

Lonely Beetles Lose Weight: Absence of Conspecific Sounds Negatively Impacts Body Mass in Larval and Adult Passalus Beetles


Abstract
1. Introduction
2. Results
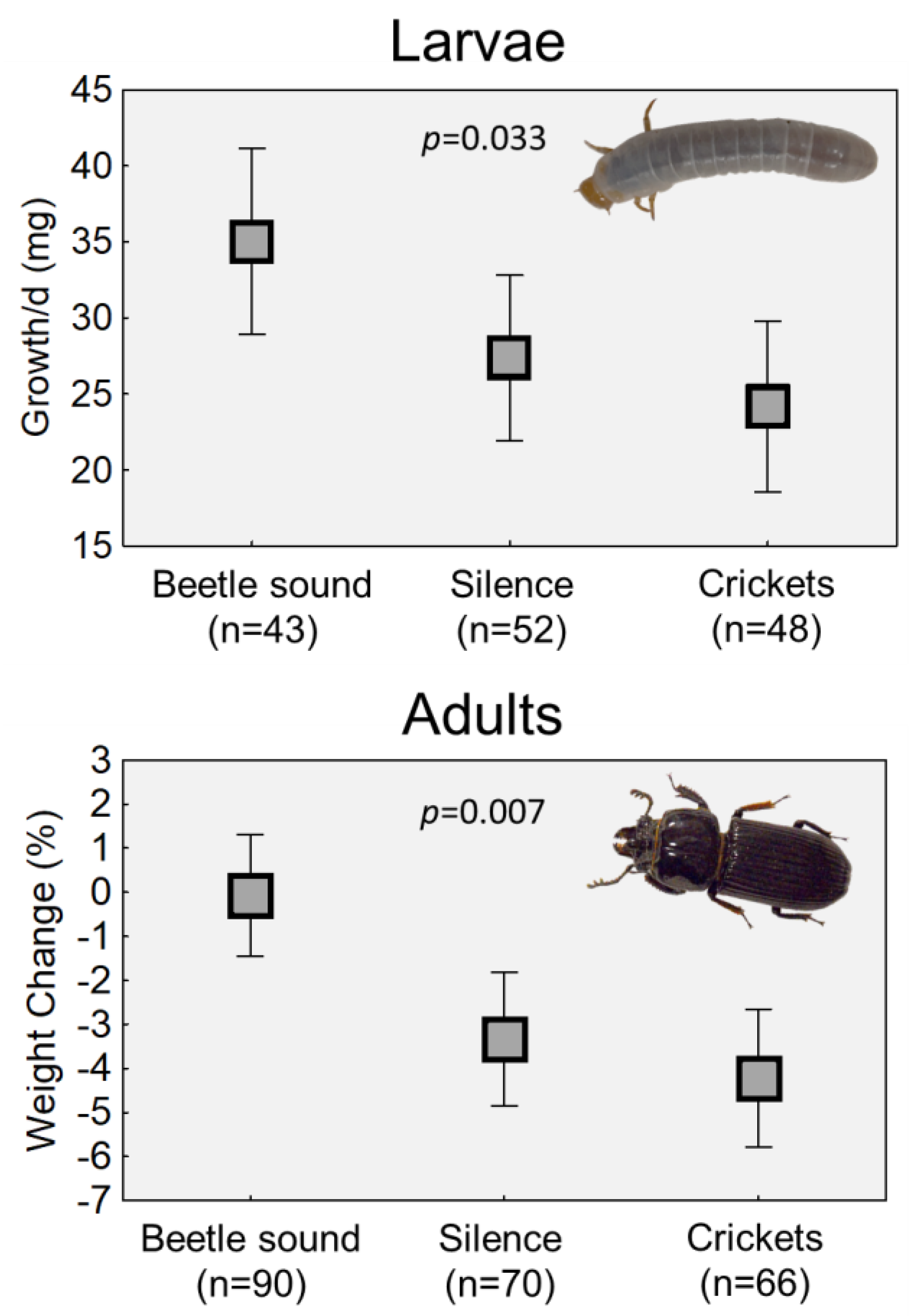
2.1. Larvae Experiment
2.2. Adult Experiments
3. Discussion
4. Methods
4.1. Larvae Experiment
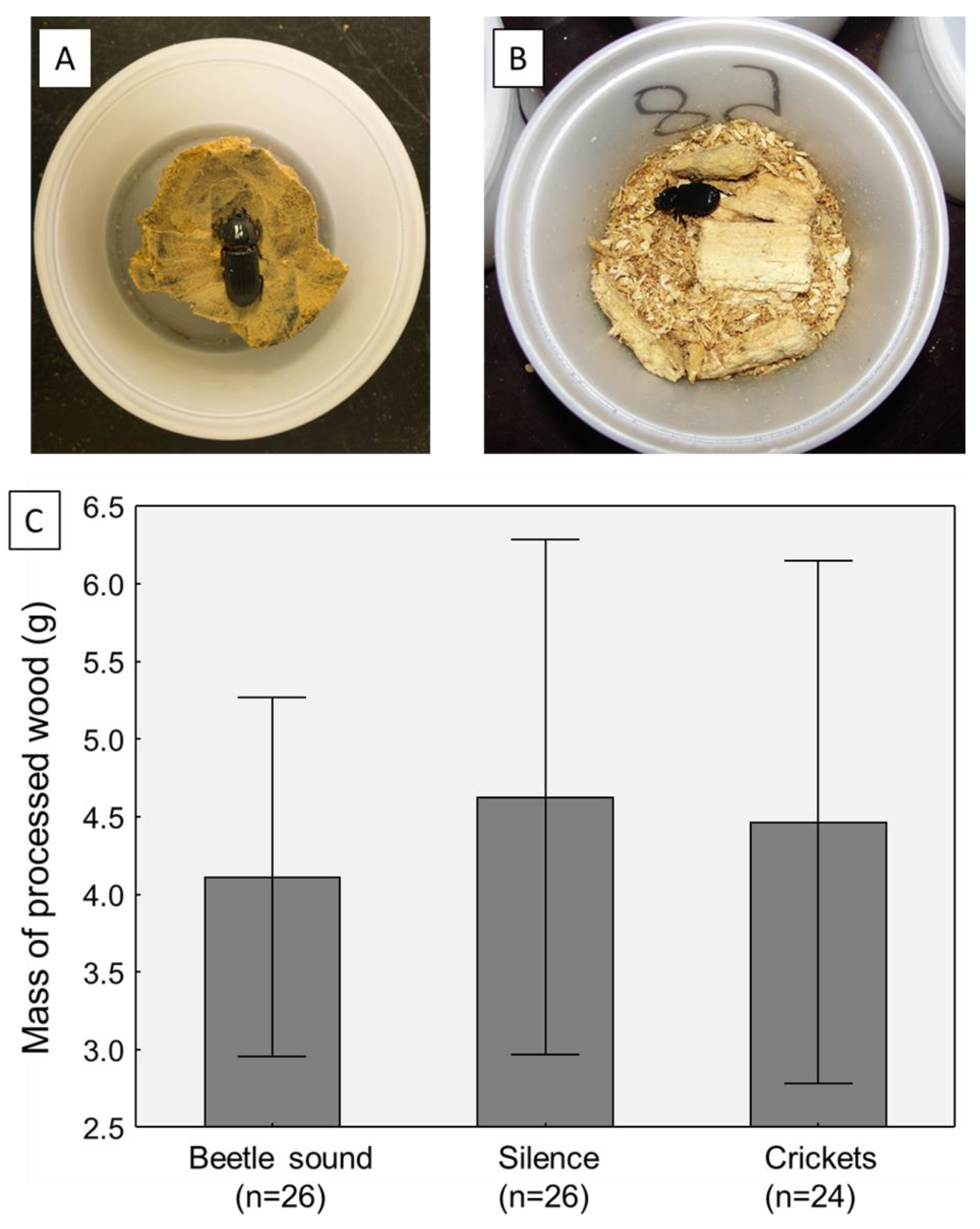
4.2. Adult Experiments
4.3. Data Analyses
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ware, H.E.; McClure, C.J.W.; Carlisle, J.D.; Barber, J.R. A phantom road experiment reveals traffic noise is an invisible source of habitat degradation. Proc. Natl. Acad. Sci. USA 2015, 112, 12105–12109. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.K.; Schroeder, H.; Yeager, I.; Pearce, J. Effects of simulated highway noise on heart rates of larval monarch butterflies, Danaus plexippus: Implications for roadside habitat suitability. Biol. Lett. 2018, 14, 20180018. [Google Scholar] [CrossRef] [PubMed]
- Erbe, C.; Marley, S.A.; Schoeman, R.P.; Smith, J.N.; Trigg, L.E.; Embling, C.B. The Effects of Ship Noise on Marine Mammals—A Review. Front. Mar. Sci. 2019, 6, 606. [Google Scholar] [CrossRef]
- Riley, J.R.; Greggers, U.; Smith, A.D.; Reynolds, D.R.; Menzel, R. The flight paths of honeybees recruited by the waggle dance. Nature 2005, 435, 205–207. [Google Scholar] [CrossRef] [PubMed]
- Richard, F.J.; Hunt, J.H. Intracolony chemical communication in social insects. Insectes Sociaux 2013, 60, 275–291. [Google Scholar] [CrossRef]
- Hunt, J.H.; Richard, F.J. Intracolony vibroacoustic communication in social insects. Insectes Sociaux 2013, 60, 403–417. [Google Scholar] [CrossRef]
- Manser, M.B.; Jansen, D.; Graw, B.; Hollén, L.I.; Bousquet, C.A.H.; Furrer, R.D.; le Roux, A. Vocal Complexity in Meerkats and Other Mongoose Species. In Advances in the Study of Behavior; Naguib, M., Barrett, L., Brockmann, H.J., Healy, S., Mitani, J.C., Roper, T.J., Simmons, L.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 46, pp. 281–310. [Google Scholar]
