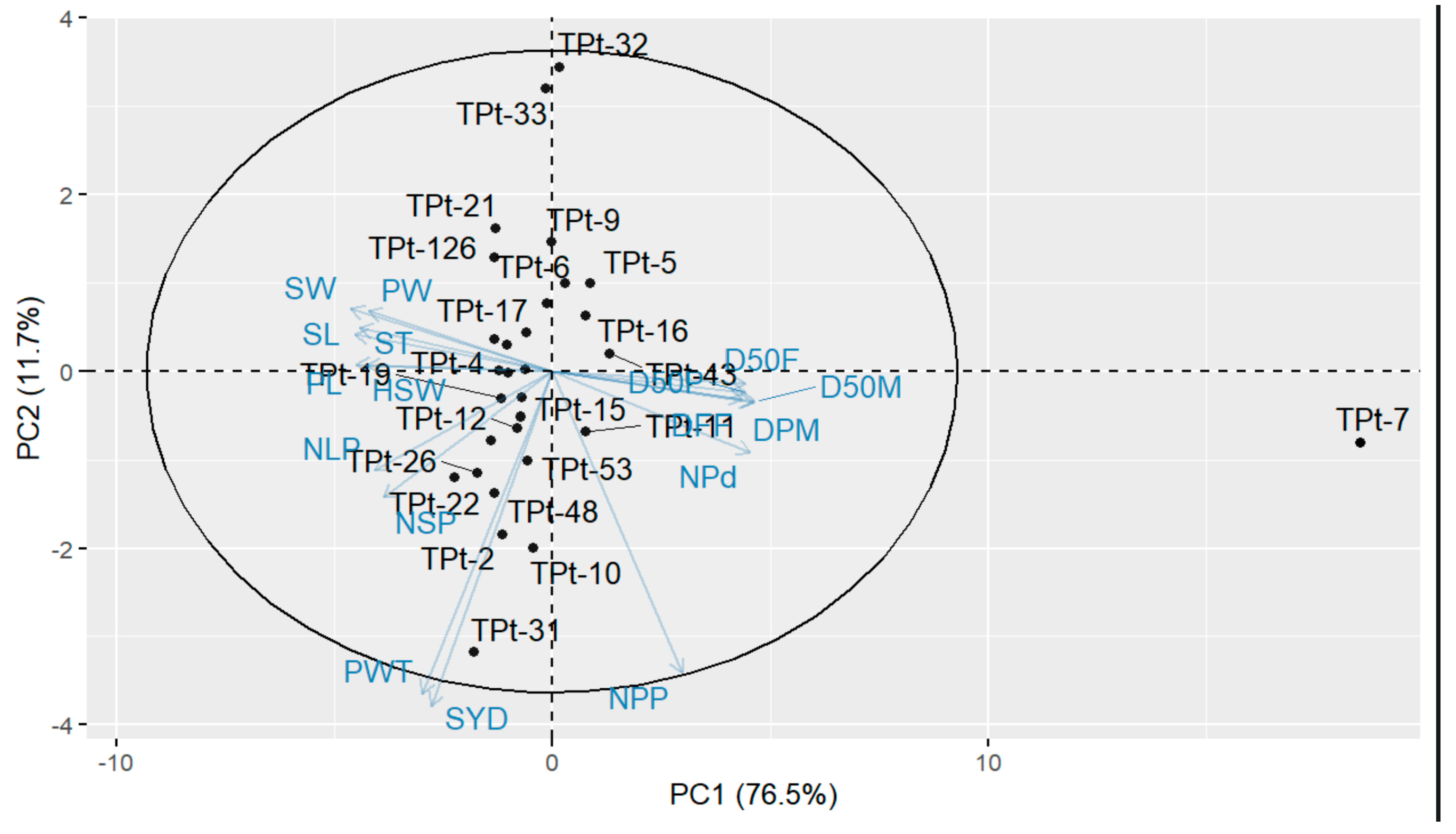
All the 17 agro-morphological traits measured were found to be relevant in discriminating the winged bean accessions based on the contribution of each of the measured traits to the most informative principal components. This implied that the traits with high coefficients in the three components could be used to select or characterize other winged bean collections [
33]. Furthermore, these traits could be considered for genetic improvement in winged beans [
31].
