
The Decline of Common Birds Exemplified by the Western Jackdaw Warns on Strong Environmental Degradation

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
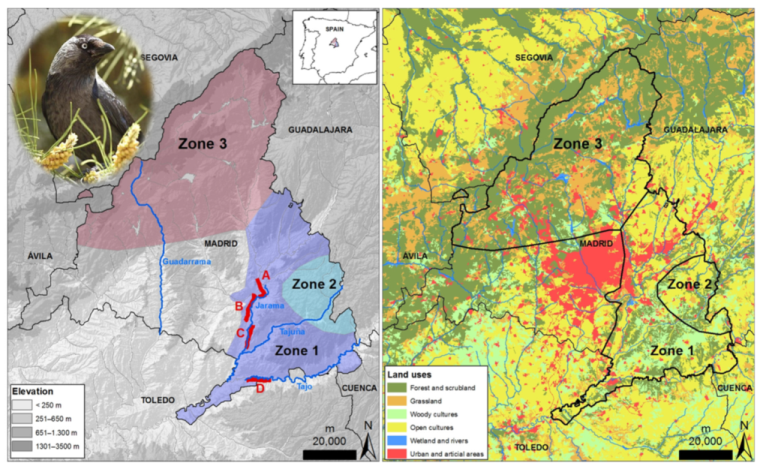
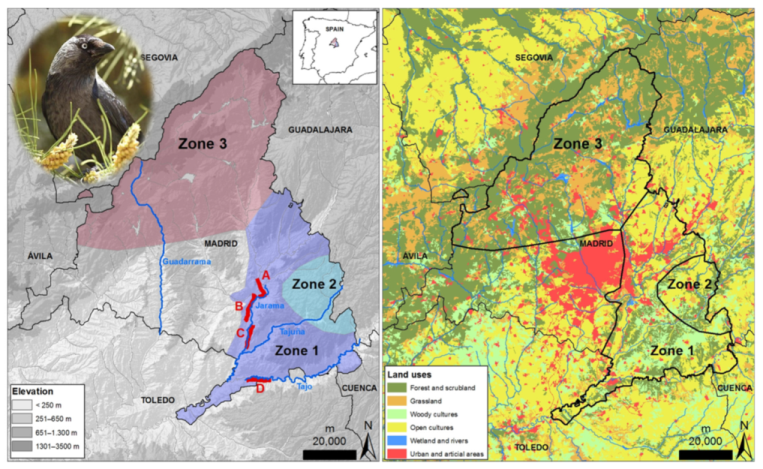
2.1. Study Species and Study Area
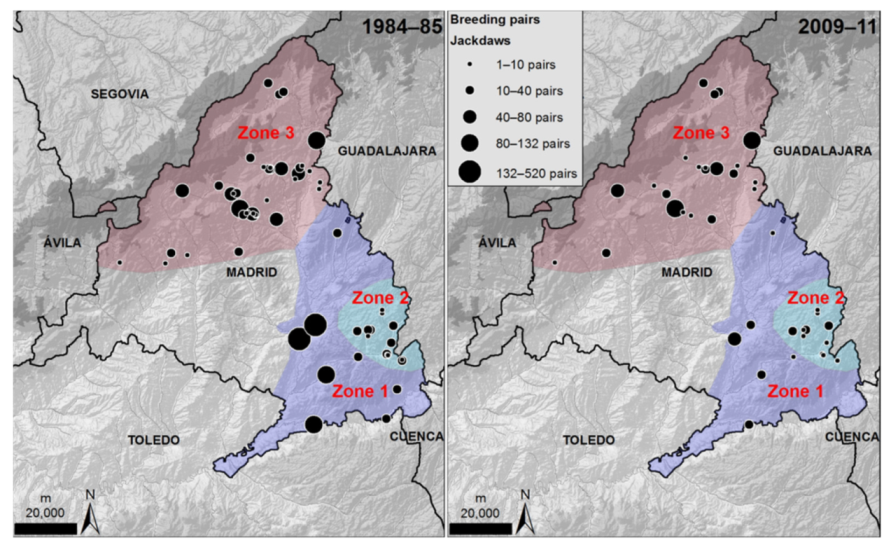
2.2. Breeding Abundance in Gypsum Cliffs
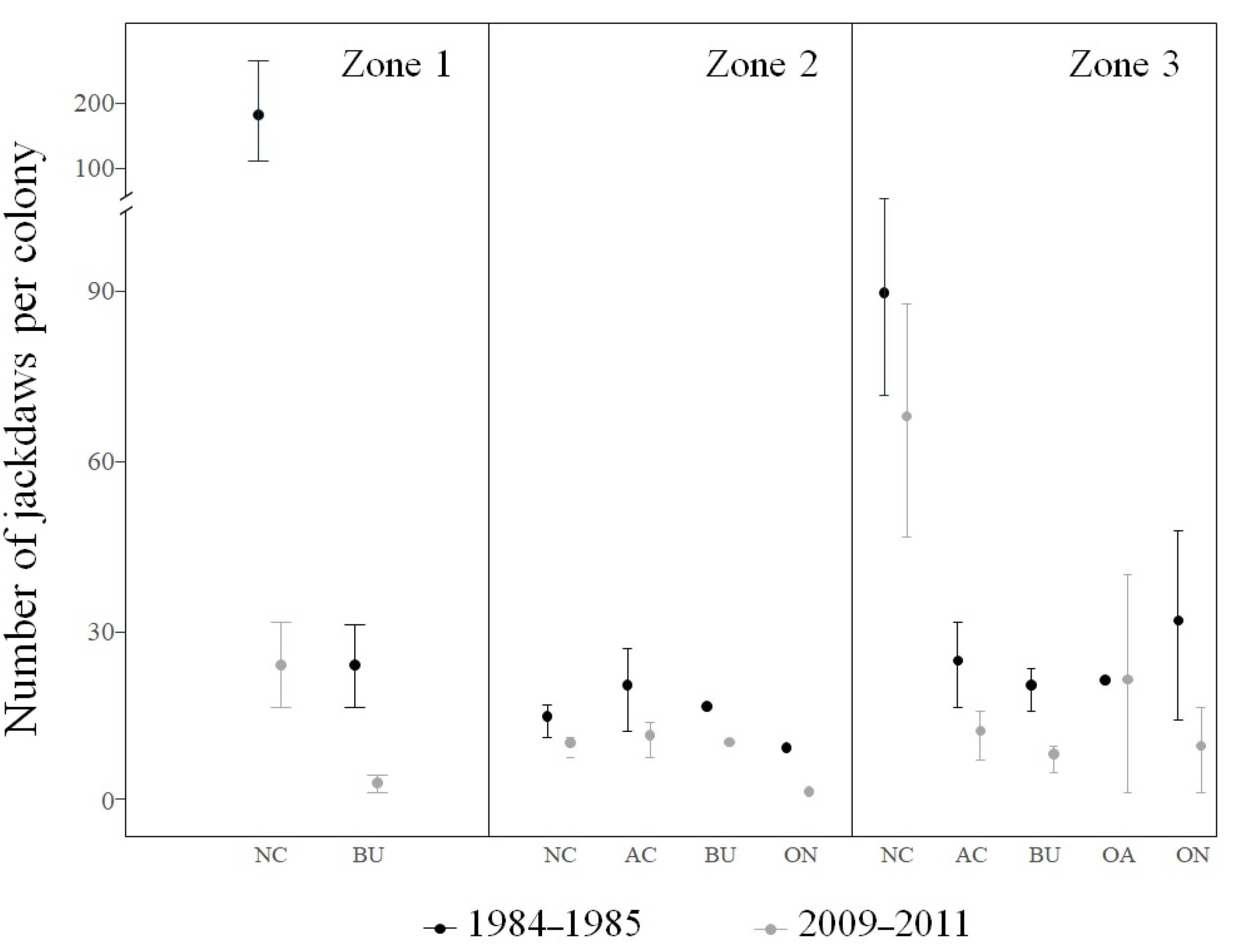
2.3. Colony Size
2.4. Breeding Abundance in Riverine Forest
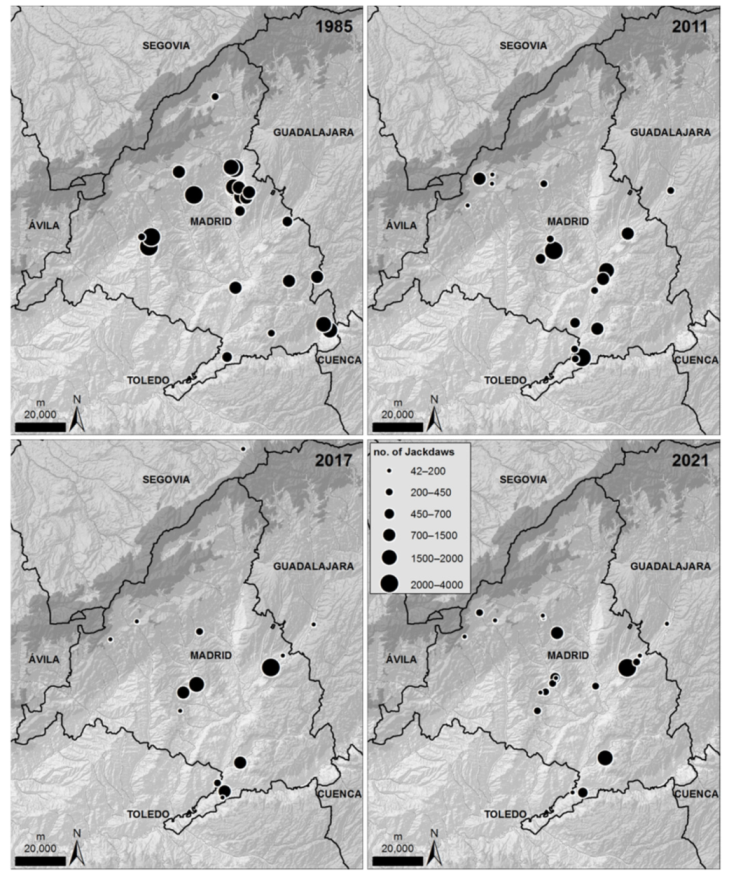
2.5. Winter Communal Roosts
2.6. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- BirdLife International. State of the World’s Birds: Taking the Pulse of the Planet; BirdLife International: Cambridge, UK, 2018. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species; Version 2021-1; IUCN: London, UK, 2021. [Google Scholar]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.; Almond, R.E.; Watson, R. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- Gamero, A.; Brotons, L.; Brunner, A.; Foppen, R.; Fornasari, L.; Gregory, R.D.; Herrando, S.; Hořák, D.; Jiguet, F.; Kmecl, P.; et al. Tracking progress toward EU biodiversity strategy targets: EU policy effects in preserving its common farmland birds. Conserv. Lett. 2017, 10, 395–402. [Google Scholar] [CrossRef]
- Sekercioglu, C.H. Increasing awareness of avian ecological function. Trends Ecol. Evol. 2006, 21, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.J. Valuing common species. Science 2010, 327, 154–155. [Google Scholar] [CrossRef]
- Green, A.J.; Elmberg, J.; Lovas-Kiss, Á. Beyond scatter-hoarding and frugivory: European corvids as overlooked vectors for a broad range of plants. Front. Ecol. Evol. 2019, 7, 133. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J.; Fuller, R.A. Commonness, population depletion and conservation biology. Trends Ecol. Evol. 2007, 23, 14–19. [Google Scholar] [CrossRef]
- Vorisek, P.; Jiguet, F.; van Strien, A.; Škorpilová, J.; Klvanová, A.; Gregory, R.D. Trends in Abundance and Biomass of Widespread European Farmland Birds: How Much Have We Lost? BOU Proceedings–Lowland Farmland Birds III. 2010. Available online: https://www.bou.org.uk/bouproc-net/lfb3/vorisek-etal.pdf (accessed on 11 December 2021).