- Jones, B.; Zapetis, M.; Samuelson, M.M.; Ridgway, S. Sounds produced by bottlenose dolphins (Tursiops): A review of the defining characteristics and acoustic criteria of the dolphin vocal repertoire. Bioacoustics 2020, 29, 399–440. [Google Scholar] [CrossRef]
- Mumtaz, F.; Khan, M.I.; Zubair, M.; Dehpour, A.R. Neurobiology and consequences of social isolation stress in animal model-A comprehensive review. Biomed. Pharmacother. 2018, 105, 1205–1222. [Google Scholar] [CrossRef]
- Fontanella, F.M.; Whitaker, M.; Smallwood, J.; Enderle, K.; Shinall-Graham, A.; Adams, J. Disentangling disjunctus: An integrated analysis of the patent leather beetle, Odontotaenius disjunctus complex (Passalidae: Coleoptera) with the description of two new species. Syst. Biodivers. 2024, 22, 2413503. [Google Scholar] [CrossRef]
- Pearse, A.S.; Patterson, M.; Rankin, J.S.; Wharton, G.W. The ecology of Passalus cornutus Fabricius, a beetle which lives in rotting logs. Ecol. Monogr. 1936, 6, 456–490. [Google Scholar] [CrossRef]
- Dillard, J.R. High rates of extra-pair paternity in a socially monogamous beetle with biparental care. Ecol. Entomol. 2017, 42, 1–10. [Google Scholar] [CrossRef]
- Reyes-Castillo, P.; Jarman, M. Some notes on larval stridulation in neotropical Passalidae (Coleoptera: Lamellicornia). Coleopt. Bull. 1980, 34, 263–270. [Google Scholar]
- Schuster, J.C. Acoustical signals of passalid beetles: Complex repertoires. Fla. Entomol. 1983, 66, 486–496. [Google Scholar] [CrossRef]
- Schuster, J.C.; Schuster, L.B. Social behavior in passalid beetles (Coleoptera, Passalidae): Cooperative brood care. Fla. Entomol. 1985, 68, 266–272. [Google Scholar] [CrossRef]
- LeFeuvre, J.; Davis, A.K. Effects of a naturally-occurring nematode parasite on lifting strength and captivity-related body mass patterns in horned passalus beetles, Odontotaenius disjunctus. Coleopt. Bull. 2015, 69, 744–750. [Google Scholar] [CrossRef]