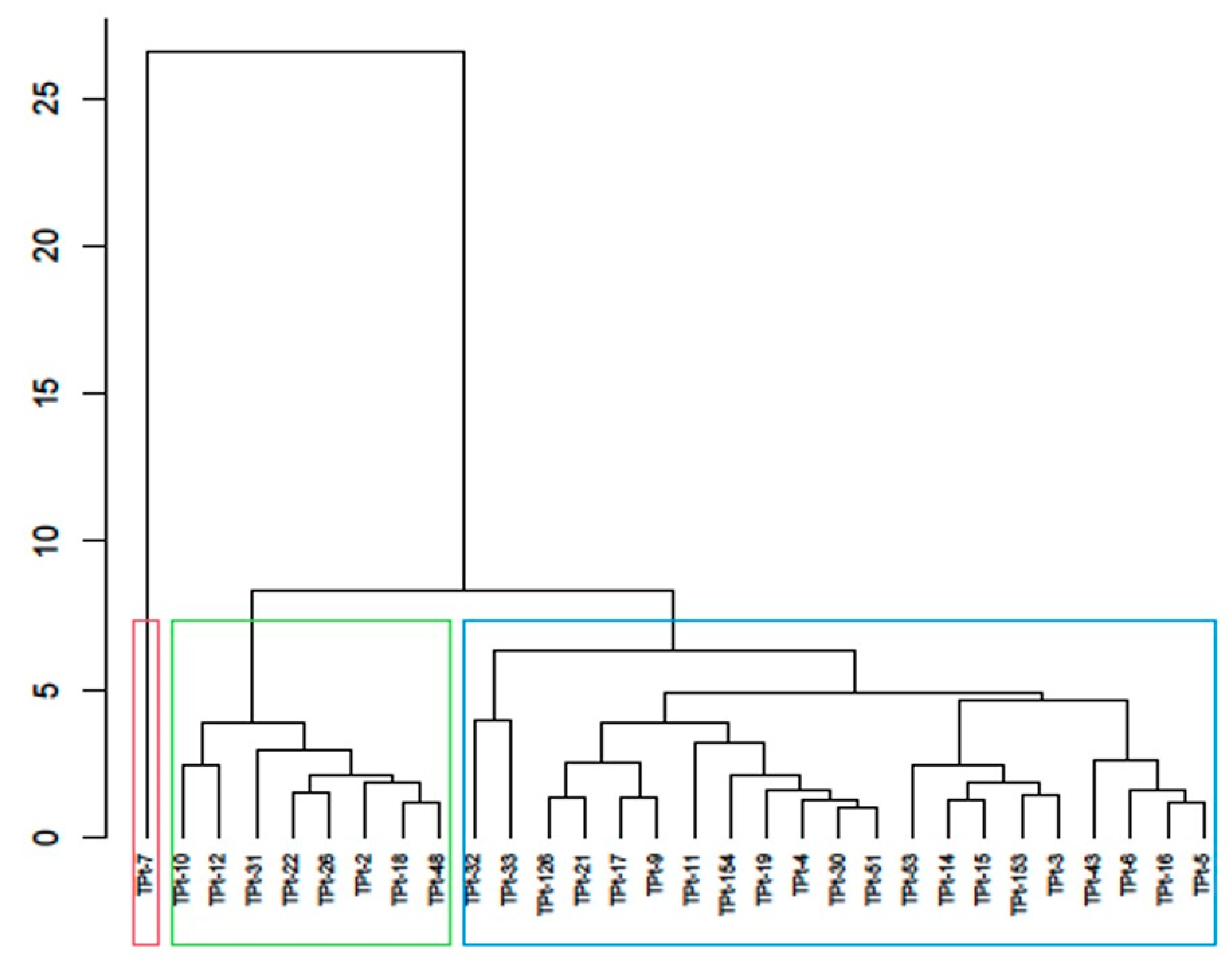
To assess the existing diversity among the winged bean accessions, the dendrogram generated from cluster analysis classified the 30 accessions into three distinct groups. The classification was based on their similarity for one or more agro-morphological traits studied, irrespective of their geographical origin. TPt-7 was placed in cluster I and characterized by the longest duration to flowering, longest days to maturity, the highest number of pods per peduncle, and the highest number of pods per plant. This implied that TPt-7 possesses unique characteristics that could be harnessed in crop improvement programs. Accessions in cluster II possess earliness to flowering and maturity, a high number of pods per plant, high pod weight, and high seed yield, while accessions in cluster III have early to mid-number of days to flowering with a high number of pods. This result suggests that accessions were grouped based on their distinct features and could be a reliable source for hybridization when improving for reduced days to flowering and high seed yield in the winged bean. This further suggests that heterosis could be exploited by selecting and hybridizing distinct accessions belonging to different groups [
31,
33].