- Pejchar, L.; Clough, Y.; Ekroos, J.; Nicholas, K.A.; Olsson, O.; Ram, D.; Tschumi, M.; Smith, H.G. Net effects of birds in agroecosystems. BioScience 2018, 68, 896–904. [Google Scholar] [CrossRef]
- Donald, P.F.; Sanderson, F.J.; Burfield, I.J.; van Bommel, F.P.J. Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agric. Ecosyst. Environ. 2006, 116, 189–196. [Google Scholar] [CrossRef]
- Butler, S.J.; Boccaccio, L.; Gregory, R.D.; Vorisek, P.; Norris, K. Quantifying the impact of land-use change to European farmland bird populations. Agric. Ecosyst. Environ. 2010, 137, 348–357. [Google Scholar] [CrossRef]
- Guerrero, I.; Morales, M.B.; Oñate, J.J.; Geiger, F.; Berendse, F.; de Snoo, G.; Sönke, E.; Tomas, P.; Jan, B.; Lars, W.C.; et al. Response of ground-nesting farmland birds to agricultural intensification across Europe: Landscape and field level management factors. Biol. Conserv. 2012, 152, 74–80. [Google Scholar] [CrossRef]
- Emmerson, M.; Morales, M.B.; Oñate, J.J.; Batary, P.; Berendse, F.; Liira, J.; Aavik, T.; Guerrero, I.; Bommarco, R.; Eggers, S.; et al. How agricultural intensification affects biodiversity and ecosystem services. Adv. Ecol. Res. 2016, 55, 43–97. [Google Scholar]
- Green, R.E.; Cornell, S.J.; Scharlemann, J.P.W.; Balmford, A. Farming and the fate of wild nature. Science 2005, 307, 550–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaston, K.J.; Fuller, R.A. Biodiversity and extinction: Losing the common and the widespread. Prog. Phys. Geogr. 2007, 31, 213–225. [Google Scholar] [CrossRef]
- Pe’er, G.; Dicks, L.V.; Visconti, P.; Arlettaz, R.; Báldi, A.; Benton, T.G.; Scott, A.V. EU agricultural reform fails on biodiversity. Science 2014, 344, 1090–1092. [Google Scholar] [CrossRef] [PubMed]
- Reif, J.; Vermouzek, Z. Collapse of farmland bird populations in an Eastern European country following its EU accession. Conserv. Lett. 2019, 12, e12585. [Google Scholar] [CrossRef]
- Nichols, J.D.; Williams, B.K. Monitoring for conservation. Trends Ecol. Evol. 2006, 21, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Stanton, J.C.; Blancher, P.; Rosenberg, K.V.; Panjabi, A.O.; Thogmartin, W.E. Estimating uncertainty of North American landbird population sizes. Avian Conserv. Ecol. 2019, 14, 4. [Google Scholar] [CrossRef]
- Marsh, D.M.; Trenham, P.C. Current trends in plant and animal population monitoring. Conserv. Biol. 2008, 22, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Scholefield, P.; Firbank, L.; Butler, S.; Norris, K.; Jones, L.M.; Petit, S. Modelling the European farmland bird indicator in response to forecast land-use change in Europe. Ecol. Indic. 2011, 11, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.D.; Williams, B.K. The potential for citizen science to produce reliable and useful information in ecology. Conserv. Biol. 2019, 33, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Pollock, K.H.; Nichols, J.D.; Simonst, T.R.; Farnsworth, G.L.; Bailey, L.; Sauer, J.R. Large scale wildlife monitoring studies: Statistical methods for design and analysis. Environmetrics 2002, 13, 105–119. [Google Scholar] [CrossRef]
- Elphick, C.S. How you count counts: The importance of methods research in applied ecology. J. Appl. Ecol. 2008, 45, 1313–1320. [Google Scholar] [CrossRef]
- Snäll, T.; Kindvall, O.; Nilsson, J.; Pärt, T. Evaluating citizen-based presence data for bird monitoring. Biol. Conserv. 2011, 144, 804–810. [Google Scholar] [CrossRef]
- Blanco, G.; Sergio, F.; Sánchez-Zapata, J.A.; Pérez-García, J.M.; Botella, F.; Martínez, F.; Zuberogoitia, I.; Frías, Ó.; Roviralta, F.; Martínez, J.E. Safety in numbers? Supplanting data quality with fanciful models in wildlife monitoring and conservation. Biodiver. Conserv. 2012, 21, 3269–3276. [Google Scholar] [CrossRef] [Green Version]
- Overmars, K.P.; Schulp, C.J.; Alkemade, R.; Verburg, P.H.; Temme, A.J.; Omtzigt, N.; Schaminée, J.H. Developing a methodology for a species-based and spatially explicit indicator for biodiversity on agricultural land in the EU. Ecol. Indic. 2014, 37, 186–198. [Google Scholar] [CrossRef]
- Lewandowski, E.; Specht, H. Influence of volunteer and project characteristics on data quality of biological surveys. Conserv. Biol. 2015, 29, 713–723. [Google Scholar] [CrossRef]
- Kamp, J.; Oppel, S.; Heldbjerg, H.; Nyegaard, T.; Donald, P.F. 2016. Unstructured citizen science data fail to detect long-term population declines of common birds in Denmark. Divers. Distrib. 2016, 22, 1024–1035. [Google Scholar] [CrossRef]
- Sutherland, W.J. (Ed.) Ecological Census Techniques: A Handbook; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Hardey, J.; Crick, H.; Wernham, C.; Riley, H.; Etheridge, B.; Thompson, D. Raptors: A Field Guide to Surveys and Monitoring; The Stationery Office: Edinburgh, UK, 2009. [Google Scholar]
- Norman, D.; Harris, R.J.; Newson, S.E. Producing regional estimates of population size for common and widespread breeding birds from national monitoring data. Bird Study 2012, 59, 10–21. [Google Scholar] [CrossRef]
- Reynolds, J.H.; Thompson, W.L.; Russell, B. Planning for success: Identifying effective and efficient survey designs for monitoring. Biol. Conserv. 2011, 144, 1278–1284. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burgess, N.D.; Hill, D.A.; Mustoe, S.H. Bird Census Techniques, 2nd ed.; Academic Press: London, UK, 2000. [Google Scholar]
- Thompson, W.L. Sampling Rare and Elusive Species; Island Press: Washington, DC, USA, 2004. [Google Scholar]
- Tella, J.L.; Romero-Vidal, P.; Dénes, F.V.; Hiraldo, F.; Toledo, B.; Rossetto, F. Roadside car surveys: Methodological constraints and solutions for estimating parrot abundances across the world. Diversity 2021, 13, 300. [Google Scholar] [CrossRef]
- Hertzog, L.R.; Frank, C.; Klimek, S.; Röder, N.; Böhner, H.G.; Kamp, J. Model-based integration of citizen science data from disparate sources increases the precision of bird population trends. Divers. Distrib. 2021, 27, 1106–1119. [Google Scholar] [CrossRef]
- Marzluff, J.M.; Angell, T. In the Company of Crows and Ravens; Yale University Press: New Haven, CT, USA, 2005. [Google Scholar]
- Madden, C.F.; Arroyo, B.; Amar, A. A review of the impacts of corvids on bird productivity and abundance. IBIS 2015, 157, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Benmazouz, I.; Jokimäki, J.; Lengyel, S.; Juhász, L.; Kaisanlahti-Jokimäki, M.L.; Kardos, G.; Paládi, P.; Kövér, L. Corvids in urban environments: A systematic global literature review. Animals 2021, 11, 3226. [Google Scholar] [CrossRef] [PubMed]
- Marina, G.; Bezares, E. Información sobre los cuervos en España. Inst. For. Investig. Y Exp. 1933, 12, 1–47. [Google Scholar]
- Brochet, A.L.; Van Den Bossche, W.; Jones, V.R.; Arnardottir, H.; Damoc, D.; Demko, M.; Driessens, G.; Flensted, K.; Gerber, M.; Ghasabyan, M.; et al. Illegal killing and taking of birds in Europe outside the Mediterranean: Assessing the scope and scale of a complex issue. Bird Conserv. Int. 2019, 29, 10–40. [Google Scholar] [CrossRef] [Green Version]
- Blanco, G.; Cuevas, J.A.; Frías, Ó.; del Barrio, J.L.G. A shot in the dark: Sport hunting of declining corvids promotes the inadvertent shooting of threatened red-billed choughs. J. Nat. Conserv. 2019, 52, 125739. [Google Scholar] [CrossRef]
- BirdLife International. European Birds of Conservation Concern: Populations, Trends and National Responsibilities; BirdLife International: Cambridge, UK, 2017. [Google Scholar]
- Božič, L. Numbers, distribution and nest site characteristics of Jackdaw Corvus monedula in Slovenia and its conservation status. Acrocephalus 2016, 37, 123–150. [Google Scholar] [CrossRef] [Green Version]
- López-Jiménez, N. (Ed.) Libro Rojo de las Aves de España; SEO/BirdLife: Madrid, Spain, 2021. [Google Scholar]
- Vandewalle, M.; De Bello, F.; Berg, M.P.; Bolger, T.; Doledec, S.; Dubs, F.; Feld, C.K.; Harrington, R.; Harrison, P.A.; Lavorel, S.; et al. Functional traits as indicators of biodiversity response to land use changes across ecosystems and organisms. Biodivers. Conserv. 2010, 19, 2921–2947. [Google Scholar] [CrossRef] [Green Version]
- Mineau, P.; Whiteside, M. Pesticide acute toxicity is a better correlate of U.S. grassland bird declines than agricultural intensification. PLoS ONE 2013, 8, e57457. [Google Scholar] [CrossRef] [Green Version]
- García-Fernández, A.J. Ecotoxicology, avian. Encycl. Toxicol. 2014, 2, 289–294. [Google Scholar]
- Goulson, D. Pesticides linked to bird declines. Nature 2014, 511, 295–296. [Google Scholar] [CrossRef]
- Soler, M. Grajilla—Corvus monedula. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Morales, M.B., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2012; Available online: http://www.vertebradosibericos.org/ (accessed on 13 November 2021).