- Kuti, D.; Winkler, Z.; Horváth, K.; Juhász, B.; Szilvásy-Szabó, A.; Fekete, C.; Ferenczi, S.; Kovács, K.J. The metabolic stress response: Adaptation to acute- repeated- and chronic challenges in mice. iScience 2022, 25, 104693. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, D., Jr.; Willoughby, A.; Davis, A.K. Fighting while parasitized: Can nematode infections affect the outcome of staged combat in beetles? PLoS ONE 2015, 10, e0121614. [Google Scholar] [CrossRef]
- Davis, A.K.; Coogler, B.; Johnson, I. The heartrate reaction to acute stress in horned passalus beetles (Odontotaenius disjunctus) is negatively affected by a naturally-occurring nematode parasite. Insects 2017, 8, 110. [Google Scholar] [CrossRef]
- Davis, A.K.; Calderon, L.; Lefeuvre, J.; Sims, S.; Pearce, J.; Prouty, C. Healing while parasitized: Impact of a naturally-occurring nematode during energy-intensive wound-healing in a beetle. Physiol. Entomol. 2020, 45, 72–80. [Google Scholar] [CrossRef]
- Koto, A.; Mersch, D.; Hollis, B.; Keller, L. Social isolation causes mortality by disrupting energy homeostasis in ants. Behav. Ecol. Sociobiol. 2015, 69, 583–591. [Google Scholar] [CrossRef]
- Schausberger, P.; Gratzer, M.; Strodl, M.A. Early social isolation impairs development, mate choice and grouping behaviour of predatory mites. Anim. Behav. 2017, 127, 15–21. [Google Scholar] [CrossRef]
- Stevenson, P.A.; Rillich, J. Isolation Associated Aggression—A Consequence of Recovery from Defeat in a Territorial Animal. PLoS ONE 2013, 8, e74965. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.K.; Prouty, C. The sicker the better: Nematode-infected passalus beetles provide enhanced ecosystem services. Biol. Lett. 2019, 15, 4. [Google Scholar] [CrossRef] [PubMed]
- Dillard, J.R.; Maigret, T.A. Delayed dispersal and prolonged brood care in a family-living beetle. J. Evol. Biol. 2017, 30, 2230–2243. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.K.; Vasquez, D.; LeFeuvre, J.; Sims, S.; Craft, M.; Vizurraga, A. Parasite manipulation of its host’s physiological reaction to acute stress: Experimental results from a natural beetle-nematode system. Physiol. Biochem. Zool. 2017, 90, 273–280. [Google Scholar] [CrossRef]
