
Molecular Survey of Anaplasmataceae Agents and Coxiellaceae in Non-Hematophagous Bats and Associated Ectoparasites from Brazil

 , , , ,
, , , , {kind=link}
Abstract
1. Introduction
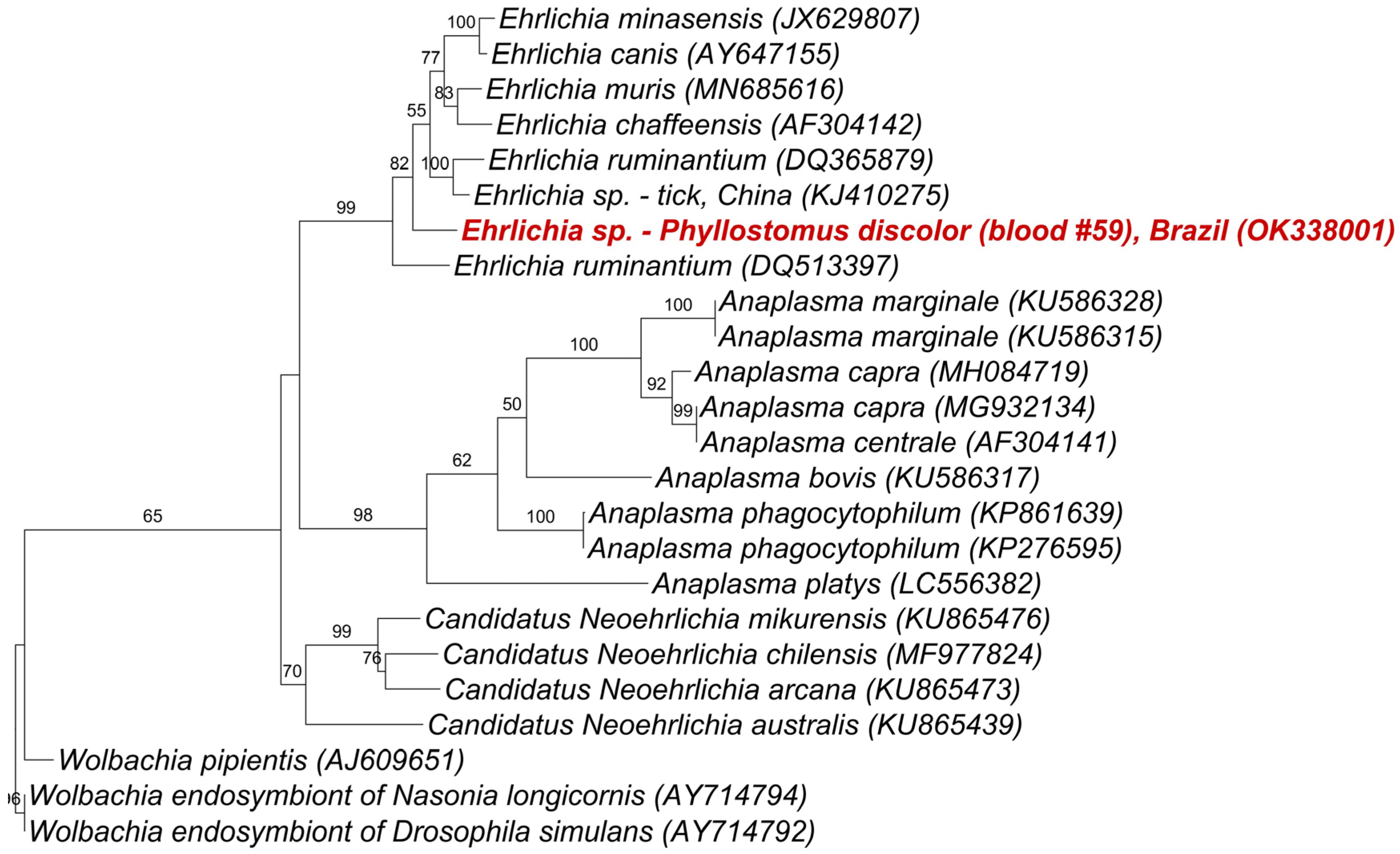
2. Results
3. Discussion
4. Material and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schipper, J.; Chanson, J.S.; Chiozza, F.; Cox, N.A.; Hoffmann, M.; Katariya, V.; Lamoreux, J.; Rodrigues, A.S.; Stuart, S.N.; Temple, H.J.; et al. The status of the world’s land and marine mammals: Diversity, threat, and knowledge. Science 2008, 322, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Garbino, G.S.T.; Gregorin, R.; Lima, I.P.; Loureiro, L.; Moras, L.M.; Moratelli, R.; Nogueira, M.R.; Pavan, A.C.; Tavares, V.C.; do Nascimento, M.C.; et al. Updated Checklist of Brazilian Bats: Versão 2020. Comitê da Lista de Morcegos do Brasil—CLMB. Sociedade Brasileira para o Estudo de Quirópteros (Sbeq). Available online: https://www.sbeq.net/lista-de-especies (accessed on 6 July 2021).
