
Ontogenetic Habitat Use and Density of the Green Lizard (Lacerta bilineata) in Contrasted Landscapes in France and Italy




Abstract
:1. Introduction
- (1)
- Are there density differences between L. bilineata populations inhabiting contrasted habitats (mixed forest–grassland mosaics versus hedgerows within an agricultural matrix)? This question is important in order to better define the characteristics of the sites that are most suitable for L. bilineata ecological needs, and therefore for defining more precisely the management options for this declining lizard species.
- (2)
- Do L. bilineata individuals select the same habitat types or is there significant intrapopulation variation in habitat selection? That is, are there significant habitat differences between sexes and age classes? If so, what are the ecological reasons behind these intraspecific differences? These questions are important, because habitat management can generate variation in a cascade of organismal traits linked to the environment [18], and knowing the intraspecific habitat use variation may better guide management strategies. In addition, efficient management for conservation of a given species may require a more integrated approach in case of substantial intraspecific (intersexual or age-related) differences in habitat use [17,18]. In this case, the green lizard may be a very suitable study case, as cannibalistic habits have been already detected in this species, and cannibalism may lead to habitat shifts by age or by sex [8].
- (3)
- Are human-made structures important for the habitat requirements of L. bilineata or are there specific plant species (bushes or trees) that are positively correlated with the species’ presence? This question is important in order to better define whether active human intervention (replanting of given species or building of specific features, for instance stony walls, etc.) may help management of the suitable habitat for the study species, or whether passive management (that is just natural habitat monitoring) may be a sufficient condition for the effective conservation of L. bilineata populations.
2. Materials and Methods
2.1. Study Area
2.1.1. Italy
2.1.2. France
2.2. Protocol
2.2.1. Italy
2.2.2. France
2.3. Statistical Analyses
t = s/√∑ (x − mean)2
3. Results
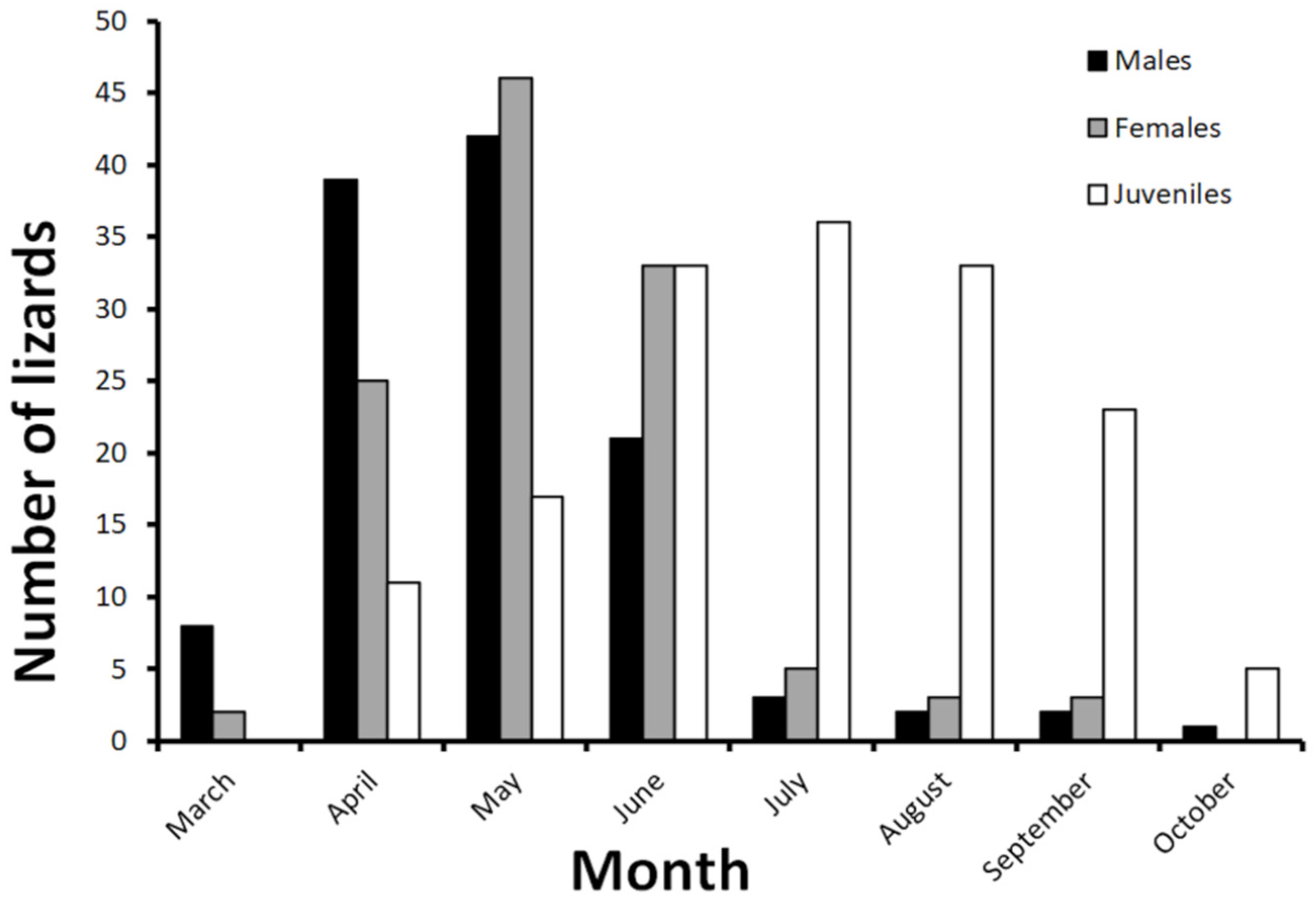
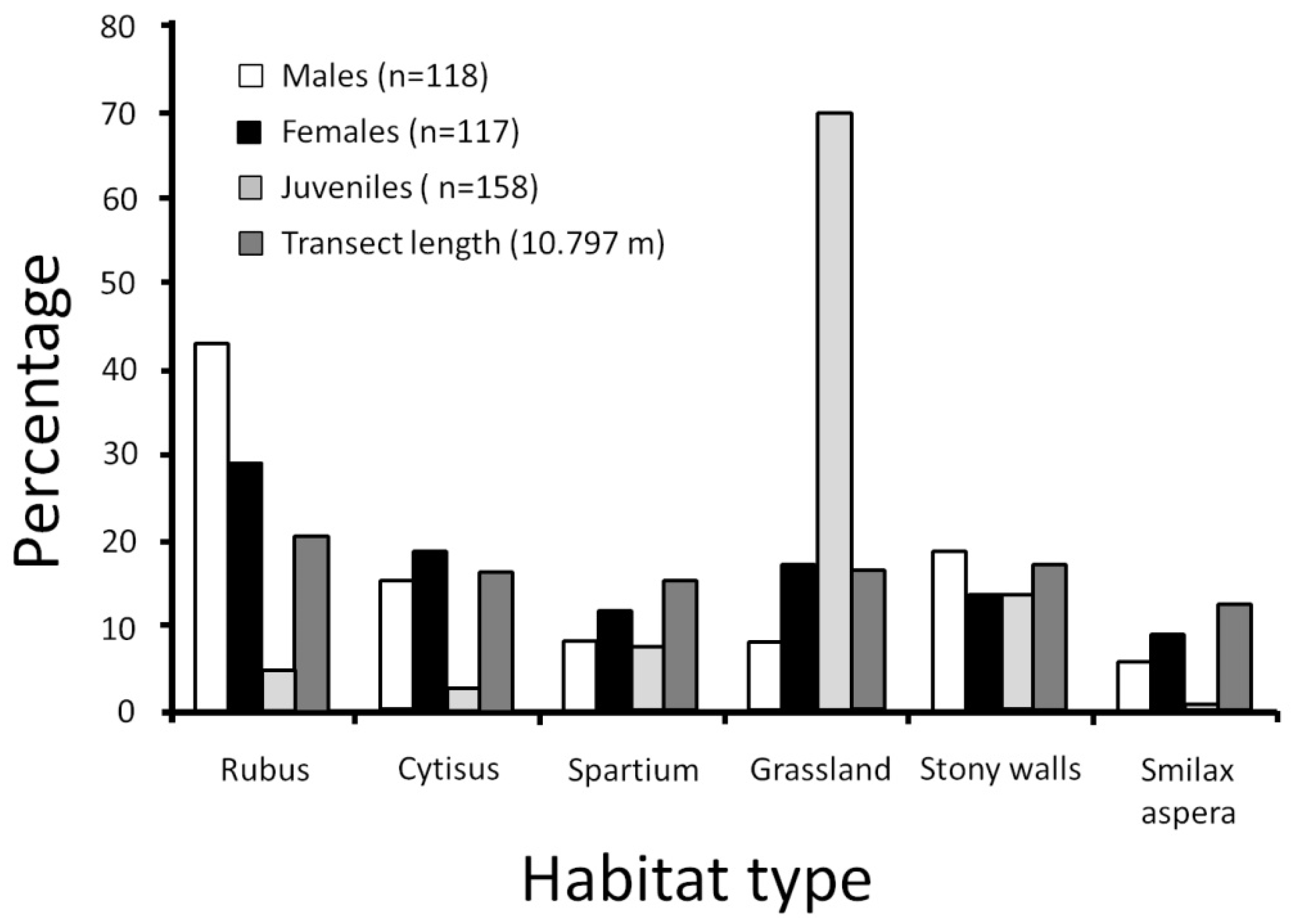
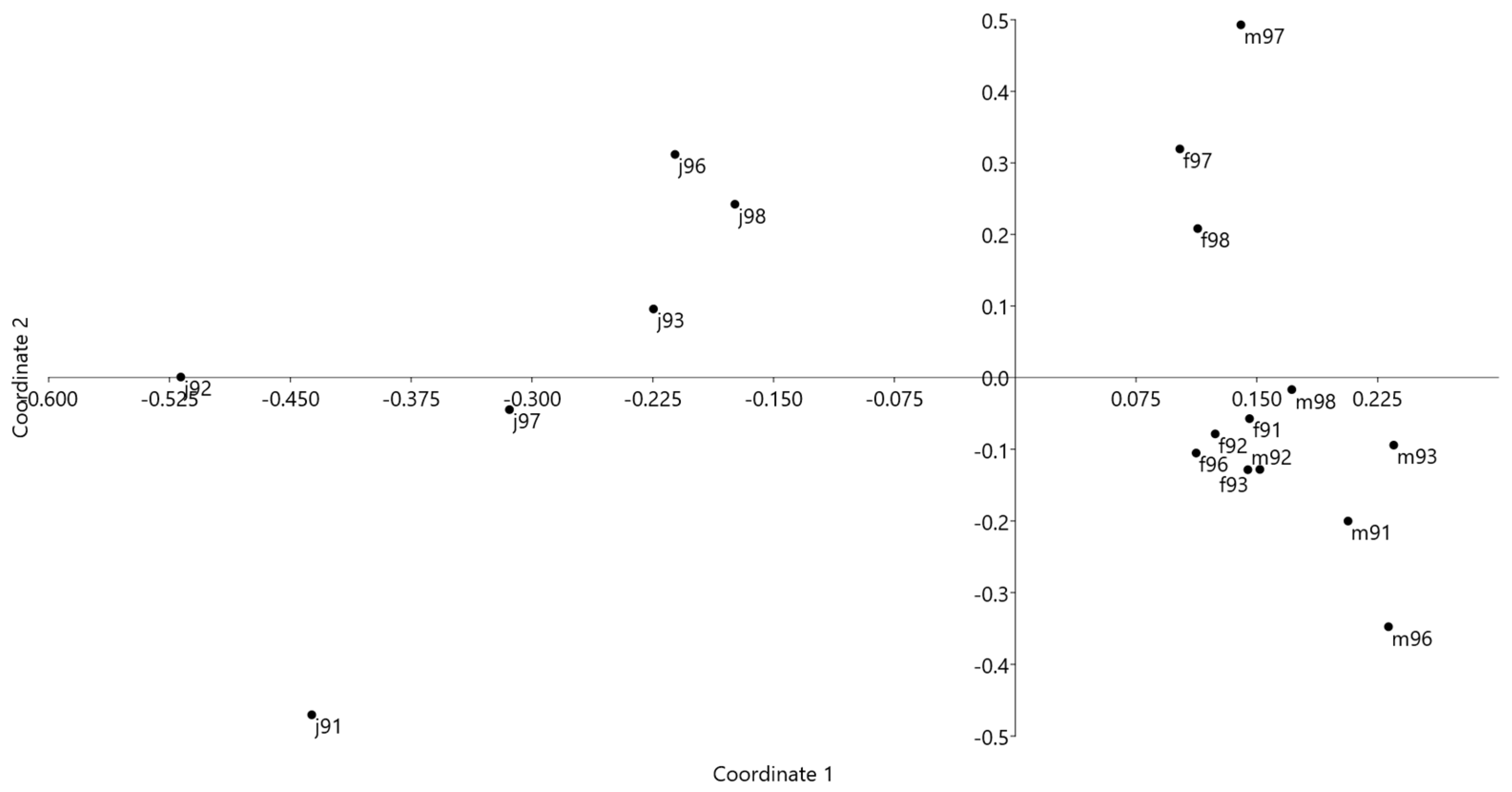
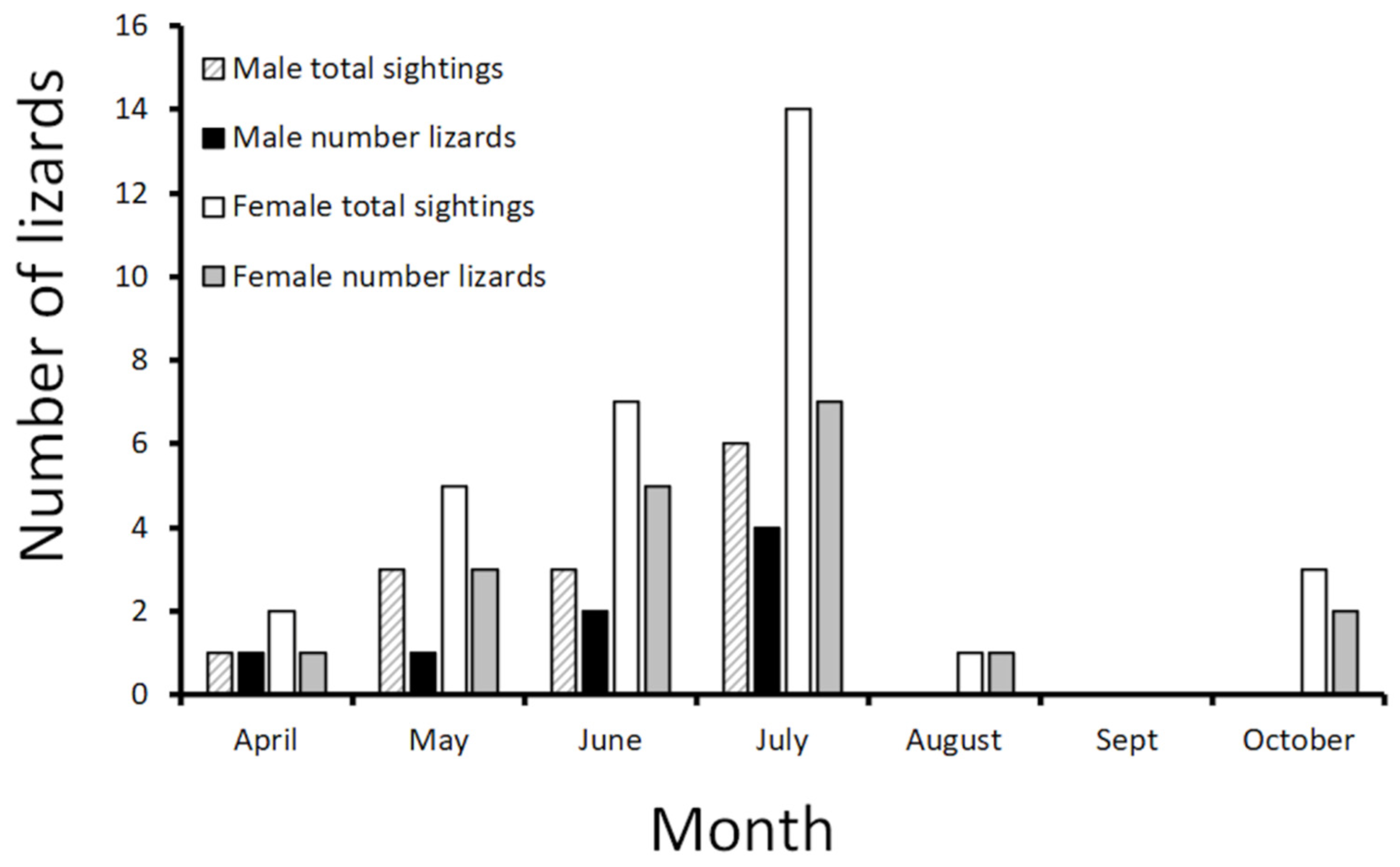
3.1. Italy
3.2. France
4. Discussion
4.1. Ecological Considerations
4.2. Conservation Considerations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doherty, T.S.; Balouch, S.; Bell, K.; Burns, T.J.; Feldman, A.; Fist, C.; Garvey, T.F.; Jessop, T.S.; Meiri, S.; Driscoll, D.A. Reptile responses to anthropogenic habitat modification: A global meta-analysis. Glob. Ecol. Biogeog. 2020, 29, 1265–1279. [Google Scholar] [CrossRef]
- Luiselli, L.; Vignoli, L.; Rugiero, L.; Meek, R. Declining occupancy rates in the hibernacula of aspic vipers (Vipera aspis) in Italy and France; evidence for climatic effects? Herpetol. J. 2018, 28, 137–142. [Google Scholar]
- Pérez-Mellado, V.; Cheylan, M.; Geniez, P.; Nettmann, H.K.; Schmidt, B.; Podloucky, R.; Sindaco, R.; Romano, A. Lacerta bilineata. In IUCN 2012. IUCN Red List of Threatened Species; Version 2012.2.; IUCN: Gland, Switzerland, 2009; Available online: www.iucnredlist.org (accessed on 14 December 2020).