- Ebot-Ojong, F.; Jurado, E.; Davis, A.K. Direct measurement of fight or flight behavior in a beetle reveals individual variation and the influence of parasitism. PLoS ONE 2019, 14, e0216387. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.; Davis, A.K. Effect of a parasitic nematode, Chondronema passali Leidy (Incertae sedis), on the size and strength of the horned passalus, Odontotaenius disjunctus Illiger (Coleoptera: Passalidae). Coleopt. Bull. 2013, 67, 179–185. [Google Scholar] [CrossRef]
- Davis, A.K.; Hurd, C.; Brandon, C.; Vasquez, D. Walking while parasitized: Effects of a naturally-occurring nematode on locomotor activity of horned passalus beetles. J. Insect Behav. 2021, 34, 89–95. [Google Scholar] [CrossRef]
- Gray, I.E. Observations on the life history of the horned passalus. Am. Midl. Nat. 1946, 35, 728–746. [Google Scholar] [CrossRef]
- Davis, A.K.; Ladd, R.R.E.; Smith, F.; Shattuck, A. Sex-specific effects of a parasite on stress-induced freezing behavior in a natural beetle-nematode system. PLoS ONE 2023, 18, e0281149. [Google Scholar] [CrossRef] [PubMed]
- Adamo, S.A.; Baker, J.L. Conserved features of chronic stress across phyla: The effects of long-term stress on behavior and the concentration of the neurohormone octopamine in the cricket, Gryllus texensis. Horm. Behav. 2011, 60, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Calderon, L.; Davis, A.K. Observations of Steinernema nematode and Tachinid fly parasites in horned passalus beetles, Odontotaenius disjunctus, from Georgia, U.S.A. Comp. Parasitol. 2016, 83, 265–268. [Google Scholar] [CrossRef]
- Reinert, J.A. Parasites associated with Popilius disjunctus in South Carolina (Coleoptera: Passalidae). Fla. Entomol. 1973, 56, 273–276. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| A. Beetle grubs | ||||||
