

Effect of Salinity and Nitrogen on Heavy Metal Tolerance and Accumulation Potential in Rumex maritimus

 , , and
, , and 
Abstract
1. Introduction
2. Results
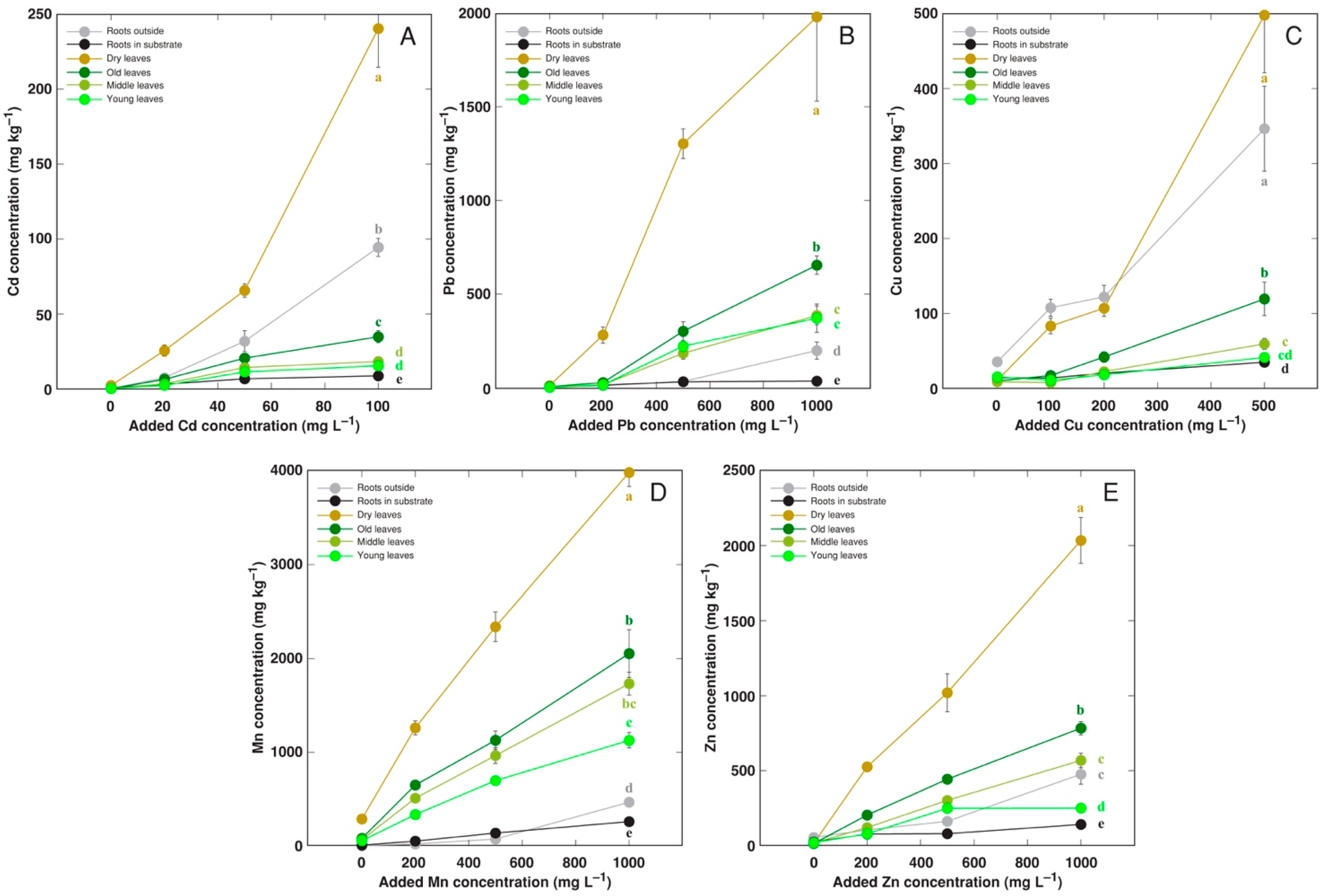
2.1. Experiment 1: Gradual Treatment with Cd, Pb, Cu, Mn, and Zn
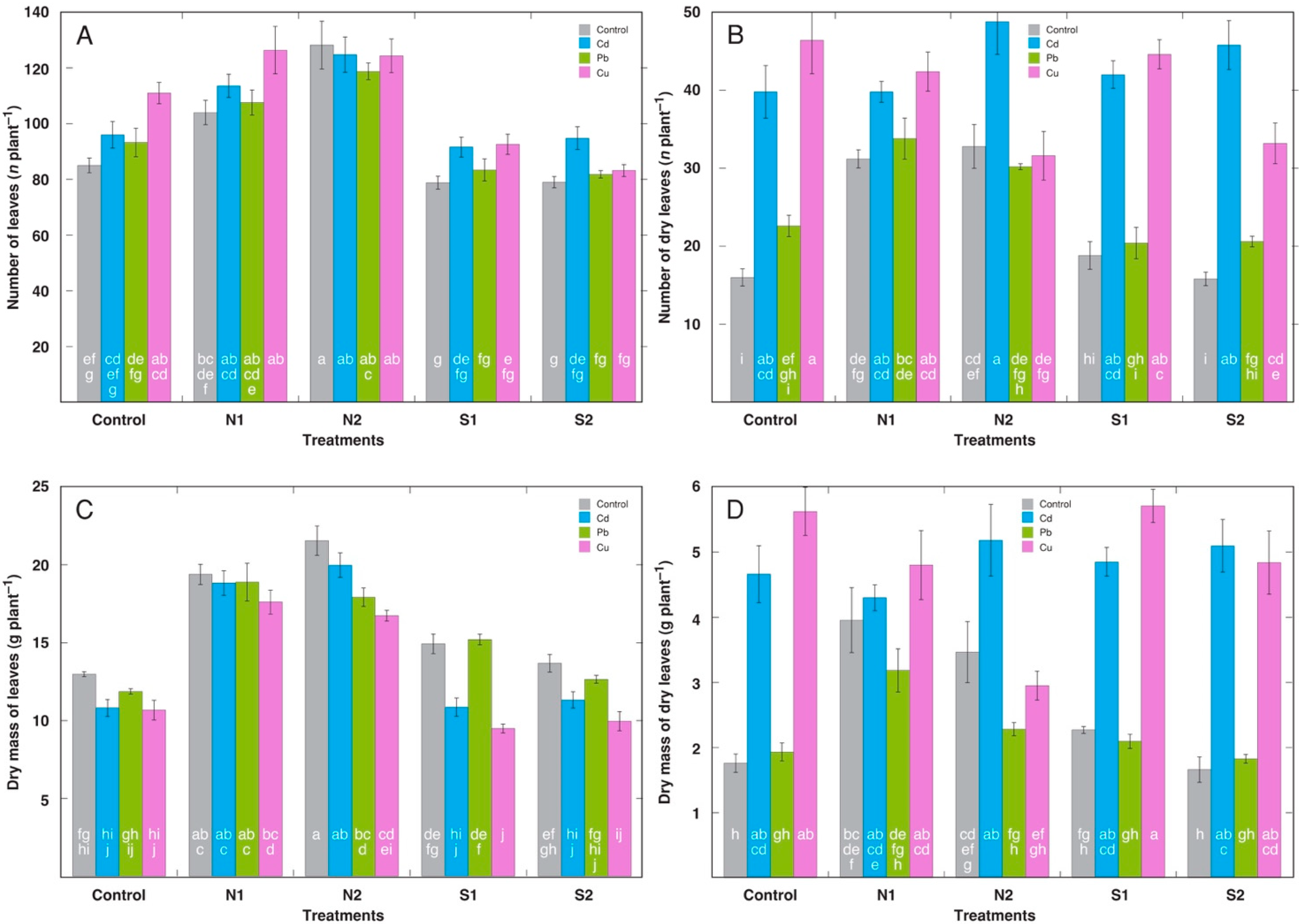
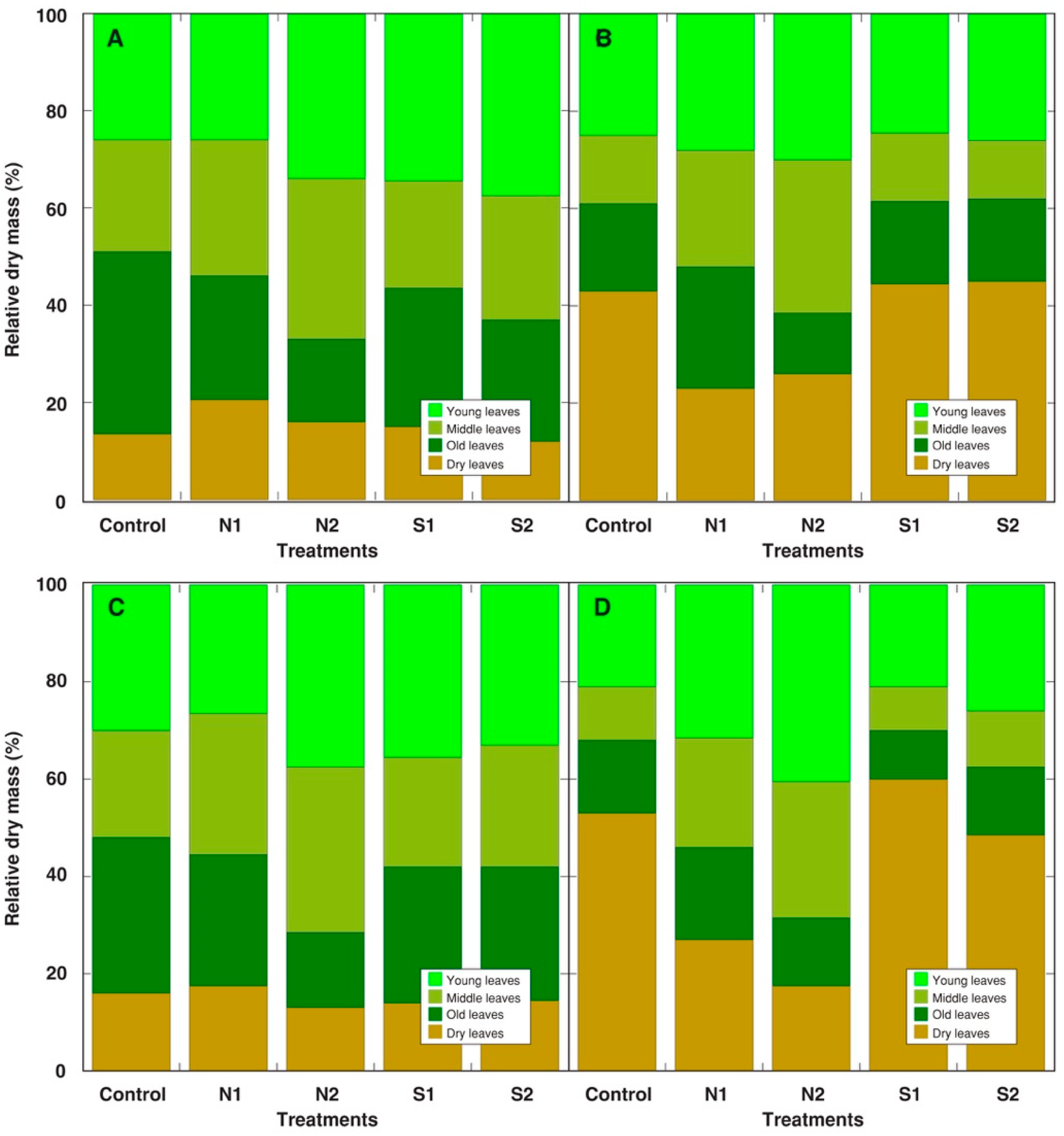
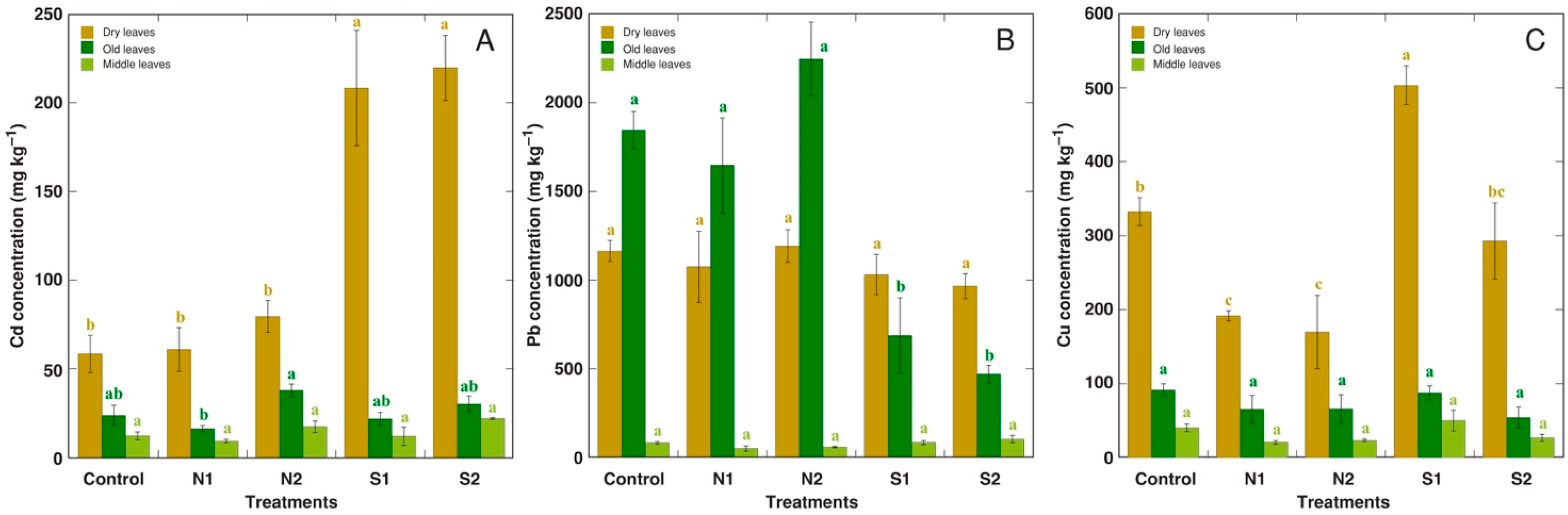
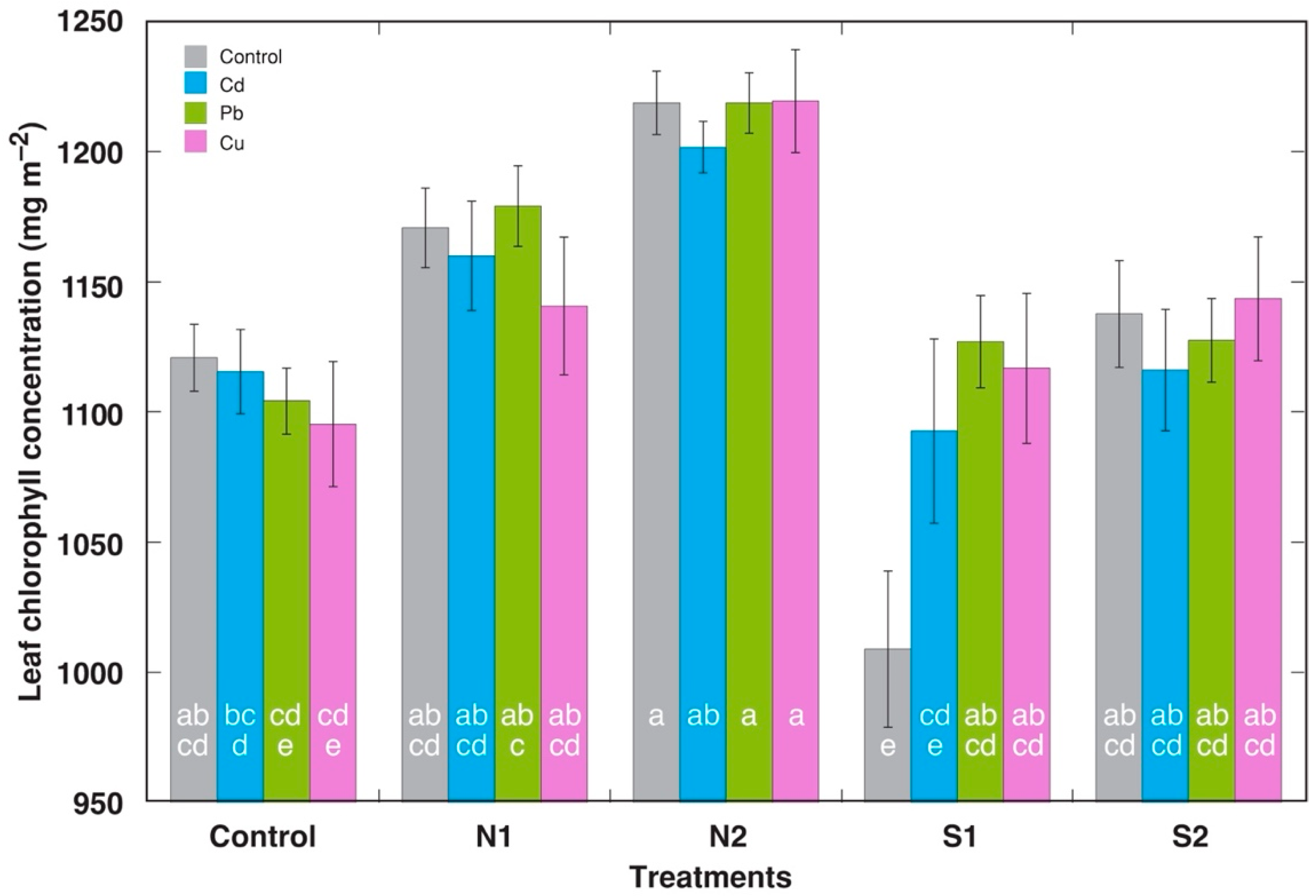
2.2. Experiment 2: Acute Treatment with Cd, Pb, and Cu on the Background of Different N Content and Salinity
3. Discussion
3.1. Metal Tolerance
3.2. Metal Accumulation
3.3. Possible Limitations and Perspectives
4. Materials and Methods
4.1. Plant Material
4.2. Experiment 1
4.3. Experiment 2
4.4. Measurement of Photosynthesis-Related Parameters
4.5. Termination of the Experiments
4.6. Metal Analysis
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asati, A.; Pichhode, M.; Nikhil, K. Effect of heavy metals on plants: An overview. Int. J. Appl. Innov. Eng. Manag. 2016, 5, 56–66. [Google Scholar]
- Ahamad, M.I.; Yao, Z.; Ren, L.; Zhang, C.; Li, T.; Lu, H.; Mehmood, M.; Rehman, A.; Muhammad, A.; Lu, S.; et al. Impact of heavy metals on aquatic life and human health: A case study of River Ravi Pakistan. Front. Mar. Sci. 2024, 11, 1374835. [Google Scholar] [CrossRef]
- Lv, Y.; Wang, M.; Tian, W.; Hui, H. The content and potential ecological risk assessment of heavy metals in coastal wetlands around the Bohai Sea. Plant Soil Environ. 2024, 70, 356–365. [Google Scholar] [CrossRef]
- Orson, R.A.; Simpson, R.J.; Good, R.E. A mechanism for the accumulation and retention of heavy metals in tidal freshwater marshes of the upper Delawere river. Estuar. Coast. Shelf Sci. 1992, 34, 171–186. [Google Scholar] [CrossRef]
- Williams, T.P.; Bubb, J.M.; Lester, J.N. The occurrence and distribution of trace metals in halophytes. Chemosphere 1994, 28, 1189–1199. [Google Scholar] [CrossRef]
- Doyle, M.O.; Otte, M.L. Organism-induced accumulation of iron, zinc and arsenic in wetland soils. Environ. Pollut. 1998, 96, 1–11. [Google Scholar] [CrossRef]
- Sruthi, P.; Shackira, A.M.; Puthur, J.T. Heavy metal detoxification mechanisms in halophytes: An overview. Wetl. Ecol. Manag. 2017, 25, 129–148. [Google Scholar] [CrossRef]
- Lutts, S.; Lefèvre, I. How can we take advantage of halophyte properties to cope with heavy metal toxicity in salt-affected areas? Ann. Bot. 2015, 115, 509–528. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Maggio, A. Functional biology of halophytes in the phytoremediation of heavy metal contaminated soils. Environ. Exp. Bot. 2015, 111, 135–146. [Google Scholar] [CrossRef]
- Caparrós, P.G.; Ozturk, M.; Gul, A.; Batool, T.S.; Pirasteh-Anosheh, H.; Unal, B.T.; Altay, V.; Toderich, K.N. Halophytes have potential as heavy metal phytoremediators: A comprehensive review. Environ. Exp. Bot. 2022, 193, 104666. [Google Scholar] [CrossRef]
- Tonelli, F.M.P.; Bhat, R.A.; Dar, G.H.; Hakeem, K.R. The history of phytoremediation. In Phytoremediation. Biotechnological Strategies for Promoting Invigorating Environs; Bhat, R.A., Tonelli, F.M.P., Dar, G.H., Hakeem, K.R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 1–18. [Google Scholar]