- Mühldorfer, K. Bats and Bacterial Pathogens: A Review. Zoonoses Public Health 2013, 60, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51 Pt 6, 2145–2165. [Google Scholar] [CrossRef] [PubMed]
- Cabezas-Cruz, A.; Zweygarth, E.; Vancová, M.; Broniszewska, M.; Grubhoffer, L.; Passos, L.M.F.; Ribeiro, M.F.B.; Alberdi, P.; de la Fuente, J. Ehrlichia minasensis sp. nov., isolated from the tick Rhipicephalus microplus. Int. J. Syst. Evol. Microbiol. 2016, 66, 1426–1430. [Google Scholar] [CrossRef] [PubMed]
- André, M.R. Diversity of Anaplasma and Ehrlichia/Neoehrlichia Agents in Terrestrial Wild Carnivores Worldwide: Implications for Human and Domestic Animal Health and Wildlife Conservation. Front. Vet. Sci. 2018, 5, 293. [Google Scholar] [CrossRef]
- Bakken, J.S.; Dumler, J.S. Clinical diagnosis and treatment of human granulocytotropic anaplasmosis. Ann. N. Y. Acad. Sci. 2006, 1078, 236–247. [Google Scholar] [CrossRef]
- Ismail, N.; McBride, J.W. Tick-Borne Emerging Infections: Ehrlichiosis and Anaplasmosis. Clin. Lab. Med. 2017, 37, 317–340. [Google Scholar] [CrossRef]
- Rar, V.; Tkachev, S.; Tikunova, N. Genetic diversity of Anaplasma bacteria: Twenty years later. Infect. Genet. Evol. 2021, 91, 104833. [Google Scholar] [CrossRef]
- Silaghi, C.; Santos, A.S.; Gomes, J.; Christova, I.; Matei, I.A.; Walder, G.; Domingos, A.; Bell-Sakyi, L.; Sprong, H.; von Loewenich, F.D.; et al. Guidelines for the Direct Detection of Anaplasma spp. in Diagnosis and Epidemiological Studies. Vector Borne Zoonotic Dis. 2017, 17, 12–22. [Google Scholar] [CrossRef]
- Moraes-Filho, J.; Krawczak, F.S.; Costa, F.B.; Soares, J.F.; Labruna, M.B. Comparative Evaluation of the Vector Competence of Four South American Populations of the Rhipicephalus sanguineus Group for the Bacterium Ehrlichia canis, the Agent of Canine Monocytic Ehrlichiosis. PLoS ONE 2015, 10, e0139386. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.; Bloch, K.C.; McBride, J.W. Human ehrlichiosis and anaplasmosis. Clin. Lab. Med. 2010, 30, 261–292. [Google Scholar] [CrossRef]
- Rochlin, I.; Toledo, A. Emerging tick-borne pathogens of public health importance: A mini-review. J. Med. Microbiol. 2020, 69, 781–791. [Google Scholar] [CrossRef]
- Kawahara, M.; Rikihisa, Y.; Isogai, E.; Takahashi, M.; Misumi, H.; Suto, C.; Shibata, S.; Zhang, C.; Tsuji, M. Ultrastructure and phylogenetic analysis of ‘Candidatus Neoehrlichia mikurensis’ in the family Anaplasmataceae, isolated from wild rats and found in Ixodes ovatus ticks. Int. J. Syst. Evol. Microbiol. 2004, 54 Pt 5, 1837–1843. [Google Scholar] [CrossRef]
- Yabsley, M.J.; Murphy, S.M.; Luttrell, M.P.; Wilcox, B.R.; Howerth, E.W.; Munderloh, U.G. Characterization of ‘Candidatus Neoehrlichia lotoris’ (family Anaplasmataceae) from raccoons (Procyon lotor). Int. J. Syst. Evol. Microbiol. 2008, 58 Pt 12, 2794–2798. [Google Scholar] [CrossRef]
- Gofton, A.W.; Doggett, S.; Ratchford, A.; Ryan, U.; Irwin, P. Phylogenetic characterisation of two novel Anaplasmataceae from Australian Ixodes holocyclus ticks: ‘Candidatus Neoehrlichia australis’ and ‘Candidatus Neoehrlichia arcana’. Int. J. Syst. Evol. Microbiol. 2016, 66, 4256–4261. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Monti, G.; Otth, C.; Sepúlveda, P.; Bittencourt, P.; Nachum-Biala, Y.; Gutiérrez, R.; Harrus, S. “Candidatus Neoehrlichia chilensis” sp. nov.: Molecular detection and characterization of a novel Anaplasmataceae in wild rodents from Valdivia, southern Chile. Transbound. Emerg. Dis. 2018, 65, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Brouqui, P.; Sanogo, Y.O.; Caruso, G.; Merola, F.; Raoult, D. Candidatus Ehrlichia walkerii: A new Ehrlichia detected in Ixodes ricinus tick collected from asymptomatic humans in Northern Italy. Ann. N. Y. Acad. Sci. 2003, 990, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Gofton, A.W.; Waudby, H.P.; Petit, S.; Greay, T.L.; Ryan, U.M.; Irwin, P.J. Detection and phylogenetic characterisation of novel Anaplasma and Ehrlichia species in Amblyomma triguttatum subsp. from four allopatric populations in Australia. Ticks Tick-Borne Dis. 2017, 8, 749–756. [Google Scholar] [CrossRef]
- Rar, V.A.; Pukhovskaya, N.M.; Ryabchikova, E.I.; Vysochina, N.P.; Bakhmetyeva, S.V.; Zdanovskaia, N.I.; Ivanov, L.I.; Tikunova, N.V. Molecular-genetic and ultrastructural characteristics of ‘Candidatus Ehrlichia khabarensis’, a new member of the Ehrlichia genus. Ticks Tick-Borne Dis. 2015, 6, 658–667. [Google Scholar] [CrossRef]
- Dahmani, M.; Davoust, B.; Tahir, D.; Raoult, D.; Fenollar, F.; Mediannikov, O. Molecular investigation and phylogeny of Anaplasmataceae species infecting domestic animals and ticks in Corsica, France. Parasites Vectors 2017, 10, 302. [Google Scholar] [CrossRef]
- Gofton, A.W.; Loh, S.M.; Barbosa, A.D.; Paparini, A.; Gillett, A.; Macgregor, J.; Oskam, C.L.; Ryan, U.M.; Irwin, P.J. A novel Ehrlichia species in blood and Ixodes ornithorhynchi ticks from platypuses (Ornithorhynchus anatinus) in Queensland and Tasmania, Australia. Ticks Tick-Borne Dis. 2018, 9, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Leal, S.; Clemes, Y.S.; Lopes, M.G.; Acosta, I.; Serpa, M.; Mayorga, L.; Gennari, S.M.; González-Acuña, D.; Labruna, M.B. Novel Ehrlichia sp. detected in Magellanic penguins (Sphenicus magellanicus) and in the seabird tick Ixodes uriae from Magdalena Island, southern Chile. Ticks Tick-Borne Dis. 2019, 10, 101256. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Diao, X.N.; Zhao, G.Y.; Chen, M.H.; Xiong, Y.; Shi, M.; Fu, W.M.; Guo, Y.J.; Pan, B.; Chen, X.P.; et al. Extensive diversity of Rickettsiales bacteria in two species of ticks from China and the evolution of the Rickettsiales. BMC Evol. Biol. 2014, 14, 167. [Google Scholar] [CrossRef] [PubMed]
- Dahmani, M.; Davoust, B.; Rousseau, F.; Raoult, D.; Fenollar, F.; Mediannikov, O. Natural Anaplasmataceae infection in Rhipicephalus bursa ticks collected from sheep in the French Basque Country. Ticks Tick-Borne Dis. 2017, 8, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.P.; Huang, B.; Zhao, Q.; Xu, G.; Liu, B.; Wang, Y.H.; Zhou, E.M. Human-pathogenic Anaplasma spp., and Rickettsia spp. in animals in Xi’an, China. PLoS Negl. Trop. Dis. 2018, 12, e0006916. [Google Scholar] [CrossRef] [PubMed]
- Vanstreels, R.; Yabsley, M.J.; Parsons, N.J.; Swanepoel, L.; Pistorius, P.A. A novel candidate species of Anaplasma that infects avian erythrocytes. Parasites Vectors 2018, 11, 525. [Google Scholar] [CrossRef]
- Dahmani, M.; Davoust, B.; Sambou, M.; Bassene, H.; Scandola, P.; Ameur, T.; Raoult, D.; Fenollar, F.; Mediannikov, O. Molecular investigation and phylogeny of species of the Anaplasmataceae infecting animals and ticks in Senegal. Parasites Vectors 2019, 12, 495. [Google Scholar] [CrossRef]
- Calchi, A.C.; Vultão, J.G.; Alves, M.H.; Yogui, D.R.; Desbiez, A.; De Santi, M.; Santana, M.S.; da Silva, T.; Werther, K.; Teixeira, M.; et al. Ehrlichia spp. and Anaplasma spp. in Xenarthra mammals from Brazil, with evidence of novel ‘Candidatus Anaplasma spp.’. Sci. Rep. 2020, 10, 12615. [Google Scholar] [CrossRef]