- Sacchi, R.; Marchesi, M.; Gentilli, A.; Pellitteri-Rosa, D.; Scali, S.; Borelli, A. Western green lizards (Lacerta bilineata) do not select the composition or structure of the ecotones in Northern Italy. North West. J. Zool. 2011, 7, 213–221. [Google Scholar]
- Pernat, A.; Sellier, Y.; Préau, C.; Beaune, D. Effet du pâturage sur le lézard vert occidental (Lacerta bilineata Daudin, 1802) (Squamata: Lacertidae) en milieu de landes. Bull. Soc. Herpétol. Fr. 2017, 161, 57–66. [Google Scholar]
- Luppi, M.; Gentilli, A.; Bogliani, G. Microhabitat selection of the Western green lizard Lacerta bilineata. Nat. Hist. Sci. 2020, 7, 3–10. [Google Scholar] [CrossRef]
- Verwaijen, D.; Van Damme, R. Foraging mode and its flexibility in Lacertid lizards from Europe. J. Herpetol. 2008, 42, 124–133. [Google Scholar] [CrossRef]
- Angelici, F.M.; Luiselli, L.; Rugiero, L. Food habits of the green lizard, Lacerta bilineata, in central Italy and a reliability test of faecal pellet analysis. Ital. J. Zool. 1997, 64, 267–272. [Google Scholar] [CrossRef]
- Meek, R. Temporal trends in Podarcis muralis and Lacerta bilineata populations in a fragmented landscape in western France: Results from a 14 year time series. Herpetol. J. 2020, 30, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Luiselli, L.; Rugiero, L. Null model analysis of lizard communities in five urban parks of Rome. Amphib. Reptil. 2007, 28, 547–553. [Google Scholar] [CrossRef]
- Meek, R. Temporal distributions, habitat associations and behaviour of the green lizard (Lacerta bilineata) and wall lizard (Podarcis muralis) on roads in a fragmented landscape in Western France. Acta Herpetol. 2014, 9, 179–186. [Google Scholar]
- Rugiero, L.; Capula, M.; Capizzi, D.; Amori, G.; Milana, G.; Lai, M.; Luiselli, L. Long-term observations on the number of roadkilled Zamenis longissimus (Laurenti, 1768) in a hilly area of central Italy. Herpetozoa 2018, 30, 212–217. [Google Scholar]
- Rugiero, L.; Milana, G.; Capula, M.; Amori, G.; Luiselli, L. Long term variations in small mammal composition of a snake diet do not mirror climate change trends. Acta Oecol. 2012, 43, 158–164. [Google Scholar] [CrossRef]
- Rugiero, L.; Milana, G.; Petrozzi, F.; Capula, M.; Luiselli, L. Climate-change-related shifts in annual phenology of a temperate snake during the last 20 years. Acta Oecol. 2013, 51, 42–48. [Google Scholar] [CrossRef]
- Capula, M.; Rugiero, L.; Capizzi, D.; Franco, D.; Milana, G.; Luiselli, L. Long-term, climate-change-related shifts in feeding frequencies of a Mediterranean snake population. Ecol. Res. 2016, 31, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Capula, M.; Rugiero, L.; Capizzi, D.; Milana, G.; Vignoli, L.; Franco, D.; Petrozzi, F.; Luiselli, L. Long-term, climate change-related shifts in monthly patterns of roadkilled Mediterranean snakes (Hierophis viridiflavus). Herpetol. J. 2014, 24, 97–102. [Google Scholar]
- Luiselli, L.; Capizzi, D. Influences of area, isolation and habitat features on distribution of snakes in Mediterranean fragmented woodlands. Biodiv. Conserv. 1997, 6, 1339–1351. [Google Scholar] [CrossRef]
- McBrayer, L.D.; Parker, S.E. Variation in habitat management alters risk aversion behavior in lizards. Behav. Ecol. Sociobiol. 2018, 72, 1–9. [Google Scholar] [CrossRef]
- Bonnet, X.; Billy, G.; Lakušić, M. Puncture versus capture: Which stresses animals the most? J. Comp. Physiol. B 2020. [Google Scholar] [CrossRef]