- BirdLife International. Corvus monedula (Eurasian Jackdaw). In European Red List of Birds; Office for Official Publications of the European Communities: Luxembourg, 2015. [Google Scholar]
- Blanco, G.; Frías, Ó.; Cuevas, J.A.; González, J.L.; Martínez, F. Commonness of not-so-common birds: The need for baseline knowledge of actual population size for the validation of population size predictions. Bird Study 2014, 61, 351–360. [Google Scholar] [CrossRef]
- Frías, Ó.; González del Barrio, J.L.; Martínez, F.; Blanco, G. Grajilla Occidental, Corvus monedula. In SEO/BirdLife: III Atlas de las Aves en Época de Reproducción en ESPAÑA; SEO/BirdLife: Madrid, Spain, 2021; in press. [Google Scholar]
- Frías, Ó.; Bautista, L.M.; Dénes, F.V.; Cuevas, J.A.; Martínez, F.; Blanco, G. Influence of habitat suitability and sex-related detectability on density and population size estimates of habitat-specialist warblers. PLoS ONE 2018, 13, e0201482. [Google Scholar]
- Cuevas, J.A.; Acha, A.; Blanco, G.; Ruiz, P.; Velasco, T.; Delgado, J.A.; De Miguel, J.A. Biodiversidad en ecosistemas fluviales: Las aves acuáticas en la cuenca media del Tajo. Ser. Doc. 2000, 31, 114. [Google Scholar]
- Molina-Holgado, P.M.; Jendrzyczkowski Rieth, L.; Berrocal Menarguez, A.B.; Allende Álvarez, F. The analysis of urban fluvial landscapes in the centre of Spain, their characterization, values and interventions. Sustainability 2020, 12, 4661. [Google Scholar] [CrossRef]
- Gómez Mendoza, J.; Mata Olmo, R.; Sanz Herráiz, C.; Galiana Martín, L.; Manuel Valdés, C.M.; Molina Holgado, P. Los Paisajes de Madrid: Naturaleza y Medio Rural; Alianza Editorial-Fundación Caja Madrid: Madrid, Spain, 1999; 301p. [Google Scholar]
- Blanco, G.; Cuevas, J.A.; Fargallo, J.A. La población de Chova Piquirroja Pyrrhocorax pyrrhocorax en el Sureste de Madrid (Centro de España). Ardeola 1991, 38, 91–99. [Google Scholar]
- Domínguez, L. Ecología de la Grajilla (Corvus monedula Linnaeus 1758) en la Provincial de Madrid. Ph.D Thesis, Universidad Complutense de Madrid, Madrid, Spain, 1999. [Google Scholar]
- Arroyo, B. La avifauna de un cantil estepárico. Ardeola 1976, 23, 41–49. [Google Scholar]
- Velasco, T.; Blanco, G. Avifauna nidificante en los sotos fluviales de la Comunidad de Madrid. Anu. Ornitológico Madr. 2001, 2000, 56–67. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V. Emmeans: Estimated Marginal Means, aka Least-Squares Means; R Package, Version 1.6.2-1. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 15 December 2021).
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models; R Package, Version 0.2.0. Available online: https://cran.r-project.org/package=DHARMa (accessed on 15 December 2018).