- Greiman, S.E.; Tkach, M.; Vaughan, J.A.; Tkach, V.V. Laboratory maintenance of the bacterial endosymbiont, Neorickettsia sp., through the life cycle of a digenean, Plagiorchis elegans. Exp. Parasitol. 2015, 157, 78–83. [Google Scholar] [CrossRef]
- Teymournejad, O.; Lin, M.; Bekebrede, H.; Kamr, A.; Toribio, R.E.; Arroyo, L.G.; Baird, J.D.; Rikihisa, Y. Isolation and Molecular Analysis of a Novel Neorickettsia Species That Causes Potomac Horse Fever. mBio 2020, 11, e03429-19. [Google Scholar] [CrossRef]
- Paulino, P.G.; Almosny, N.; Oliveira, R.; Viscardi, V.; Müller, A.; Guimarães, A.; Baldani, C.; da Silva, C.; Peckle, M.; Massard, C.; et al. Detection of Neorickettsia risticii, the agent of Potomac horse fever, in horses from Rio de Janeiro, Brazil. Sci. Rep. 2020, 10, 7208. [Google Scholar] [CrossRef]
- Dutra, F.; Schuch, L.D.F.; Delucchi, E.; Curcio, B.R.; Coimbra, H.S.; Raffi, M.B.; Dellagostin, O.; Riet-Correa, F. Equine monocytic Ehrlichiosis (Potomac Horse Fever) in horses in Uruguay and southern Brazil. J. Vet. Diagn. Investig. 2001, 13, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, H.S.; Fernandes, C.G.; Soares, M.P.; Meireles, M.C.A.; Radamés, R.; Schuch, L.F.D. Equine monocytic Ehrlichiosis in Rio Grande do Sul: Clinical, pathological and epidemiological aspects. Pesqui. Veterinária Bras. 2006, 26, 97–101. [Google Scholar] [CrossRef]
- Afonso, E.; Goydadin, A.C. Molecular detection of Anaplasma phagocytophilum DNA in the lesser horseshoe bat (Rhinolophus hipposideros) guano. Epidemiol. Infect. 2018, 146, 1253–1258. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Szoke, K.; Meli, M.L.; Sándor, A.D.; Görföl, T.; Estók, P.; Wang, Y.; Tu, V.T.; Kováts, D.; Boldogh, S.A.; et al. Molecular detection of vector-borne bacteria in bat ticks (Acari: Ixodidae, Argasidae) from eight countries of the Old and New Worlds 06 Biological Sciences 0604 Genetics. Parasites Vectors 2019, 12, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zabashta, M.V.; Orlova, M.V.; Pichurina, N.L.; Khametova, A.P.; Romanova, L.V.; Borodina, T.N.; Zabashta, A.V. Participation of Bats (Chiroptera, Mammalia) and Their Ectoparasites in Circulation of Pathogens of Natural Focal Infections in the South of Russia. Entmol. Rev. 2019, 99, 513–521. [Google Scholar] [CrossRef]
- Socolovschi, C.; Kernif, T.; Raoult, D.; Parola, P. Borrelia, Rickettsia, and Ehrlichia species in bat ticks, France, 2010. Emerg. Infect. Dis. 2012, 18, 1966–1975. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.K.; Beck, J.; Orlova, M.V.; Daly, J.L.; Pippin, K.; Revan, F.; Loftis, A.D. Ecology of Bats, Their Ectoparasites, and Associated Pathogens on Saint Kitts Island. J. Med. Entomol. 2016, 53, 1218–1225. [Google Scholar] [CrossRef]
- Lv, J.; Fernández de Marco, M.; Goharriz, H.; Phipps, L.P.; McElhinney, L.M.; Hernández-Triana, L.M.; Wu, S.; Lin, X.; Fooks, A.R.; Johnson, N. Detection of tick-borne bacteria and babesia with zoonotic potential in Argas (Carios) vespertilionis (Latreille, 1802) ticks from British bats. Sci. Rep. 2018, 8, 1865. [Google Scholar] [CrossRef] [PubMed]
- Pusterla, N.; Johnson, E.M.; Chae, J.S.; Madigan, J.E. Digenetic trematodes, Acanthatrium sp. and Lecithodendrium sp., as vectors of Neorickettsia risticii, the agent of Potomac horse fever. J. Helminthol. 2003, 77, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Gibson, K.E.; Rikihisa, Y.; Zhang, C.; Martin, C. Neorickettsia risticii is vertically transmitted in the trematode Acanthatrium oregonense and Horizontally transmitted to bats. Environ. Microbiol. 2005, 2, 203–212. [Google Scholar] [CrossRef]