- Buscaroli, A. An overview of indexes to evaluate terrestrial plants for phytoremediation purposes (Review). Ecol. Indic. 2017, 82, 367–380. [Google Scholar] [CrossRef]
- Kafle, A.; Timilsina, A.; Gautam, A.; Adhikari, K.; Bhattarai, A.; Aryal, N. Phytoremediation: Mechanisms, plant selection and enhancement by natural and synthetic agents. Environ. Adv. 2022, 8, 100203. [Google Scholar] [CrossRef]
- Liang, Z.; Neményi, A.; Kovács, G.P.; Gyuricza, C. Incorporating functional traits into heavy metals phytoremediation: The future of field-based phytoremediation. Ecol. Indic. 2024, 166, 112262. [Google Scholar] [CrossRef]
- Mateos-Naranjo, E.; Pérez-Romero, J.A.; Redondo-Gómez, S.; Mesa-Marín, J.; Castellanos, E.M.; Davy, A.J. Salinity alleviates zinc toxicity in the saltmarsh zinc-accumulator Juncus acuttus. Ecotoxicol. Environ. Saf. 2018, 163, 478–485. [Google Scholar] [CrossRef]
- Nezhadasad-Aghbash, B.; Radjabian, T.; Hajiboland, R. Tolerance to Zn toxicity in the halophyte Lepidium latifolium L. and the effect of salt on Zn tolerance and accumulation. Acta Agric. Slov. 2023, 191, 1–17. [Google Scholar] [CrossRef]
- Zhou, M.; Engelmann, T.; Lutts, S. Salinity modifies heavy metals and arsenic absorption by the halophyte plant species Kosteletzkya pentacarpos and pollutant leaching from a polycontaminated substrate. Ecotoxicol. Environ. Saf. 2019, 182, 109460. [Google Scholar] [CrossRef]
- Kumari, V.V.; Banerjee, P.; Verma, V.C.; Sukumaran, S.; Chandran, M.A.S.; Gopinath, K.A.; Venkatesh, G.; Yadav, S.K.; Singh, V.K.; Awasthi, N.K. Plant nutrition: An effective way to alleviate abiotic stress in agricultural crops. Int. J. Mol. Sci. 2022, 23, 8519. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Nazir, F.; Maheshwari, C.; Chopra, P.; Chhillar, H.; Sreenivasulu, N. Mineral nutrients in plants under changing environments: A road to future food and nutrition security. Plant Genome 2023, 16, e20362. [Google Scholar] [CrossRef]
- Monsant, A.C.; Wang, Y.; Tang, C. Nitrate nutrition enhances zinc hyperaccumulation in Noccaea caerulescens (Prayon). Plant Soil 2010, 336, 391–404. [Google Scholar] [CrossRef]
- Hu, P.; Yin, Y.-G.; Ishikawa, S.; Suzui, N.; Kawachi, N.; Fujimaki, S.; Igura, M.; Yuan, C.; Huang, J.; Li, Z.; et al. Nitrate facilitates cadmium uptake, transport and accumulation in the hyperaccumulator Sedum plumbizincicola. Environ. Sci. Pollut. Res. 2013, 20, 6306–6316. [Google Scholar] [CrossRef]
- Wei, S.; Ji, D.; Twardowska, I.; Li, W.; Zhu, J. Effect of different nitrogenous nutrients on the cadmium hyperaccumulation efficiency of Rorippa globosa (Turcz.) Thell. Environ. Sci. Pollut. Res. 2015, 22, 1999–2007. [Google Scholar] [CrossRef] [PubMed]
- Kanso, A.; Azoury, S.; Benizri, E.; Kobaissi, A.; Echevarria, G.; Sirguey, C. Improvement of Ni phytoextaction by Alyssum murale and its rhizosphere microbial activities by applying nitrogen fertilizer. Ecol. Res. 2018, 33, 811–821. [Google Scholar] [CrossRef]