- Rugiero, L.; Capula, M.; Dendi, D.; Petrozzi, F.; Di Vittorio, M.; Luiselli, L. A 28-years-long monitoring study suggests heavy decline and habitat shifts in three populations of a Mediterranean tortoise species. Anim. Biol. 2021, 71, 21–35. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression Analysis, 2nd ed.; John Wiley and Sons: Hoboken, NJ, USA, 2000. [Google Scholar]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models; Chapman and Hall/CRC: London, UK, 1989. [Google Scholar]
- Pianka, E.R. Niche overlap and diffuse competition. Proc. Natl. Acad. Sci. USA 1974, 71, 2141–2145. [Google Scholar] [CrossRef] [Green Version]
- Bailey, N.T.J. Statistical Methods in Biology; Cambridge University Press: Cambridge, UK, 1995; p. 255. [Google Scholar]
- Paulissen, M.A. Ontogenetic and seasonal shifts in microhabitat use by the lizard Cnemidophorus sexlineatus. Copeia 1988, 1988, 1021–1029. [Google Scholar] [CrossRef]
- Irschick, D.J.; Macrini, T.E.; Koruba, S.; Forman, J. Ontogenetic differences in morphology, habitat use, behavior, and sprinting capacity in two West Indian Anolis lizards. J. Herpetol. 2000, 34, 444–451. [Google Scholar] [CrossRef]
- Smith, G.R.; Ballinger, R.E. The ecological consequences of habitat and microhabitat use in lizards: A review. Contemp. Herpetol. 2001, 3, 1–27. [Google Scholar] [CrossRef]
- Law, B.S. Habitat Shift in the Eastern Australian Water Skink (Eulamprus quoyii)? Copeia 1991, 1991, 1117–1120. [Google Scholar] [CrossRef]
- Jenssen, T.A.; Hovde, K.A.; Taney, K.G. Size-related habitat use by nonbreeding Anolis carolinensis lizards. Copeia 1998, 1998, 774–779. [Google Scholar] [CrossRef]
- Nemes, S.; Vogrin, M.; Hartel, T.; Ölerer, K. Habitat selection at the sand lizard (Lacerta agilis): Ontogenetic shifts. North-West. J. Zool. 2006, 2, 17–26. [Google Scholar]
- Keren-Rotem, T.; Bouskila, A.; Geffen, E. Ontogenetic habitat shift and risk of cannibalism in the common chameleon (Chamaeleo chamaeleon). Behav. Ecol. Sociobiol. 2006, 59, 723–731. [Google Scholar] [CrossRef]
- Imansyah, M.J.; Jessop, T.S.; Ciofi, C.; Akbar, Z. Ontogenetic differences in the spatial ecology of immature Komodo dragons. J. Zool. 2007. [Google Scholar] [CrossRef]
- Kolanek, A.; Bury, S.; Turniak, E.; Szymanowski, M. Age-dependent utilization of shelters and habitat in two reptile species with contrasting intraspecific interactions. Animals 2019, 9, 995. [Google Scholar] [CrossRef] [Green Version]
- Powell, G.L.; Russell, A.P. Locomotor correlates of ecomorph designation in Anolis: An examination of three sympatric species from Jamaica. Can. J. Zool. 1992, 70, 725–727. [Google Scholar] [CrossRef]
- Andrews, R.M. Growth rate in island and mainland lizards. Copeia 1976, 1976, 477–482. [Google Scholar] [CrossRef]
- Lathi, M.E.; Beck, D.D. Ecology and ontogenetic variation of diet in the pigmy short-horned lizard (Phrynosoma douglasii). Am. Midl. Nat. 2008, 159, 327–339. [Google Scholar]
- Le Galliard, J.-F.; Fitze, P.S.; Ferriere, R.; Clobert, J. Sex ratio bias, male aggression, and population collapse in lizards. Proc. Natl. Acad. Sci. USA 2005, 102, 18231–18236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meek, R. Natural History Note; Lacerta bilineata (western green lizard): Field injury. Herpetol. Bull. 2011, 116, 40–41. [Google Scholar]
- Mitchell, J.C. Cannibalism in reptiles: A worldview review. St. Amphib. Reptiles Herpetol. Cir. 1986, 15, 1–23. [Google Scholar]
- Beebee, T.J.C.; Griffiths, R.A. Amphibians and Reptiles; Harper-Collins: London, UK, 2000. [Google Scholar]
- Stamps, J.A. The relationship between onto- genetic habitat shifts, competition and predator avoidance in a juvenile lizard (Anolis aeneus). Behav. Ecol. Sociobiol. 1983, 12, 19–33. [Google Scholar] [CrossRef]
- Pounds, J.A. Ecomorphology, locomotion, and microhabitat structure: Patterns in a tropical mainland Anolis community. Ecol. Monogr. 1988, 58, 299–320. [Google Scholar] [CrossRef]