- Cicuttin, G.L.; Boeri, E.J.; Beltrán, F.J.; Dohmen, F.E.G. Molecular detection of Neorickettsia risticii in Brazilian free-tailed bats (Tadarida brasiliensis) from Buenos Aires, Argentina. Pesqui. Veterinária Bras. 2013, 33, 648–650. [Google Scholar] [CrossRef]
- Cicuttin, G.L.; De Salvo, M.N.; La Rosa, I.; Dohmen, F.E.G. Neorickettsia risticii, Rickettsia sp. and Bartonella sp. in Tadarida brasiliensis bats from Buenos Aires, Argentina. Comp. Immunol. Microbiol. Infect. Dis. 2017, 52, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Greiman, S.E.; Vaughan, J.A.; Elmahy, R.; Adisakwattana, P.; Van Ha, N.; Fayton, T.J.; Khalil, A.I.; Tkach, V.V. Real-time PCR detection and phylogenetic relationships of Neorickettsia spp. in digeneans from Egypt, Philippines, Thailand, Vietnam and the United States. Parasitol. Int. 2017, 66, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Szőke, K.; Estók, P.; Krawczyk, A.; Haarsma, A.J.; Kováts, D.; Boldogh, S.A.; Morandini, P.; Szekeres, S.; Takács, N.; et al. Assessing bat droppings and predatory bird pellets for vector-borne bacteria: Molecular evidence of bat-associated Neorickettsia sp. in Europe. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2018, 111, 1707–1717. [Google Scholar] [CrossRef] [PubMed]
- Maurin, M.; Raoult, D. Q fever. Clin. Microbiol. Rev. 1999, 12, 518–553. [Google Scholar] [CrossRef] [PubMed]
- Eldin, C.; Mélenotte, C.; Mediannikov, O.; Ghigo, E.; Million, M.; Edouard, S.; Mege, J.L.; Maurin, M.; Raoult, D. From Q Fever to Coxiella burnetii Infection: A Paradigm Change. Clin. Microbiol. Rev. 2017, 30, 115–190. [Google Scholar] [CrossRef]
- Damasceno, I.; Guerra, R.C. Coxiella burnetii e a febre Q no Brasil, uma questão de saúde pública [Coxiella burnetii and Q fever in Brazil: A public health issue]. Cienc. Saude Coletiva 2018, 23, 4231–4239. [Google Scholar] [CrossRef]
- Angelakis, E.; Raoult, D. Q Fever. Vet. Microbiol. 2010, 140, 297–309. [Google Scholar] [CrossRef]
- Leulmi, H.; Aouadi, A.; Bitam, I.; Bessas, A.; Benakhla, A.; Raoult, D.; Parola, P. Detection of Bartonella tamiae, Coxiella burnetii and rickettsiae in arthropods and tissues from wild and domestic animals in northeastern Algeria. Parasites Vectors 2016, 9, 27. [Google Scholar] [CrossRef]
- Dietrich, M.; Kearney, T.; Seamark, E.C.; Markotter, W. The excreted microbiota of bats: Evidence of niche specialisation based on multiple body habitats. FEMS Microbiol. Lett. 2017, 364, fnw284. [Google Scholar] [CrossRef]
- Ferreira, M.S.; Guterres, A.; Rozental, T.; Novaes, R.L.M.; Vilar, E.M.; De Oliveira, R.C.; Fernandes, J.; Forneas, D.; Junior, A.A.; Brandão, M.L.; et al. Coxiella and Bartonella spp. In bats (Chiroptera) captured in the Brazilian Atlantic Forest biome. BMC Vet. Res. 2018, 14, 279. [Google Scholar] [CrossRef] [PubMed]
- Szubert-Kruszyńska, A.; Stańczak, J.; Cieniuch, S.; Podsiadły, E.; Postawa, T.; Michalik, J. Bartonella and Rickettsia Infections in Haematophagous Spinturnix myoti Mites (Acari: Mesostigmata) and their Bat Host, Myotis myotis (Yangochiroptera: Vespertilionidae), from Poland. Microb. Ecol. 2019, 77, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Mongruel, A.C.B.; Benevenute, J.L.; Ikeda, P.; André, M.R.; Machado, R.Z.; Carrasco, A.O.T.; Seki, M.C. Detection of Anaplasma sp. Phylogenetically related to A. phagocytophilum in a free-living bird in Brazil. Rev. Bras. De Parasitol. Vet. 2017, 26, 505–510. [Google Scholar] [CrossRef]