- Lin, Z.; Dou, C.; Li, Y.; Wang, H.; Niazi, N.K.; Zhang, S.; Liu, D.; Zhao, K.; Fu, W.; Li, Y.; et al. Nitrogen fertilizer enhances zinc and cadmium uptake by hyperaccumulator Sedum alfredii Hance. J. Soils Sedim. 2020, 20, 320–329. [Google Scholar] [CrossRef]
- Ievinsh, G.; Landorfa-Svalbe, Z.; Andersone-Ozola, U.; Karlsons, A.; Osvalde, A. Salinity and heavy metal tolerance, and phytoextraction potential of Ranunculus sceleratus plants from a sandy coastal beach. Life 2022, 12, 1959. [Google Scholar] [CrossRef]
- Landorfa-Svalbe, Z.; Andersone-Ozola, U.; Ievinsh, G. Type of anion largely determines salinity tolerance in four Rumex species. Plants 2023, 12, 92. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, C.; Hu, P.; Luo, W.; Wu, L.; Sale, P.; Tang, C. Influence of nitrogen form on the phytoextraction of cadmium by a newly discovered hyperaccumulator Carpobrotus rossii. Environ. Sci. Pollut. Res. 2016, 23, 1246–1253. [Google Scholar] [CrossRef]
- Ievinsh, G.; Ieviņa, S.; Andersone-Ozola, U.; Samsone, I. Leaf sodium, potassium and electrolyte accumulation capacity of plant species from salt-affected coastal habitats of the Baltic Sea: Towards a definition of Na hyperaccumulation. Flora 2021, 274, 151748. [Google Scholar] [CrossRef]
- Van der Sman, A.J.M.; Blom, C.W.P.; Barendse, G.W.M. Flooding resistance and shoot elongation in relation to developmental stage and environmental conditions in Rumex maritimus L. and Rumex palustris Sm. New Phytol. 1993, 125, 73–84. [Google Scholar] [CrossRef]
- Van der Sman, A.J.M.; van Tongeren, O.F.R.; Blom, C.W.P.M. Growth and reproduction of Rumex maritimus and Chenopodium rubrum under different waterlogging regimes. Acta Bot. Neerl. 1988, 37, 439–460. [Google Scholar] [CrossRef]
- Van der Sman, A.J.M.; Voesenek, L.A.C.J.; Blom, C.W.P.M.; Harren, F.J.M.; Reuss, J. The role of ethylene in shoot elongation with respect to survival and seed output of flooded Rumex maritimus L. plants. Funct. Ecol. 1991, 5, 304–313. [Google Scholar] [CrossRef]
- Samsone, I.; Ievinsh, G. Different plant species accumulate various concentration of Na+ in a sea-affected coastal wetland during a vegetation season. Environ. Exp. Biol. 2018, 16, 117–127. [Google Scholar]
- Ievinsh, G.; Dišlere, E.; Karlsons, A.; Osvalde, A.; Vikmane, M. Physiological responses of wetland species Rumex hydrolapathum to increased concentration of biogenous heavy metals Zn and Mn in substrate. Proc. Latv. Acad. Sci. B 2020, 7, 35–47. [Google Scholar] [CrossRef]
- Ieviņa, S.; Karlsons, A.; Osvalde, A.; Andersone-Ozola, U.; Ievinsh, G. Coastal wetland species Rumex hydrolapathum: Tolerance against flooding, salinity and heavy metals for its potential use in phytoremediation and environmental restoration technologies. Life 2023, 13, 1604. [Google Scholar] [CrossRef] [PubMed]
- Tyler, T.; Herbertsson, L.; Olofsson, J.; Olsson, P.A. Ecological indicator and trait values for Swedish vascular plants. Ecol. Indic. 2021, 120, 106923. [Google Scholar] [CrossRef]
- Samsone, I.; Ievinsh, G. Comparison of the effects of gradual and acute treatment with Mn on physiological responses of Rumex hydrolapathum plants. Stresses 2024, 4, 225–237. [Google Scholar] [CrossRef]