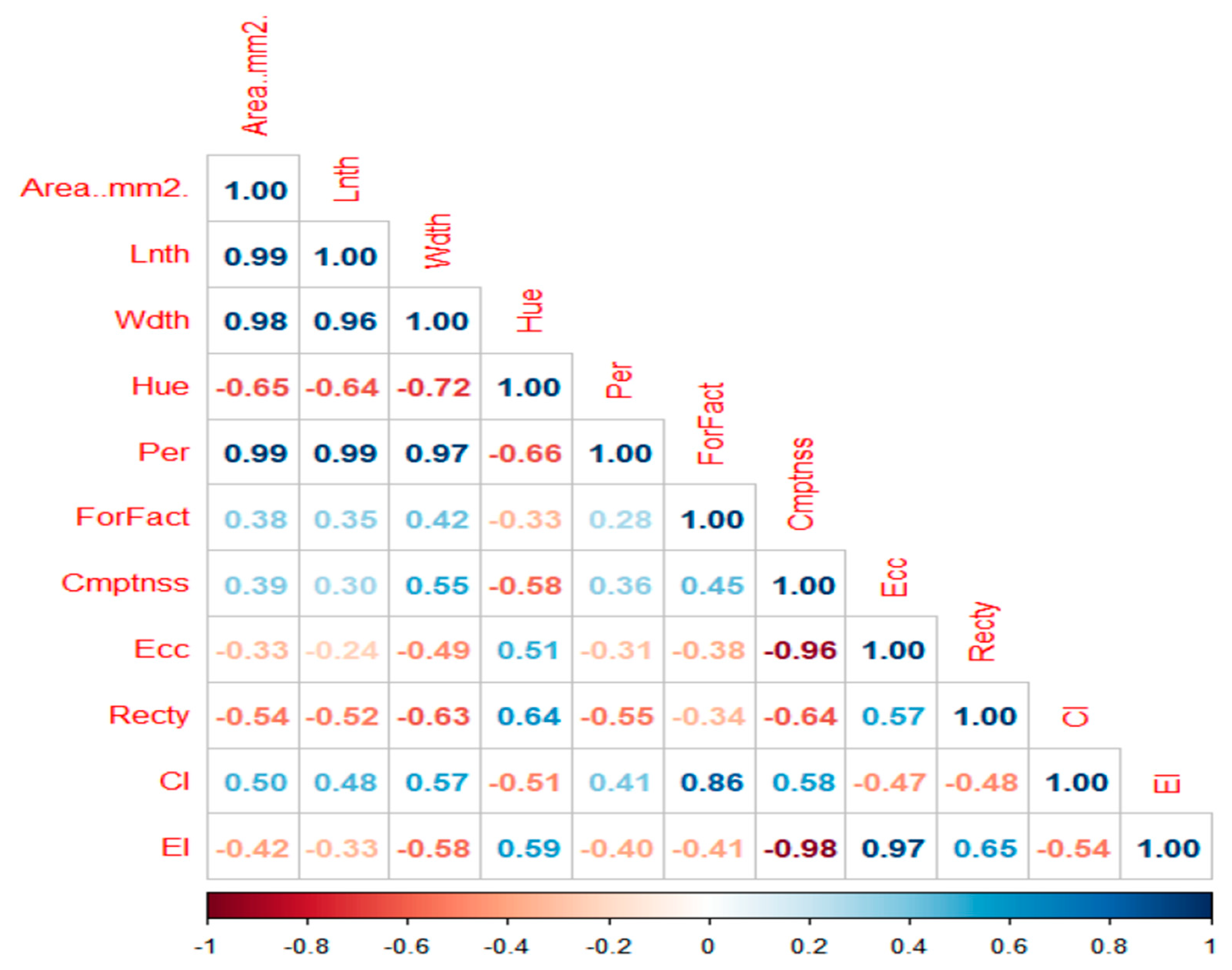
A positive correlation was observed between seed length and seed area; seed width and seed area; seed perimeter and seed area, implying that an increase in one trait simultaneously improves the other. On the other hand, a negative association was recorded between seed hue and seed area, seed eccentricity and seed area, seed rectangularity and seed area, and seed eccentricity index and seed area, which denotes that if one trait increases, the other trait decreases. This result agrees with Daniel et al. and Adewale et al. [
27,
28], who reported similar findings for AYB and maize seeds, respectively. Adewale et al. [
28] recommended using six seed traits, including seed length and width, as unique parameters of AYB based on the wide variability between them and using the four seed shape indices, including eccentricity index, as a complement for seed shape description in AYB.