- Allsopp, B.A. Heartwater--Ehrlichia ruminantium infection. Rev. Sci. Et Tech. 2015, 34, 557–568. [Google Scholar] [CrossRef]
- Widmer, C.E.; Azevedo, F.C.; Almeida, A.P.; Ferreira, F.; Labruna, M.B. Tick-borne bacteria in free-living jaguars (Panthera onca) in Pantanal, Brazil. Vector Borne Zoonotic Dis. 2011, 11, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.P.; Souza, T.D.; Marcili, A.; Labruna, M.B. Novel Ehrlichia and Hepatozoon agents infecting the crab-eating fox (Cerdocyon thous) in southeastern Brazil. J. Med. Entomol. 2013, 50, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Cruz, A.C.; Zweygarth, E.; Ribeiro, M.F.; da Silveira, J.A.; de la Fuente, J.; Grubhoffer, L.; Valdés, J.J.; Passos, L.M. New species of Ehrlichia isolated from Rhipicephalus (Boophilus) microplus shows an ortholog of the E. canis major immunogenic glycoprotein gp36 with a new sequence of tandem repeats. Parasites Vectors 2012, 5, 291. [Google Scholar] [CrossRef]
- Aguiar, D.M.; Araujo, J.P., Jr.; Nakazato, L.; Bard, E.; Cabezas-Cruz, A. Complete Genome Sequence of an Ehrlichia minasensis Strain Isolated from Cattle. Microbiol. Resour. Announc. 2019, 8, e00161-19. [Google Scholar] [CrossRef]
- Aguiar, D.M.; Ziliani, T.F.; Zhang, X.; Melo, A.L.; Braga, I.A.; Witter, R.; Freitas, L.C.; Rondelli, A.L.; Luis, M.A.; Sorte, E.C.; et al. A novel Ehrlichia genotype strain distinguished by the TRP36 gene naturally infects cattle in Brazil and causes clinical manifestations associated with ehrlichiosis. Ticks Tick-Borne Dis. 2014, 5, 537–544. [Google Scholar] [CrossRef]
- Benevenute, J.L.; Dumler, J.S.; Ogrzewalska, M.; Roque, A.; Mello, V.; de Sousa, K.; Gonçalves, L.R.; D’Andrea, P.S.; de Sampaio Lemos, E.R.; Machado, R.Z.; et al. Assessment of a quantitative 5’ nuclease real-time polymerase chain reaction using groEL gene for Ehrlichia and Anaplasma species in rodents in Brazil. Ticks Tick-Borne Dis. 2017, 8, 646–656. [Google Scholar] [CrossRef]
- De Sousa, K.; Calchi, A.C.; Herrera, H.M.; Dumler, J.S.; Barros-Battesti, D.M.; Machado, R.Z.; André, M.R. Anaplasmataceae agents among wild mammals and ectoparasites in Brazil. Epidemiol. Infect. 2017, 145, 3424–3437. [Google Scholar] [CrossRef]
- Holland, C.; Weiss, E.; Burgdorfer, W.; Cole, A.I.; Kakoma, I. Ehrlichia risticii sp. nov.: Etiological Agent of Equine Monocytic Ehrlichiosis (Synonym, Potomac Horse Fever). Int. J. Syst. Evol. Microbiol. 1985, 35, 524–526. [Google Scholar] [CrossRef][Green Version]
- Barlough, J.E.; Reubel, G.H.; Madigan, J.E.; Vredevoe, L.K.; Miller, P.E.; Rikihisa, Y. Detection of Ehrlichia risticii, the agent of Potomac horse fever, in freshwater stream snails (Pleuroceridae: Juga spp.) from northern California. Appl. Environ. Microbiol. 1998, 4, 2888–2893. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.A.; Tkach, V.V.; Greiman, S.E. Chapter 3—Neorickettsial Endosymbionts of the Digenea: Diversity, Transmission and Distribution. Adv. Parasitol. 2012, 79, 253–297. [Google Scholar] [CrossRef] [PubMed]
- Baldridge, G.D.; Scoles, G.A.; Burkhardt, N.Y.; Schloeder, B.; Kurtti, T.J.; Munderloh, U.G. Transovarial transmission of Francisella-like endosymbionts and Anaplasma phagocytophilum variants in Dermacentor albipictus (Acari: Ixodidae). J. Med. Entomol. 2009, 46, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Braga, M.; Gonçalves, L.R.; Silva, T.; Costa, F.B.; Pereira, J.G.; Santos, L.; Carvalho Neta, A.V.; Arruda, R.; Mesquita, E.; Chaves, D.P.; et al. Occurrence of Bartonella genotypes in bats and associated Streblidae flies from Maranhão state, northeastern Brazil. Rev. Bras. De Parasitol. Vet. 2020, 29, e014420. [Google Scholar] [CrossRef]