- Blanco, G.; Cuevas, J.A.; Fargallo, J.A. Breeding density and distribution of choughs Pyrrhocorax pyrrhocorax nesting in river cliffs: The role of nest-site availability. Ardea 1998, 86, 237–244. [Google Scholar]
- Ceballos, P.; Purroy, F.J. Pájaros de Nuestros Campos y Bosques; Mundi-Prensa: Madrid, Spain, 2005. [Google Scholar]
- Fagúndez, J.; Olea, P.P.; Tejedo, P.; Mateo-Tomás, P.; Gómez, D. Irrigation and maize cultivation erode plant diversity within crops in Mediterranean dry cereal agro-ecosystems. Environ. Manag. 2016, 58, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Cabodevilla, X.; Wright, A.D.; Villanua, D.; Arroyo, B.; Zipkin, E.F. The implementation of irrigation leads to declines in farmland birds. Agric. Ecosyst. Environ. 2022, 323, 107701. [Google Scholar] [CrossRef]
- Valcárcel, Y.; Alonso, S.G.; Rodríguez-Gil, J.L.; Gil, A.; Catalá, M. Detection of pharmaceutically active compounds in the rivers and tap water of the Madrid Region (Spain) and potential ecotoxicological risk. Chemosphere 2011, 84, 1336–1348. [Google Scholar] [CrossRef]
- Fernández, M.; Cuesta, S.; Jiménez, O.; Garcıa, M.A.; Hernández, L.M.; Marina, M.L.; González, M.J. Organochlorine and heavy metal residues in the water/sediment system of the southeast regional park in Madrid, Spain. Chemosphere 2000, 41, 801–812. [Google Scholar] [CrossRef]
- Blanco, G.; Sergio, F.; Frías, Ó.; Salinas, P.; Tanferna, A.; Hiraldo, F.; Barceló, D.; Eljarrat, E. Integrating population connectivity into pollution assessment: Overwintering mixing reveals flame retardant contamination in breeding areas in a migratory raptor. Environ. Res. 2018, 166, 553–561. [Google Scholar] [CrossRef]
- Eljarrat, E.; Aznar-Alemany, Ò.; Sala, B.; Frías, Ó.; Blanco, G. Decreasing but still high levels of halogenated flame retardants in wetland birds in central Spain. Chemosphere 2019, 228, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Blanco, G.; Marchamalo, J. Post-breeding inland movements and use of refuse dumps by Audouin’s Gulls in Spain. Waterbirds 1999, 22, 307–309. [Google Scholar] [CrossRef]
- Blanco, G. Can livestock carrion availability influence diet of wintering red kites? Implications of sanitary policies in ecosystem services and conservation. Popul. Ecol. 2014, 56, 593–604. [Google Scholar] [CrossRef]
- Verdú, J.R.; Lobo, J.M.; Sánchez-Piñero, F.; Gallego, B.; Numa, C.; Lumaret, J.P. Ivermectin residues disrupt dung beetle diversity, soil properties and ecosystem functioning: An interdisciplinary field study. Sci. Total Environ. 2018, 618, 219–228. [Google Scholar] [CrossRef]
- Traba, J.; Morales, M.B. The decline of farmland birds in Spain is strongly associated to the loss of fallowland. Sci. Rep. 2019, 9, 9473. [Google Scholar] [CrossRef] [PubMed]