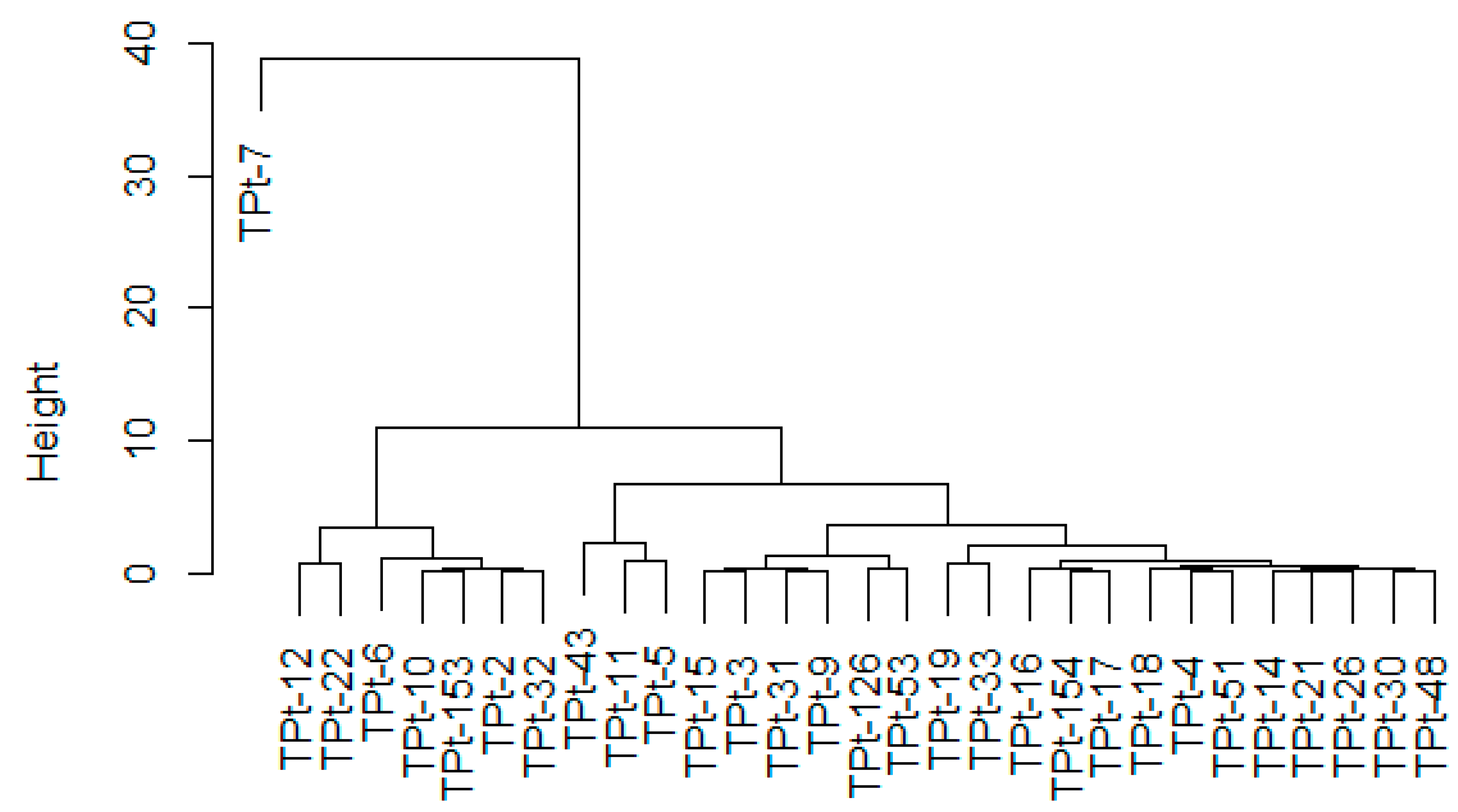
The dendrogram generated from the seed morphometry showed that the accessions clustered based on similarities between one or more seed morphometric traits. This implied that the classification formed a basis for grouping parental lines based on seed morphometric measurements. This constitutes background data for future evaluation of crosses involving these parents. The grouping pattern in the agro-morphological and seed morphometric clusters showed the solitary clustering of TPt-7due to its distinct features. This implied that TPt-7 might belong to the species
scandens, as reported by Yang et al. [
34]. They reported a successful hybridization between
Psophocarpus tetragonolobus and
Psophocarpus scandens. Accessions (TPt-10, TPt-12, TPt-22, TPt-2); (TPt-33, TPt-126, TPt-21, TPt-17, TPt-9, TPt-11, TPt-154, TPt-19, TPt-4, TPt-30, TPt-51, TPt-53, TPt-14, TPt-15, TPt-3, TPt-43, TPt-16, and TPt-5) were grouped in the same clusters in the two dendrograms generated from this study. This implied that the accessions were closely related, and the differences observed could be a result of environmental influence.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}