- Müller, A.; Sepúlveda, P.; Di Cataldo, S.; Cevidanes, A.; Lisón, F.; Millán, J. Molecular investigation of zoonotic intracellular bacteria in Chilean bats. Comp. Immunol. Microbiol. Infect. Dis. 2020, 73, 101541. [Google Scholar] [CrossRef]
- Costa, P.S.; Brigatte, M.E.; Greco, D.B. Questing one Brazilian query: Reporting 16 cases of Q fever from Minas Gerais, Brazil. Rev. Do Inst. De Med. Trop. De Sao Paulo 2006, 48, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Brenner, A.E.; Muñoz-Leal, S.; Sachan, M.; Labruna, M.B.; Raghavan, R. Coxiella burnetii and Related Tick Endosymbionts Evolved from Pathogenic Ancestors. Genome Biol. Evol. 2021, 13, evab108. [Google Scholar] [CrossRef]
- Lemos, E.R.; Rozental, T.; Mares-Guia, M.A.; Almeida, D.N.; Moreira, N.; Silva, R.G.; Barreira, J.D.; Lamas, C.C.; Favacho, A.R.; Damasco, P.V. Q fever as a cause of fever of unknown origin and thrombocytosis: First molecular evidence of Coxiella burnetii in Brazil. Vector Borne Zoonotic Dis. 2011, 11, 85–87. [Google Scholar] [CrossRef]
- Lamas, C.C.; Fournier, P.E.; Zappa, M.; Brandão, T.J.; Januário-da-Silva, C.A.; Correia, M.G.; Barbosa, G.I.; Golebiovski, W.F.; Weksler, C.; Lepidi, H.; et al. Diagnosis of blood culture-negative endocarditis and clinical comparison between blood culture-negative and blood culture-positive cases. Infection 2016, 44, 459–466. [Google Scholar] [CrossRef]
- Ikeda, P.; Menezes, T.R.; Torres, J.M.; de Oliveira, C.E.; Lourenço, E.C.; Herrera, H.M.; Machado, R.Z.; André, M.R. First molecular detection of piroplasmids in non-hematophagous bats from Brazil, with evidence of putative novel species. Parasitol. Res. 2021, 120, 301–310. [Google Scholar] [CrossRef]
- Birkenheuer, A.J.; Levy, M.G.; Breitschwerdt, E.B. Development and evaluation of a seminested PCR for detection and differentiation of Babesia gibsoni (Asian genotype) and B. canis DNA in canine blood samples. J. Clin. Microbiol. 2003, 41, 4172–4177. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Massung, R.F.; Slater, K.; Owens, J.H.; Nicholson, W.L.; Mather, T.N.; Solberg, V.B.; Olson, J.G. Nested PCR assay for detection of granulocytic ehrlichiae. J. Clin. Microbiol. 1998, 36, 1090–1095. [Google Scholar] [CrossRef]
- Doyle, C.K.; Labruna, M.B.; Breitschwerdt, E.B.; Tang, Y.W.; Corstvet, R.E.; Hegarty, B.C.; Bloch, K.C.; Li, P.; Walker, D.H.; McBride, J.W. Detection of medically important Ehrlichia by quantitative multicolor TaqMan real-time polymerase chain reaction of the dsb gene. J. Mol. Diagn. JMD 2005, 7, 504–510. [Google Scholar] [CrossRef]
- Reller, M.E.; Dumler, J.S. Development and Clinical Validation of a Multiplex Real-Time Quantitative PCR Assay for Human Infection by Anaplasma phagocytophilum and Ehrlichia chaffeensis. Trop. Med. Infect. Dis. 2018, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Drazenovich, N.; Foley, J.; Brown, R.N. Use of real-time quantitative PCR targeting the msp2 protein gene to identify cryptic Anaplasma phagocytophilum infections in wildlife and domestic animals. Vector Borne Zoonotic Dis. 2006, 6, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Sumner, J.W.; Nicholson, W.L.; Massung, R.F. PCR amplification and comparison of nucleotide sequences from the groESL heat shock operon of Ehrlichia species. J. Clin. Microbiol. 1997, 35, 2087–2092. [Google Scholar] [CrossRef] [PubMed]