- Bowler, D.E.; Heldbjerg, H.; Fox, A.D.; de Jong, M.; Böhning-Gaese, K. Long-term declines of European insectivorous bird populations and potential causes. Conserv. Biol. 2019, 33, 1120–1130. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Bayo, F.; Wyckhuys, K.A. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Wagner, D.L. Insect declines in the Anthropocene. Annu. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Antia, A.; Ortiz-Santaliestra, M.E.; Mougeot, F.; Camarero, P.R.; Mateo, R. Birds feeding on tebuconazole treated seeds have reduced breeding output. Environ. Pollut. 2021, 271, 116292. [Google Scholar] [CrossRef] [PubMed]
- Meyrier, E.; Jenni, L.; Bötsch, Y.; Strebel, S.; Erne, B.; Tablado, Z. Happy to breed in the city? Urban food resources limit reproductive output in Western Jackdaws. Ecol. Evol. 2017, 7, 1363–1374. [Google Scholar] [CrossRef]
- Plaza, P.I.; Lambertucci, S.A. How are garbage dumps impacting vertebrate demography, health, and conservation? Glob. Ecol. Conserv. 2017, 12, 9–20. [Google Scholar] [CrossRef]
- Bird, J.P.; Martin, R.; Akçakaya, H.R.; Gilroy, J.; Burfield, I.J.; Garnett, S.T.; Symes, A.; Taylor, J.; Şekercioğlu, Ç.H.; Butchart, S.H.M. Generation lengths of the world’s birds and their implications for extinction risk. Conserv. Biol. 2020, 34, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Balmford, A.; Green, R.E.; Jenkins, M. Measuring the changing state of nature. Trends Ecol. Evol. 2003, 18, 326–330. [Google Scholar] [CrossRef]
- Donald, P.F.; Sanderson, F.J.; Burfield, I.J.; Bierman, S.M.; Gregory, R.D.; Waliczky, Z. International conservation policy delivers benefits for birds in Europe. Science 2007, 317, 810–813. [Google Scholar] [CrossRef]
- Amano, T.; Lamming, J.D.L.; Sutherland, W.J. Spatial gaps in global biodiversity information and the role of citizen science. BioScience 2016, 66, 393–400. [Google Scholar] [CrossRef] [Green Version]
- SEO/BirdLife. SEO/BirdLife 2011 Birds Monitoring Programmes; SEO/BirdLife: Madrid, Spain, 2012. [Google Scholar]
- Møller, A.P. Flight distance and population trends in European breeding birds. Behav. Ecol. 2008, 19, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Guthery, F.S. Statistical ritual versus knowledge accrual in wildlife science. J. Wildl. Manag. 2008, 72, 1872–1875. [Google Scholar] [CrossRef]
- Murgi, E. How many common breeding birds are there in Spain? A comparison of census methods and national population size estimates. Ardeola 2011, 58, 343–364. [Google Scholar]