| Predictor | df | SS | MS | F | p | |
| Treatment | 2 | 2598.86 | 1299.43 | 3.50 | 0.0330 | |
| Error | 131 | 48,632.85 | 371.24 | |||
| B. Adult beetles | ||||||
| Predictor | Effect | df | SS | MS | F | p |
| Replicate | Random | 2 | 685.08 | 342.54 | 8.25 | 0.0004 |
| Treatment | Fixed | 2 | 744.49 | 372.24 | 8.96 | 0.0002 |
| Error | 220 | 9138.28 | 41.54 | |||
| C. Adult beetles | ||||||
| Predictor | Effect | df | SS | MS | F | p |
| Replicate | Random | 1 | 277.15 | 277.15 | 5.62 | 0.0190 |
| Treatment | Fixed | 2 | 501.73 | 250.86 | 5.09 | 0.0073 |
| Sex | Fixed | 1 | 292.27 | 292.27 | 5.93 | 0.0161 |
| Parasitized | Fixed | 1 | 20.96 | 20.96 | 0.43 | 0.5154 |
| Error | 146 | 7197.89 | 49.30 | |||
| Total | 8546.71 | |||||
| Experiment | Beetle Sound | No Sound | Cricket Sound | Total |
|---|---|---|---|---|
| Beetle grubs (June 2021) | 43 | 52 | 48 | 143 |
| Adult beetles (June 2021) | 43 | 22 | 23 | 88 |
| Adult beetles (February 2022) | 21 | 22 | 19 | 62 |
| Adult beetles (March 2023) | 26 | 26 | 24 | 76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davis, A.K. Lonely Beetles Lose Weight: Absence of Conspecific Sounds Negatively Impacts Body Mass in Larval and Adult Passalus Beetles. Stresses 2025, 5, 11. https://doi.org/10.3390/stresses5010011
Davis AK. Lonely Beetles Lose Weight: Absence of Conspecific Sounds Negatively Impacts Body Mass in Larval and Adult Passalus Beetles. Stresses. 2025; 5(1):11. https://doi.org/10.3390/stresses5010011
Chicago/Turabian StyleDavis, Andrew K. 2025. "Lonely Beetles Lose Weight: Absence of Conspecific Sounds Negatively Impacts Body Mass in Larval and Adult Passalus Beetles" Stresses 5, no. 1: 11. https://doi.org/10.3390/stresses5010011
APA StyleDavis, A. K. (2025). Lonely Beetles Lose Weight: Absence of Conspecific Sounds Negatively Impacts Body Mass in Larval and Adult Passalus Beetles. Stresses, 5(1), 11. https://doi.org/10.3390/stresses5010011