- Zhang, C.; Sale, P.W.G.; Tang, C. Cadmium uptake by Carpobrotus rossii (Haw.) Schwantes under different saline conditions. Environ. Sci. Pollut. Res. 2016, 23, 13480–13488. [Google Scholar] [CrossRef]
- Salama, F.M.; al-Huqail, A.A.; Ali, M.; Abeed, A.H.A. Cd phytoextraction potential in halophyte Salicornia fruticosa: Salinity impact. Plants 2022, 11, 2556. [Google Scholar] [CrossRef]
- Nawaz, I.; Iqbal, M.; Bliek, M.; Schat, H. Salt and heavy metal tolerance and expression levels of candidate tolerance genes among extremophile Cochlearia species with contrasting habitat preferences. Sci. Total Environ. 2017, 584–585, 731–741. [Google Scholar] [CrossRef]
- Purmale, L.; Jēkabsone, A.; Andersone-Ozola, U.; Ievinsh, G. Salinity tolerance, ion accumulation potential and osmotic adjustment in vitro and in planta of different Armeria maritima accessions from a dry coastal meadow. Plants 2022, 11, 2570. [Google Scholar] [CrossRef]
- Purmale, L.; Jēkabsone, A.; Andersone-Ozola, U.; Karlsons, A.; Osvalde, A.; Ievinsh, G. Comparison of in vitro and in planta heavy metal tolerance and accumulation potential of different Armeria maritima accessions from a dry coastal meadow. Plants 2022, 11, 2104. [Google Scholar] [CrossRef]
- Nikalje, G.C.; Suprasanna, P. Coping with metal toxicity—Cues from halophytes. Front. Plant Sci. 2018, 9, 777. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Guan, J.; Liang, Q.; Zhang, X.; Hu, H.; Zhang, J. Effects of cadmium stress on growth and physiological characteristics of Sassafras seedlings. Sci. Rep. 2021, 11, 9913. [Google Scholar] [CrossRef]
- Hu, Z.; Zhao, C.; Feng, Y.; Zhang, X.; Lu, Y.; Ying, R.; Yin, A.; Ji, W. Heavy metals can affect plant morphology and limit plant growth and photosynthesis processes. Agronomy 2023, 13, 2601. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]
- Ayeni, O.O.; Ndakidemi, P.A.; Snyman, R.G.; Odendaal, J.P. Chemical, biological and physiological indicators of metal pollution in wetlands. Sci. Res. Essays 2010, 5, 1938–1949. [Google Scholar]
- Singh, H.; Kumar, D.; Soni, V. Performance of chlorophyll a fluorescence parameters in Lemna minor under heavy metal stress induced by various concentrations of copper. Sci. Rep. 2022, 12, 10620. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker, A.J.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; van der Ent, A. A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef]
- Han, R.-M.; Lefèvre, I.; Ruan, C.-J.; Beukelaers, N.; Qin, P.; Lutts, S. Effects of salinity on the response of the wetland halophyte Kosteletzkya virginica (L.) Presl. to copper toxicity. Water Air Soil Pollut. 2012, 223, 1137–1150. [Google Scholar] [CrossRef]
- Manousaki, E.; Kalogerakis, N. Phytoextraction of Pb and Cd by the Mediteranean saltbush (Atriplex halimus L.): Metal uptake in relation to salinity. Environ Sci. Pollut Res. 2009, 16, 844–854. [Google Scholar] [CrossRef]