- Simberloff, D.; Cox, J. Consequences and costs of conservation corridors. Conserv. Biol. 1987, 1, 63–71. [Google Scholar] [CrossRef]
- Spellerberg, I.F. The Ecological Effects of Roads; Science Publishers: Enfield, NH, USA, 2002. [Google Scholar]
- Davies, Z.G.; Pullin, A.S. Are hedgerows effective corridors between fragments of woodland habitat? An evidence-based approach. Landsc. Ecol. 1987, 22, 333–351. [Google Scholar] [CrossRef]
- Delgado Garcia, J.D.; Arevalo, J.R.; Fernandez-Palacios, J.M. Road edge effect on the abundance of the lizard Gallotia gallotia in two Canary Islands forests. Biodivers. Conserv. 2007, 16, 2949–2963. [Google Scholar] [CrossRef]
- Gherghel, I.; Strugariu, A.; Sahlean, T.C.; Zamfirescu, O. Anthropogenic impact or anthropogenic accommodation? Distribution expansion of the common wall lizard (Podarcis muralis) by artificial habitats in the north-eastern limits of its distribution range. Acta Herpetol. 2009, 4, 183–189. [Google Scholar]
- Saint Girons, H. Structure et évolution d’une petite population de Vipera aspis (L.) dans une région de bocage de l’ouest de la France. Rev. Écol. 1996, 61, 223–241. [Google Scholar]
- Saint Girons, H.; Bradshaw, S.D. Sédentarité, déplacements et répartition des individus dans une population de Lacerta viridis (Laurenti, 1768) (Lacertilia, Lacertidae). Bijdr. Dierkd. 1989, 59, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Saint-Sirons, H.; Castanet, J.; Bradshaw, S.D.; Baron, J.P. Démographie comparée de deux populations françaises de Lacerta viridis (Laurenti, 1768). Rev. Écol. 1989, 44, 361–386. [Google Scholar]
- Michaelides, G.; Kati, V. Diversity patterns and conservation management of the lizard community in a Mediterranean reserve (Cyprus). J. Biol. Res. Thessalon. 2009, 12, 211–220. [Google Scholar]
- Lecq, S.; Loisel, A.; Brischoux, F.; Mullin, S.J.; Bonnet, X. Importance of ground refuges for the biodiversity in agricultural hedgerows. Ecol. Ind. 2017, 72, 615–626. [Google Scholar] [CrossRef]
- Lecq, S.; Loisel, A.; Mullin, S.J.; Bonnet, X. Manipulating hedgerow quality: Embankment size influences animal biodiversity in a peri-urban context. Urban For. Urban Green. 2018, 35, 1–7. [Google Scholar] [CrossRef]
- Edgar, P.; Foster, J.; Baker, J. Reptile Habitat Management Handbook; Amphibian and Reptile Conservation: Bournemouth, UK, 2010. [Google Scholar]
- Prieto-Ramirez, A.M.; Röhler, L.; Cord, A.F.; Péer, G.; Rödder, D.; Henle, K. Differential effects of habitat loss on occupancy patterns of the eastern green lizard Lacerta viridis at the core and periphery of its distribution range. PLoS ONE 2020, 15, e0229600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, T.; Stille, B.; Shine, R. Inbreeding depression in an isolated population of adders Vipera berus. Biol. Conserv. 1996, 75, 113–118. [Google Scholar] [CrossRef]
- Miller, K.A.; Chapple, D.G.; Towns, D.R.; Ritchie, P.A.; Nelson, N.J. Assessing genetic diversity for conservation management: A case study of a threatened reptile. Anim. Conserv. 2009, 12, 163–171. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat Types | Total | ||||||
|---|---|---|---|---|---|---|---|
| 1991 | Rubus | Cytisus | Spartium | grassland | stony walls | Smilax aspera | |
| transect length (m) | 416 | 339 | 255 | 410 | 350 | 235 | 2005 |
| males (n) | 11 | 3 | 1 | 2 | 6 | 3 | 26 |
| females (n) | 7 | 3 | 3 | 3 | 4 | 4 | 24 |
| juveniles (n) | 2 | 6 | 7 | 24 | 3 | 1 | 43 |
| 1992 | |||||||
| transect length (m) | 339 | 245 | 270 | 337 | 411 | 190 | 1792 |
| males (n) | 8 | 4 | 3 | 3 | 4 | 1 | 23 |