- Bird, T.J.; Bates, A.E.; Lefcheck, J.S.; Hill, N.A.; Thomson, R.J.; Edgar, G.J.; Stuart-Smith, R.D.; Wotherspoon, S.; Krkosek, M.; Stuart-Smith, J.F.; et al. Statistical solutions for error and bias in global citizen science datasets. Biol. Conserv. 2014, 173, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Pellikka, J.; Kuikka, S.; Lindén, H.; Varis, O. The role of game management in wildlife populations: Uncertainty analysis of expert knowledge. Eur. J. Wildl. Res. 2005, 51, 48–59. [Google Scholar] [CrossRef]
- Mustin, K.; Arroyo, B.; Beja, P.; Newey, S.; Irivine, R.J.; Kestler, J.; Redpath, S.M. Consequences of game bird management for non-game species in Europe. J. Appl. Ecol. 2018, 55, 2285–2295. [Google Scholar] [CrossRef] [Green Version]
- Inger, R.; Gregory, R.; Duffy, J.P.; Stott, I.; Voříšek, P.; Gaston, K.J. Common European birds are declining rapidly while less abundant species’ numbers are rising. Ecol. Lett. 2015, 18, 28–36. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
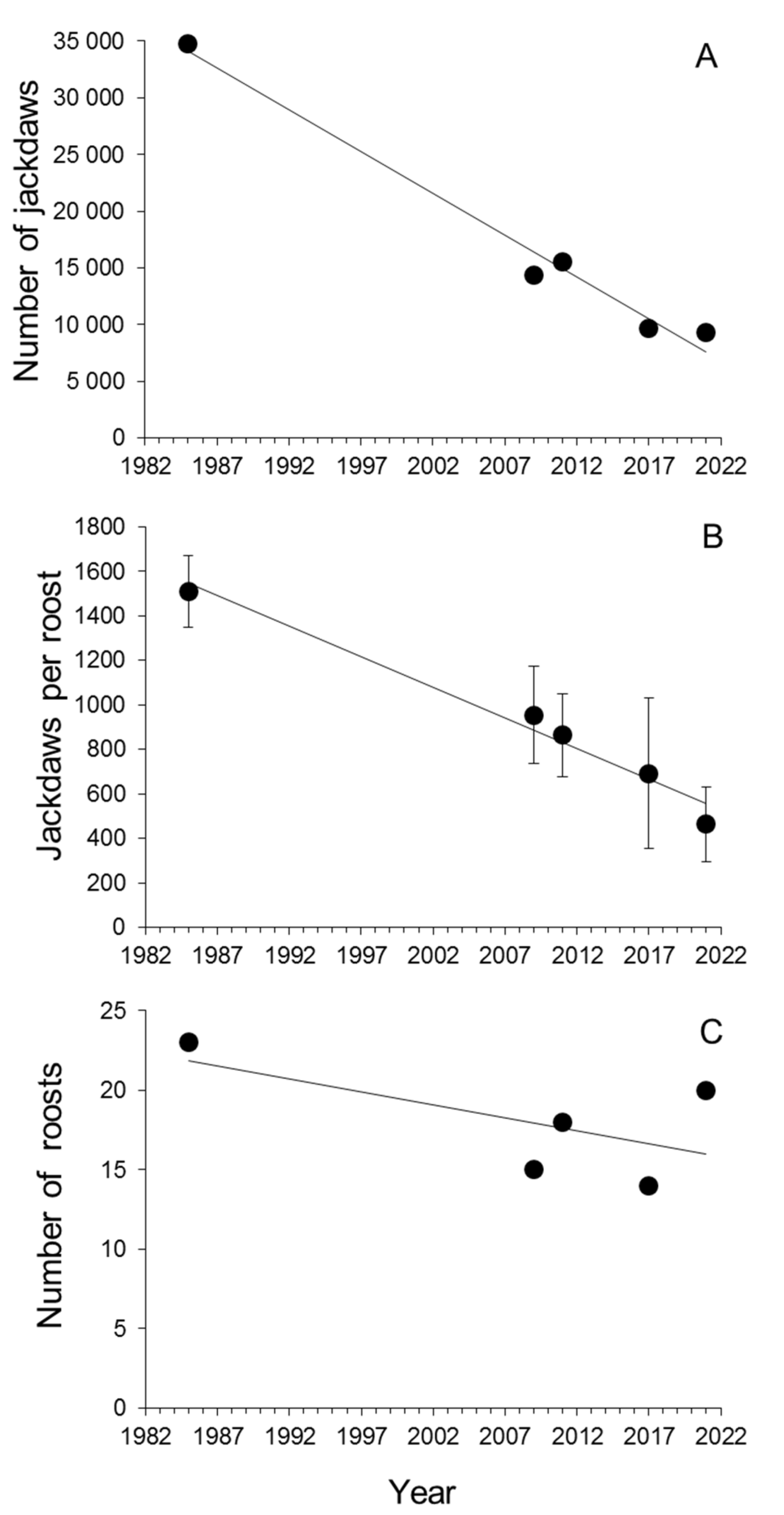{kind=link}
| Total Number of Jackdaws | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| A | χ2 | Df | p Value | Sector | Period | Mean | SE | Df | CI |
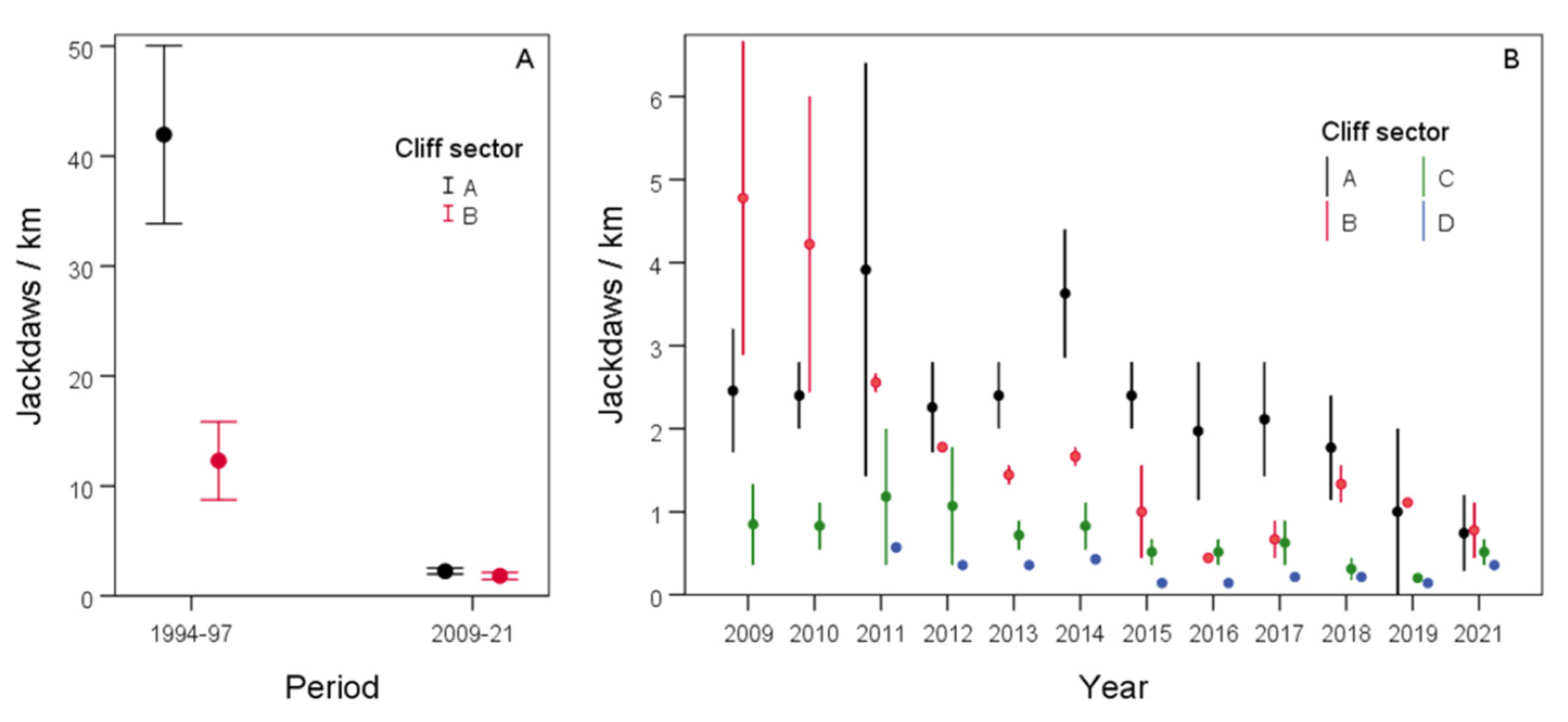
| km | 0.01 | 1 | 0.9031 | A | 1994–1997 | 5.46 | 0.29 | 59 | 4.71–6.22 |
| sector | 4.07 | 1 | 0.0438 | B | 1994–1997 | 4.35 | 0.31 | 59 | 3.54–5.16 |
| period | 66.22 | 1 | 0.0000 | A | 2009–2021 | 1.84 | 0.20 | 59 | 1.31–2.36 |
| sectorXperiod | 14.38 | 1 | 0.0001 | B | 2009–2021 | 2.01 | 0.16 | 59 | 1.61–2.42 |
| B | χ2 | Df | p Value | Sector | Period | Trend | SE | Df | CI |
| km | 12.59 | 1 | 0.0004 | A | 2009–2021 | −0.08 | 0.03 | 72 | −0.15–0.02 |
| sector | 48.77 | 3 | 0.0000 | B | 2009–2021 | −0.18 | 0.03 | 72 | −0.24–0.11 |
| year | 62.18 | 1 | 0.0000 | C | 2009–2021 | −0.09 | 0.03 | 72 | −0.18–0.00 |
| sectorXyear | 8.31 | 3 | 0.0400 | D | 2009–2021 | −0.09 | 0.06 | 72 | −0.23–0.05 |
| Number of Breeding Pairs of Jackdaws | |||||||||
| χ2 | Df | p Value | Sector | Period | Trend | SE | Df | CI | |
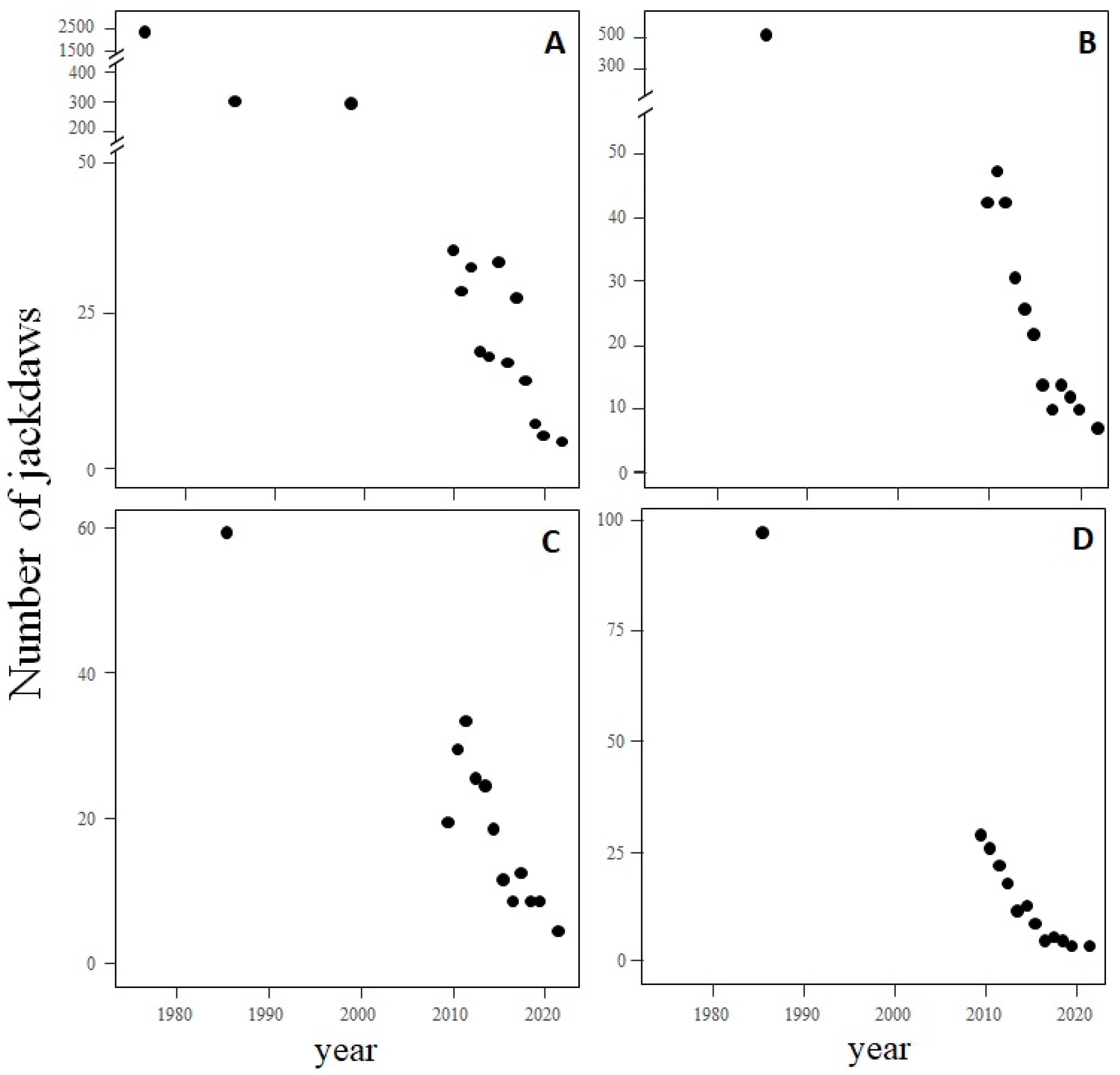
| year | 421.21 | 1 | <0.0001 | A | 1975–2021 | −1.55 | 0.09 | 45 | −1.78–1.31 |
| sector | 28.93 | 3 | <0.0001 | B | 1985–2021 | −1.47 | 0.15 | 45 | −1.86–1.09 |
| sectorXyear | 25.62 | 3 | <0.0001 | C | 1985–2021 | −0.70 | 0.16 | 45 | −1.12–0.29 |
| D | 1985–2021 | −1.03 | 0.16 | 45 | −1.45–0.62 | ||||
| Zone | Number of Colonies | Number of Pairs | ||||
|---|---|---|---|---|---|---|
| 1984–1985 | 2009–2011 | % Lost Colonies | 1984–1985 | 2009–2011 | % Decline | |
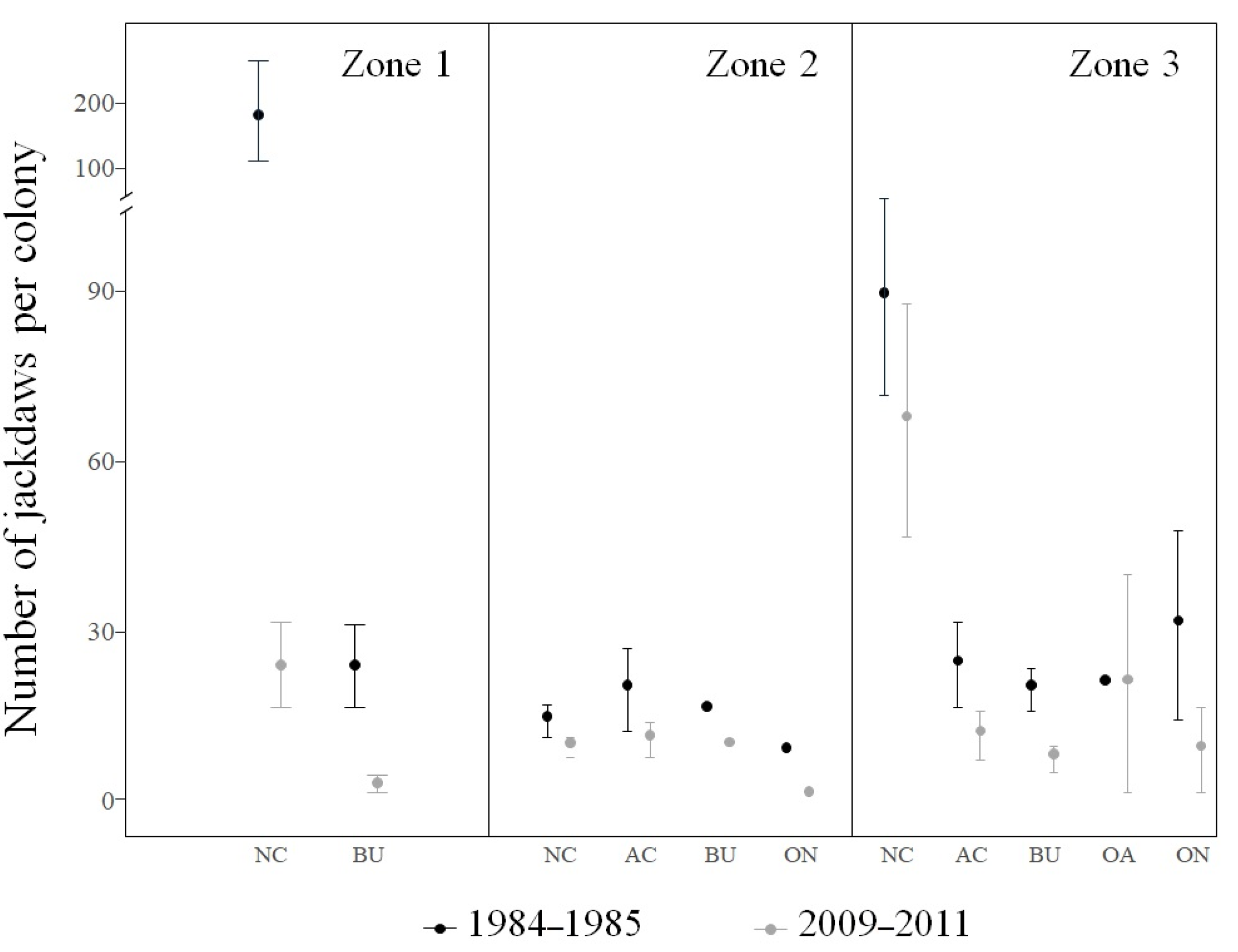
| 1 | 8 | 6 | 25.0 | 1175 | 139 | 88.2 |
| 2 | 13 | 13 | 0.0 | 182 | 102 | 44.0 |
| 3 | 37 | 24 | 35.1 | 1028 | 517 | 49.7 |
| Model | χ2 | Df | p Value |