- Khanlarian, M.; Roshanfar, M.; Rashchi, F.; Motesharezadeh, B. Phyto-extraction of zinc, lead, nickel, and cadmium from zinc leach residue by a halophyte: Salicornia europaea. Ecol. Eng. 2020, 148, 105797. [Google Scholar] [CrossRef]
- Śliva-Cebula, M.; Kaszycki, P.; Aczmarczyk, A.; Nosek, M.; Lis-Krzyścin, A.; Miszalski, Z. The common ice plant (Mesembryanthemum crystallinum L.)—phytoremediation potential for cadmium and chromate-contaminated soils. Plants 2020, 9, 1230. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Lin, Y.; Yang, Y.; Shen, Q.; Huang, J.; Wang, S.; Zhu, X.; Li, Z. Tolerance and bioaccumulation of Cd and Cu in Sesuvium portulacastrum. Ecotoxicol. Environ. Saf. 2018, 147, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xiong, J.; Tao, L.; Cao, Z.; Tang, W.; Zhang, J.; Yu, X.; Fu, G.; Zhang, X.; Lu, Y. Regulatory mechanisms of nitrogen (N) on cadmium (Cd) uptake and accumulation in plants: A review. Sci. Total Environ. 2020, 708, 135186. [Google Scholar] [CrossRef]
- Eissa, M.A.; Roshdy, N.M.K. Nitrogen fertilization: Effect on Cd-phytoextraction by the halophytic plant quail bush [Atriplex lentiformis (Torr.) S. Wats]. S. Afr. J. Bot. 2018, 115, 126–131. [Google Scholar] [CrossRef]
- Cheng, W.; Wang, P.; Kopittke, P.M.; Wang, A.; Sale, P.W.G.; Tang, C. Cadmium accumulation is enhanced by ammonium compared to nitrate in two hyperaccumulators, without affecting speciation. J. Exp. Bot. 2016, 67, 5041–5050. [Google Scholar] [CrossRef]
- Jacobs, A.; Noret, N.; Van Baekel, A.; Liénard, A.; Colinet, G.; Drouet, T. Influence of edaphic conditions and nitrogen fertilizers on cadmium and zinc phytoextraction efficiency of Noccaea caerulescens. Sci. Total Environ. 2019, 665, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.A.; Jansen, B.; Kalbitz, K.; Faz, A.; Martínez-Martínez, S. Salinity increases mobility of heavy metals in soils. Chemosphere 2011, 85, 1318–1324. [Google Scholar] [CrossRef]
- Lefèvre, I.; Marchal, G.; Meerts, P.; Corréal, E.; Lutts, S. Chloride salinity reduces cadmium accumulation by the Mediterranean halophyte species Atriplex halimus L. Environ. Exp. Bot. 2009, 65, 142–152. [Google Scholar] [CrossRef]
- Manousaki, E.; Kokkali, F.; Kalogerakis, N. Influence of salinity on lead and cadmium accumulation by the salt cedar (Tamarix smyrnensis Bunge). J. Chem. Technol. Biotechnol. 2009, 84, 877–883. [Google Scholar] [CrossRef]
- Nosek, M.; Kaczmarczyk, A.; Jędrzejczyk, R.J.; Supel, P.; Kaszycki, P.; Miszalski, Z. Expression of genes involved in heavy metal trafficking in plants exposed to salinity stress and elevated Cd concentrations. Plants 2020, 9, 475. [Google Scholar] [CrossRef]
- Zurayk, R.A.; Khoury, N.F.; Talhouk, S.N.; Baalbaki, R.Z. Salinity-heavy metal interactions in four salt-tolerant plant species. J. Plant Nutr. 2001, 24, 1773–1786. [Google Scholar] [CrossRef]
- Zhou, M.-X.; Dailly, H.; Renard, M.-E.; Han, R.-M.; Lutts, S. NaCl impact on Kosteletzkya pentacarpos seedlings simultaneously exposed to cadmium and zinc toxicities. Environ. Sci. Pollut. Res. 2018, 25, 17444–17456. [Google Scholar] [CrossRef] [PubMed]