- O’Nion, V.L.; Montilla, H.J.; Qurollo, B.A.; Maggi, R.G.; Hegarty, B.C.; Tornquist, S.J.; Breitschwerdt, E.B. Potentially novel Ehrlichia species in horses, Nicaragua. Emerg. Infect. Dis. 2015, 21, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Inayoshi, M.; Naitou, H.; Kawamori, F.; Masuzawa, T.; Ohashi, N. Characterization of Ehrlichia species from Ixodes ovatus ticks at the foot of Mt. Fuji, Japan. Microbiol. Immunol. 2004, 48, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Rejmanek, D.; Bradburd, G.; Foley, J. Molecular characterization reveals distinct genospecies of Anaplasma phagocytophilum from diverse North American hosts. J. Med. Microbiol. 2012, 61 Pt 2, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.S.; Kim, E.H.; Kim, M.S.; Kim, M.J.; Cho, Y.H.; Park, B.K. Prevalence and sequence analyses of Neorickettsia Risticii. Ann. N. Y. Acad. Sci. 2003, 990, 248–256. [Google Scholar] [CrossRef]
- Kanter, M.; Mott, J.; Ohashi, N.; Fried, B.; Reed, S.; Lin, Y.C.; Rikihisa, Y. Analysis of 16S rRNA and 51-kilodalton antigen gene and transmission in mice of Ehrlichia risticii in virgulate trematodes from Elimia livescens snails in Ohio. J. Clin. Microbiol. 2000, 38, 3349–3358. [Google Scholar] [CrossRef]
- Gibson, K.E.; Pastenkos, G.; Moesta, S.; Rikihisa, Y. Neorickettsia risticii surface-exposed proteins: Proteomics identification, recognition by naturally-infected horses, and strain variations. Vet. Res. 2011, 42, 71. [Google Scholar] [CrossRef]
- Klee, S.R.; Tyczka, J.; Ellerbrok, H.; Franz, T.; Linke, S.; Baljer, G.; Appel, B. Highly sensitive real-time PCR for specific detection and quantification of Coxiella burnetii. BMC Microbiol. 2006, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Ostell, J.; Pruitt, K.D.; Sayers, E.W. GenBank. Nucleic Acids Res. 2018, 46, D41–D47. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikeda, P.; Torres, J.M.; Placa, A.J.V.; Mello, V.V.C.d.; Lourenço, E.C.; Herrera, H.M.; Oliveira, C.E.d.; Hemsley, C.; Titball, R.W.; Machado, R.Z.; et al. Molecular Survey of Anaplasmataceae Agents and Coxiellaceae in Non-Hematophagous Bats and Associated Ectoparasites from Brazil. Parasitologia 2021, 1, 197-209. https://doi.org/10.3390/parasitologia1040021
Ikeda P, Torres JM, Placa AJV, Mello VVCd, Lourenço EC, Herrera HM, Oliveira CEd, Hemsley C, Titball RW, Machado RZ, et al. Molecular Survey of Anaplasmataceae Agents and Coxiellaceae in Non-Hematophagous Bats and Associated Ectoparasites from Brazil. Parasitologia. 2021; 1(4):197-209. https://doi.org/10.3390/parasitologia1040021
Chicago/Turabian StyleIkeda, Priscila, Jaire Marinho Torres, Ana Julia Vidal Placa, Victória Valente Califre de Mello, Elizabete Captivo Lourenço, Heitor Miraglia Herrera, Carina Elisei de Oliveira, Claudia Hemsley, Richard W. Titball, Rosangela Zacarias Machado, and et al. 2021. "Molecular Survey of Anaplasmataceae Agents and Coxiellaceae in Non-Hematophagous Bats and Associated Ectoparasites from Brazil" Parasitologia 1, no. 4: 197-209. https://doi.org/10.3390/parasitologia1040021
APA StyleIkeda, P., Torres, J. M., Placa, A. J. V., Mello, V. V. C. d., Lourenço, E. C., Herrera, H. M., Oliveira, C. E. d., Hemsley, C., Titball, R. W., Machado, R. Z., & André, M. R. (2021). Molecular Survey of Anaplasmataceae Agents and Coxiellaceae in Non-Hematophagous Bats and Associated Ectoparasites from Brazil. Parasitologia, 1(4), 197-209. https://doi.org/10.3390/parasitologia1040021