|---|---|---|---|
| area | 18.91 | 2 | 0.0001 |
| period | 157.82 | 1 | <0.0001 |
| substrate | 19.14 | 4 | 0.0007 |
| areaXperiod | 74.19 | 2 | <0.0001 |
| periodXsubstrate | 17.34 | 4 | 0.0017 |
| Contrasts | Mean (± SE) | Df | CI |
| area 1, 1984–1985 | 4.00 (± 0.38) | 99 | 2.98–5.01 |
| area 1, 2009–2011 | 1.98 (± 0.41) | 99 | 0.88–3.08 |
| area 2, 1984–1985 | 2.03 (± 0.31) | 99 | 1.19–2.86 |
| area 2, 2009–2012 | 1.52 (± 0.33) | 99 | 0.62–2.42 |
| area 3, 1984–1985 | 3.14 (± 0.19) | 99 | 2.62–3.66 |
| area 3, 2009–2013 | 2.72 (± 0.21) | 99 | 2.15–3.30 |
| Contrasts | Mean (± SE) | Df | CI |
| NC, 1984–1985 | 3.79 (± 0.21) | 99 | 3.19–4.39 |
| NC, 2009–2012 | 2.85 (± 0.21) | 99 | 2.24–3.47 |
| AC, 1984–1985 | 3.23 (± 0.36) | 99 | 2.21–4.25 |
| AC, 2009–2012 | 2.07 (± 0.37) | 99 | 1.00–3.15 |
| BU, 1984–1985 | 2.44 (± 0.24) | 99 | 1.74–3.14 |
| BU, 2009–2012 | 1.21 (± 0.27) | 99 | 0.45–2.00 |
| OA, 1984–1985 | 2.90 (± 0.65) | 99 | 1.03–4.78 |
| OA, 2009–2012 | 3.01 (± 0.69) | 99 | 1.03–5.00 |
| ON, 1984–1985 | 2.92 (± 0.43) | 99 | 1.68–4.15 |
| ON, 2009–2012 | 1.23 (± 0.55) | 99 | −0.36–2.81 |
| 1996 | 2010 | |||||
|---|---|---|---|---|---|---|
| River | % Positive | n | Density | % Positive | n | Density |
| Tajo | 15 | 80 | 3.98 | 0 | 34 | 0 |
| Tajuña | 3.2 | 31 | 0.82 | 0 | 27 | 0 |
| Jarama | 16.7 | 30 | 5.52 | 0 | 45 | 0 |
| Guadarrama | 11.1 | 63 | 2.02 | 0 | 25 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco, G.; Domínguez, L.; Fernández, L.; Martínez, F.; González del Barrio, J.L.; Frías, Ó.; Cuevas, J.A.; Carrete, M. The Decline of Common Birds Exemplified by the Western Jackdaw Warns on Strong Environmental Degradation. Conservation 2022, 2, 80-96. https://doi.org/10.3390/conservation2010007
Blanco G, Domínguez L, Fernández L, Martínez F, González del Barrio JL, Frías Ó, Cuevas JA, Carrete M. The Decline of Common Birds Exemplified by the Western Jackdaw Warns on Strong Environmental Degradation. Conservation. 2022; 2(1):80-96. https://doi.org/10.3390/conservation2010007
Chicago/Turabian StyleBlanco, Guillermo, Luis Domínguez, Luis Fernández, Félix Martínez, José L. González del Barrio, Óscar Frías, Jesús A. Cuevas, and Martina Carrete. 2022. "The Decline of Common Birds Exemplified by the Western Jackdaw Warns on Strong Environmental Degradation" Conservation 2, no. 1: 80-96. https://doi.org/10.3390/conservation2010007
APA StyleBlanco, G., Domínguez, L., Fernández, L., Martínez, F., González del Barrio, J. L., Frías, Ó., Cuevas, J. A., & Carrete, M. (2022). The Decline of Common Birds Exemplified by the Western Jackdaw Warns on Strong Environmental Degradation. Conservation, 2(1), 80-96. https://doi.org/10.3390/conservation2010007