- Jarell, W.M.; Beverly, R.B. The dilution effect in plant nutrition studies. Adv. Agron. 1981, 34, 197–224. [Google Scholar]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy metal tolerance in plants: The role of transcriptomics, proteomics, metabolomics, and ionomics. Front. Plant Sci. 2016, 8, 1143. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Song, N. Salinity-induced alterations in plant growth, antioxidant enzyme activities, and lead transportation and accumulation in Suaeda salsa: Implications for phytoremediation. Ecotoxicology 2019, 28, 250–267. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, J.; Baran, A.; Bubak, A. Mobility, bioaccumulation in plants, and risk assessment of metals in soils. Sci. Total Environ. 2023, 882, 163574. [Google Scholar] [CrossRef]
- Wan, Y.; Liu, J.; Zhuang, Z.; Wang, Q.; Li, H. Heavy metals in agricultural soils: Sources, influencing factors, and remediation strategies. Toxics 2024, 12, 63. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Balgobind, A.; Pillay, B. Bioavailability of heavy metals in soil: Impact of microbial biodegradation of organic compounds and possible improvement strategies. Int. J. Mol. Sci. 2013, 14, 10197–10228. [Google Scholar] [CrossRef]
- Alves, A.R.A.; Yin, Q.; Oliveira, R.S.; Silva, E.F.; Novo, L.A.B. Plant growth-promoting bacteria in phytoremediation of metal-polluted soils: Current knowledge and future directions. Sci. Total Environ. 2022, 838, 156435. [Google Scholar] [CrossRef]
- Ievinsh, G.; Osvalde, A.; Karlsons, A.; Andersone-Ozola, U. Hylotelephium maximum from coastal drift lines is a promising Mn and Zn accumulator with a high tolerance to biogenous heavy metals. Stresses 2022, 2, 450–465. [Google Scholar] [CrossRef]
- Andersone-Ozola, U.; Jēkabsone, A.; Karlsons, A.; Osvalde, A.; Banaszczyk, L.; Samsone, I.; Ievinsh, G. Heavy metal tolerance and accumulation potential of a rare coastal species, Anthyllis vulneraria subsp. maritima. Stresses 2025, 5, 6. [Google Scholar] [CrossRef]
- Jēkabsone, A.; Kuļika, J.; Romanovs, M.; Andersone-Ozola, U.; Ievinsh, G. Salt tolerance and ion accumulation in several halophytic plant species depending on the type of anion. Int. J. Plant Biol. 2023, 14, 1131–1154. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M. Revisting JIP-test: An educative review on concepts, assumptions, approximations, definitions and terminology. Photosynthetica 2020, 58, 275–292. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Total Number of Leaves (n Plant−1) | Number of Dry Leaves (n Plant−1) | Total Leaf DM (g Plant−1) | Dry Leaf DM (g Plant−1) | Total Root DM (g Plant−1) |
|---|---|---|---|---|---|
| Control | 85.6 ± 6.4 ab | 11.4 ± 1.7 d | 10.4 ± 0.8 ab | 2.54 ± 0.28 abc | 14.5 ± 0.4 abcde |
| Cd (mg L−1) | |||||
| 20 | 89.6 ± 6.5 ab | 14.0 ± 1.6 d | 11.6 ± 0.8 ab | 1.00 ± 0.12 e | 16.0 ± 0.7 abc |
| 50 | 88.8 ± 4.2 ab | 18.2 ± 1.1 abcd | 11.8 ± 1.3 ab | 2.99 ± 0.48 ab | 15.1 ± 1.0 abcd |
| 100 | 82.6 ± 3.7 ab | 23.2 ± 2.8 ab | 9.6 ± 0.3 b | 1.74 ± 0.28 abc | 12.5 ± 0.7 def |
| Pb (mg L−1) | |||||
| 200 | 95.8 ± 3.4 ab | 16.4 ± 1.6 bcd | 11.4 ± 0.7 ab | 2.72 ± 0.18 abc | 16.3 ± 0.4 ab |