| females (n) | 8 | 2 | 3 | 4 | 2 | 2 | 21 |
| juveniles (n) | 0 | 0 | 1 | 26 | 3 | 0 | 30 |
| 1993 | |||||||
| transect length (m) | 402 | 330 | 308 | 294 | 332 | 208 | 1874 |
| males (n) | 8 | 6 | 2 | 0 | 2 | 1 | 19 |
| females (n) | 6 | 8 | 2 | 3 | 3 | 1 | 23 |
| juveniles (n) | 1 | 2 | 2 | 15 | 4 | 0 | 24 |
| 1996 | |||||||
| transect length (m) | 320 | 275 | 290 | 255 | 285 | 225 | 1650 |
| males (n) | 14 | 3 | 1 | 2 | 4 | 1 | 25 |
| females (n) | 6 | 5 | 4 | 4 | 2 | 2 | 23 |
| juveniles (n) | 3 | 0 | 2 | 19 | 6 | 0 | 30 |
| 1997 | |||||||
| transect length (m) | 402 | 302 | 267 | 281 | 256 | 244 | 1752 |
| males (n) | 2 | 0 | 0 | 1 | 4 | 1 | 8 |
| females (n) | 3 | 1 | 1 | 3 | 3 | 2 | 13 |
| juveniles (n) | 0 | 0 | 0 | 14 | 3 | 0 | 17 |
| 1998 | |||||||
| transect length (m) | 355 | 315 | 274 | 245 | 260 | 275 | 1724 |
| males (n) | 8 | 2 | 3 | 2 | 2 | 0 | 17 |
| females (n) | 4 | 3 | 1 | 3 | 2 | 0 | 13 |
| juveniles (n) | 1 | 1 | 0 | 13 | 3 | 0 | 18 |
| Year | Males vs. Females | Males vs. Juveniles | Females vs. Juveniles |
|---|---|---|---|
| 1991 | 0.939 | 0.333 | 0.522 |
| 1992 | 0.956 | 0.331 | 0.439 |
| 1993 | 0.924 | 0.193 | 0.473 |
| 1996 | 0.799 | 0.347 | 0.560 |
| 1997 | 0.854 | 0.387 | 0.620 |
| 1998 | 0.885 | 0.339 | 0.621 |
| Estimate | St. Error | Wald | p | ||
|---|---|---|---|---|---|
| Male | Intercept | 2.242248 | 0.271097 | 68.40954 | 0.000000 |
| Rubus sp. | 0.081649 | 0.026769 | 9.30322 | 0.002288 | |
| Female | Intercept | 5.474814 | 5.589709 | 0.959313 | 0.327360 |
| Rubus sp. | 2.475033 | 1.008660 | 6.021048 | 0.014136 | |
| Juveniles | Intercept | 19.88394 | 2.535319 | 61.50914 | 0.000000 |
| Spartium sp. | 3.55803 | 1.036835 | 11.77605 | 0.000600 |
| Year | Transect Length (m) | Number of Individuals | Density ha−1 |
|---|---|---|---|
| 1991 | 2005 | 93 | 23 |
| 1992 | 1792 | 74 | 20 |
| 1993 | 1874 | 66 | 18 |
| 1996 | 1650 | 78 | 23 |
| 1997 | 1752 | 88 | 25 |
| 1998 | 1724 | 48 | 14 |
| April | May | June | July | August | Sept | October | |
|---|---|---|---|---|---|---|---|
| Male 1 | x | xxx | xx | ||||
| Male 2 | x | ||||||
| Male 3 | xx | ||||||
| Male 4 | x | ||||||
| Male 5 | xx | ||||||
| Male 6 | x | ||||||
| Female 1 | x | x | xxxx | ||||
| Female 2 | xx | x | xx | x | |||
| Female 3 | xx | xx | |||||
| Female 4 | x | xx | xxx | ||||
| Female 5 | x | x | |||||
| Female 6 | xx | ||||||
| Female 7 | x | ||||||
| Female 8 | x | ||||||
| Female 9 | x | ||||||
| Female 10 | xx |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rugiero, L.; Capula, M.; Di Vittorio, M.; Dendi, D.; Meek, R.; Luiselli, L. Ontogenetic Habitat Use and Density of the Green Lizard (Lacerta bilineata) in Contrasted Landscapes in France and Italy. Conservation 2021, 1, 1-16. https://doi.org/10.3390/conservation1010001
Rugiero L, Capula M, Di Vittorio M, Dendi D, Meek R, Luiselli L. Ontogenetic Habitat Use and Density of the Green Lizard (Lacerta bilineata) in Contrasted Landscapes in France and Italy. Conservation. 2021; 1(1):1-16. https://doi.org/10.3390/conservation1010001
Chicago/Turabian StyleRugiero, Lorenzo, Massimo Capula, Massimiliano Di Vittorio, Daniele Dendi, Roger Meek, and Luca Luiselli. 2021. "Ontogenetic Habitat Use and Density of the Green Lizard (Lacerta bilineata) in Contrasted Landscapes in France and Italy" Conservation 1, no. 1: 1-16. https://doi.org/10.3390/conservation1010001
APA StyleRugiero, L., Capula, M., Di Vittorio, M., Dendi, D., Meek, R., & Luiselli, L. (2021). Ontogenetic Habitat Use and Density of the Green Lizard (Lacerta bilineata) in Contrasted Landscapes in France and Italy. Conservation, 1(1), 1-16. https://doi.org/10.3390/conservation1010001