| 500 | 104.0 ± 3.8 a | 18.4 ± 1.1 abcd | 13.2 ± 0.5 a | 1.25 ± 0.09 de | 16.3 ± 0.5 ab |
| 1000 | 94.8 ± 6.2 ab | 14.6 ± 2.0 cd | 11.5 ± 0.8 ab | 2.31 ± 0.21 abcd | 13.6 ± 0.5 bcde |
| Cu (mg L−1) | |||||
| 100 | 93.8 ± 2.7 ab | 12.4 ± 0.8 d | 11.4 ± 0.5 ab | 0.92 ± 0.09 e | 14.5 ± 0.6 abcde |
| 200 | 80.4 ± 1.3 b | 12.2 ± 1.0 d | 10.7 ± 0.5 ab | 3.04 ± 0.18 ab | 11.7 ± 0.2 ef |
| 500 | 82.0 ± 3.9 b | 18.2 ± 1.6 abcd | 10.4 ± 0.3 ab | 1.25 ± 0.13 de | 11.3 ± 0.6 f |
| Mn (mg L−1) | |||||
| 200 | 91.2 ± 3.4 ab | 11.6 ± 0.7 d | 11.5 ± 0.7 ab | 3.26 ± 0.25 a | 16.8 ± 0.6 a |
| 500 | 85.2 ± 2.6 ab | 12.4 ± 0.7 d | 12.0 ± 1.0 ab | 0.92 ± 0.08 e | 13.1 ± 0.9 cdef |
| 1000 | 88.6 ± 4.3 ab | 15.6 ± 1.8 bcd | 12.6 ± 0.8 ab | 2.83 ± 0.33 abc | 12.5 ± 1.1 ef |
| Zn (mg L−1) | |||||
| 200 | 95.0 ± 1.5 ab | 17.8 ± 0.7 bcd | 13.0 ± 0.5 ab | 1.21 ± 0.05 de | 14.2 ± 0.5 abcde |
| 500 | 92.8 ± 3.3 ab | 22.2 ± 1.3 abc | 12.2 ± 0.8 ab | 2.73 ± 0.27 abc | 13.2 ± 0.6 bcdef |
| 1000 | 92.0 ± 6.1 ab | 26.0 ± 2.5 a | 10.7 ± 0.4 ab | 1.94 ± 0.18 bcde | 10.2 ± 0.6 f |
| Treatment | Concentration (mg L−1) | Performance Index Total (Units) |
|---|---|---|
| Control | – | 1.14 ± 0.09 b |
| Cd | 20 | 1.07 ± 0.16 b |
| 50 | 0.92 ± 0.08 b | |
| 100 | 1.28 ± 0.12 b | |
| Pb | 200 | 1.23 ± 0.03 b |
| 500 | 1.29 ± 0.08 b | |
| 1000 | 1.27 ± 0.08 b | |
| Cu | 100 | 1.21 ± 0.14 b |
| 200 | 1.13 ± 0.06 b | |
| 500 | 1.33 ± 0.10 b | |
| Mn | 200 | 1.18 ± 0.07 b |
| 500 | 1.20 ± 0.03 b | |
| 1000 | 1.36 ± 0.08 b | |
| Zn | 200 | 1.45 ± 0.10 b |
| 500 | 1.56 ± 0.15 ab | |
| 1000 | 1.99 ± 0.19 a |
| Treatment | Time After Full Treatment (Weeks) | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| Control | 0.92 ± 0.11 g | 2.35 ± 0.18 d | 1.70 ± 0.14 cde |
| Cd | 1.06 ± 0.14 g | 2.40 ± 0.20 d | 1.94 ± 0.19 cde |
| Pb | 0.97 ± 0.15 g | 2.77 ± 0.20 d | 1.34 ± 0.14 e |
| Cu | 1.55 ± 0.21 fg | 2.43 ± 0.33 d | 1.85 ± 0.28 cde |
| N1 | 2.81 ± 0.22 cdef | 3.92 ± 0.43 d | 1.50 ± 0.20 de |
| N1 Cd | 2.07 ± 0.15 fg | 5.20 ± 0.58 cd | 2.96 ± 0.30 c |
| N1 Pb | 4.03 ± 0.50 cde | 4.69 ± 0.75 cd | 1.81 ± 0.41 cde |
| N1 Cu | 4.02 ± 0.46 cde | 7.51 ± 1.79 bc | 2.87 ± 0.35 cd |
| N2 | 4.16 ± 0.39 cd | 11.31 ± 1.17 a | 2.95 ± 0.28 cd |
| N2 Cd | 6.47 ± 0.77 a | 9.15 ± 0.72 ab | 4.54 ± 0.42 ab |
| N2 Pb | 6.09 ± 0.55 ab | 9.89 ± 1.22 ab | 5.67 ± 0.54 aa |
| N2 Cu | 4.52 ± 0.61 bc | 10.78 ± 1.03 ab | 5.07 ± 0.51 ab |
| S1 | 2.95 ± 0.37 cdef | 2.46 ± 0.16 d | 1.30 ± 0.14 e |
| S1 Cd | 1.78 ± 0.20 fg | 3.44 ± 0.36 d | 1.70 ± 0.09 cde |
| S1 Pb | 2.42 ± 0.22 defg | 2.85 ± 0.31 d | 1.50 ± 0.21 cde |
| S1 Cu | 2.46 ± 0.20 defg | 3.38 ± 0.21 d | 4.68 ± 0.50 ab |
| S2 | 1.87 ± 0.31 fg | 3.29 ± 0.22 d | 2.07 ± 0.09 cde |
| S2 Cd | 2.35 ± 0.34 efg | 3.37 ± 0.34 d | 2.32 ± 0.34 cde |
| S2 Pb | 3.00 ± 0.34 cdef | 2.78 ± 0.21 d | 2.20 ± 0.13 cde |
| S2 Cu | 3.15 ± 0.35 cdef | 3.62 ± 0.69 d | 4.25 ± 0.49 b |
| Treatment (mg L−1 of Metal) | Salt | Total Amount of Salt (g per L of Soil) | Amount of Salt (g per L of Soil) | |
|---|---|---|---|---|
| 1st Treatment | 2nd Treatment | |||
| Control | – | – | – | – |
| Cd 20 | CdCl2 2.5H2O | 0.041 | 0.041 | – |
| Cd 50 | CdCl2 2.5H2O | 0.103 | 0.041 | 0.062 |
| Cd 100 | CdCl2 2.5H2O | 0.205 | 0.041 | 0.164 |
| Pb 200 | PbOAc 3H2O | 0.366 | 0.366 | – |
| Pb 500 | PbOAc 3H2O | 0.916 | 0.366 | 0.550 |
| Pb 1000 | PbOAc 3H2O | 1.832 | 0.366 | 1.466 |
| Cu 100 | CuSO4 5H2O | 0.390 | 0.390 | – |
| Cu 200 | CuSO4 5H2O | 0.780 | 0.390 | 0.390 |
| Cu 500 | CuSO4 5H2O | 1.950 | 0.390 | 1.560 |
| Mn 200 | MnSO4 H2O | 0.600 | 0.600 | – |
| Mn 500 | MnSO4 H2O | 1.500 | 0.600 | 0.900 |
| Mn 1000 | MnSO4 H2O | 3.000 | 0.600 | 2.400 |
| Zn 200 | ZnSO4 7H2O | 0.880 | 0.880 | – |
| Zn 500 | ZnSO4 7H2O | 2.200 | 0.880 | 1.320 |
| Zn 1000 | ZnSO4 7H2O | 4.400 | 0.880 | 3.520 |
| Treatment (mg L−1 of Metal) | Amount (g per L of Soil per Treatment) | ||
|---|---|---|---|
| 1st Treatment | 2nd Treatment | 3rd Treatment | |
| Control | – | – | – |
| Cd 100 | – | 0.205 g CdCl2 2.5H2O | – |
| Pb 1000 | – | 1.832 g Pb(OAc)2 3H2O | – |
| Cu 500 | – | 1.950 g CuSO4 5H2O | – |
| N1 (0.15 g L−1) | 1.26 g Ca(NO3)2 | – | – |
| N1 Cd 100 | 1.26 g Ca(NO3)2 | 0.205 g CdCl2 2.5H2O | – |
| N1 Pb 1000 | 1.26 g Ca(NO3)2 | 1.832 g Pb(OAc)2 3H2O | – |
| N1 Cu 500 | 1.26 g Ca(NO3)2 | 1.950 g CuSO4 5H2O | – |
| N2 (0.30 g L−1) | 1.26 g Ca(NO3)2 | – | 1.26 g Ca(NO3)2 |
| N2 Cd 100 | 1.26 g Ca(NO3)2 | 0.205 g CdCl2 2.5H2O | 1.26 g Ca(NO3)2 |
| N2 Pb 1000 | 1.26 g Ca(NO3)2 | 1.832 g Pb(OAc)2 3H2O | 1.26 g Ca(NO3)2 |
| N2 Cu 500 | 1.26 g Ca(NO3)2 | 1.26 g Ca(NO3)2 | |
| S1 (50 mM) | 2.90 g NaCl | – | – |
| S1 Cd 100 | 2.90 g NaCl | 0.205 g CdCl2 2.5H2O | – |
| S1 Pb 1000 | 2.90 g NaCl | 1.832 g Pb(OAc)2 3H2O | – |
| S1 Cu 500 | 2.90 g NaCl | 1.950 g CuSO4 5H2O | – |
| S2 (100 mM) | 2.90 g NaCl | – | 2.90 g NaCl |
| S2 Cd 100 | 2.90 g NaCl | 0.205 g CdCl2 2.5H2O | 2.90 g NaCl |
| S2 Pb 1000 | 2.90 g NaCl | 1.832 g Pb(OAc)2 3H2O | 2.90 g NaCl |
| S2 Cu 500 | 2.90 g NaCl | 1.950 g CuSO4 5H2O | 2.90 g NaCl |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neiceniece, L.; Jēkabsone, A.; Andersone-Ozola, U.; Banaszczyk, L.; Karlsons, A.; Osvalde, A.; Ievinsh, G. Effect of Salinity and Nitrogen on Heavy Metal Tolerance and Accumulation Potential in Rumex maritimus. Stresses 2025, 5, 29. https://doi.org/10.3390/stresses5020029
Neiceniece L, Jēkabsone A, Andersone-Ozola U, Banaszczyk L, Karlsons A, Osvalde A, Ievinsh G. Effect of Salinity and Nitrogen on Heavy Metal Tolerance and Accumulation Potential in Rumex maritimus. Stresses. 2025; 5(2):29. https://doi.org/10.3390/stresses5020029
Chicago/Turabian StyleNeiceniece, Lāsma, Astra Jēkabsone, Una Andersone-Ozola, Lidia Banaszczyk, Andis Karlsons, Anita Osvalde, and Gederts Ievinsh. 2025. "Effect of Salinity and Nitrogen on Heavy Metal Tolerance and Accumulation Potential in Rumex maritimus" Stresses 5, no. 2: 29. https://doi.org/10.3390/stresses5020029
APA StyleNeiceniece, L., Jēkabsone, A., Andersone-Ozola, U., Banaszczyk, L., Karlsons, A., Osvalde, A., & Ievinsh, G. (2025). Effect of Salinity and Nitrogen on Heavy Metal Tolerance and Accumulation Potential in Rumex maritimus. Stresses, 5(2), 29. https://doi.org/10.3390/stresses5020029