Abstract
We describe a new species of terrestrial-breeding frog of the genus Pristimantis from the Peruvian Yungas ecoregion of central Peru, Junin Department. The description is based on the observation of morphological features, color patterns of fourteen specimens, bioacoustic traits of the advertisement calls, and a phylogenetic analysis using a sequence fragment of 16S rRNA. The new species is mainly characterized by having a rounded or truncate snout in dorsal view, presence of dorsolateral folds, tympanic membrane and annulus distinct, absence of flash marks on the hidden surfaces of flanks and hindlimbs, and a small size (maximum SVL 15.6 mm in males and 19.3 mm in females). We provide diagnostic traits to differentiate the new species from phylogenetically close relatives, as well as morphologically similar and sympatric species of Pristimantis.
1. Introduction
The neotropical frogs of the genus of Pristimantis form the most species rich amphibian taxon currently ranked as a genus. It includes 592 species named to date that occur from lower Central America (Honduras, El Salvador, Nicaragua, Costa Rica and Panama) to the Choco and Tumbesian regions (through the Pacific Slope from Colombia and Ecuador to northwestern Peru), along the Tropical Andes (from Southern Venezuela to Bolivia), throughout the Guyana Shield and Amazon Basin, the northern portion of the Mata Atlántica, and the islands of Trinidad and Tobago [1,2,3]. Two species occur in the Lesser Antilles in Grenada and Saint Vincent, the closest to mainland South America [2]. The greatest diversity of the genus occurs in the Tropical Andes, where some species inhabit elevations from sea level to up to about 4000 m [1,2].
Peru is the third country with the highest species richness of Pristimantis (148 species), after Colombia and Ecuador [1]. However, most of the country has not been well surveyed in herpetological terms [2,4,5], thus reviewing museum specimens or exploring remote places often results in the discovery of new species. For example, the herpetological exploration of the Cordillera Oriental of central Peru (Departments of Huánuco, Pasco and Junín) is continuously increasing the number of Pristimantis species. The addition of seven species of Pristimantis in the Andes of central Peru since Blair Hedges collected in the cloud forest of Pasco in 1987 suggests that this region harbors a high diversity of strabomantid frogs [6,7]. Subsequently, several collections carried out in the montane forests near Huánuco, Oxapampa, Pampa Hermosa, Huasawasi, Paucartambo, Yanachaga Chémillen National Park, Pui Pui Protected Forest and the remote Cordillera del Sira led to the discovery of 34 new species of Pristimantis [8,9,10,11,12,13,14,15,16,17,18]. These trends suggest that the number of species of Pristimantis in the Peruvian Yungas of central Peru is far from reaching saturation.
Similar to species in other genera of terrestrial-breeding frogs in the Andes, many species of Pristimantis are endemic to small montane areas and valleys, e.g., [19,20,21], where habitat destruction driven by agriculture, livestock and mining activities make them vulnerable to extinction [22]. Thus, the formal description of new species is the first step to make species visible and useful to the scientific community and to the resource managers who aim to protect and conserve biodiversity [23].
With the aim of increasing our knowledge about the diversity of Pristimantis, herein we describe and name a new species discovered during a herpetological expedition carried out in the Junín Department, central Peru, in December 2019. We provide diagnostic characters to differentiate the new species from similar congeners in Peru and we describe its call. In addition, we also describe the advertisement call of the morphologically similar congeneric and co-occurring species P. albertus Duellman and Hedges, 2007.
2. Materials and Methods
2.1. Field Research and Ethics
Specimens were collected during a herpetological survey conducted in December 2019 along the Satipo Road between Satipo and Huancayo, Junín Department. The specimens of frogs described herein were collected by hand using the complete species inventory technique [24] during slow nocturnal (20:00 to 2:00) walks along trails and in the forest. The geographic location data were taken with a GPS (Garmin, WGS84). We euthanized voucher specimens with 20% benzocaine gel, fixed them in 10% formalin for 24 h, and stored them permanently in 70% ethanol.
Collected specimens were housed in the herpetological collection of the Centro de Ornitología y Biodiversidad (CORBIDI) in Lima, Peru. The hepetological survey and collection of specimens were carried out under permit RDG 093-2020-MINAGRI-SERFOR.
2.2. Morphology
The format for the descriptions follows Lynch and Duellman [25]. The terminology and definition of diagnostic characters follows Duellman and Lehr [2]. Sex was determined by the direct observation of dimorphic characters as the presence of vocal slits or vocal sac and by dissection via gonadal inspection. Measurements were taken with digital calipers and rounded to the nearest 0.1 mm. We measured SVL (snout–vent length), TL (tibia length), FL (foot length, distance from proximal margin of inner metatarsal tubercle to tip of Toe IV), HL (head length, obliquely from angle of jaw to tip of snout), HW (head width, at level of angle of jaw), ED (eye diameter, distance between the anterior and posterior borders of the visible eye), IOD (interorbital distance, distance between the medial edge of the orbits), EW (upper eyelid width, length of the visible eye along the outer edge of eyelid), IND (internarial distance, distance between the inner edges of nares), and EN (eye–nostril distance, distance between the anterior corner of orbit and the posterior margin of nares); following Duellman and Lehr [2]. Fingers and toes are numbered preaxially to postaxially from I to IV and I to V, respectively, according with Duellman and Lehr [2]. Comparative lengths of Toes III and V were determined when both were adpressed against Toe IV; lengths of Fingers I and II were compared when adpressed against each other following Lynch and Duellman [25]. Specimens examined for comparisons are listed in Appendix A.
2.3. Molecular Analyses
We examined the phylogenetic relationships of the new species within Pristimantis based on the sequence fragment of the non-coding 16S rRNA mitochondrial gene. To compare our newly generated sequence with congeners, we considered sequences of the 12 species of Pristimantis most similar to the new species on the basis of BLAST results across Genbank sequences, as well as the outgroup spieces P. buccinator Rodriguez, 1994, and P. sinschi Moravec, Lehr, and Kodejs, 2020, and the new sequences of P. clarae (sequence for the holotype CORBIDI 21664, Genbank Accession Code OQ556784), for a total of 20 terminals. Extraction, amplification, and sequencing of DNA followed standard protocols [10]. We used the 16Sar (forward) primer (5′-3′ sequence: CGCCTGTTTATCAAAAACAT) and the 16Sbr (reverse) primer (5′-3′ sequence: CCGGTCTGAACTCAGATCACGT) with the following thermocycling conditions (PCR) on a Proflex PCR system (Applied Biosystems, Carlsbad, CA, USA): one cycle of 96 °C/3 min; 35 cycles of 95 °C/30 s, 55 °C/45 s, 72 °C/1.5 min; one cycle 72 °C/7 min. We purified PCR products with Exosap-IT (Affymetrix, Santa Clara, CA, USA) and obtained sequences from MCLAB (San Francisco, CA, USA). We used Geneious, version 11.1.5 (Biomatters, http://www.geneious.com/ accessed on 3 April 2023) to align sequences with the MAFFT v7.017 alignment program [26], and trimmed sequences to a length of 542 bp without removing any variable fragment. We calculated p-distances using MEGA v. 7 [27]. We inferred phylogenetic relationships with a Maximum Likelihood (ML) inference approach on IQ-TREE [28] using the online server (http://iqtree.cibiv.univie.ac.at/accessed on 4 April 2023), default options (ultrafast bootstrap method, 1000 bootstrap alignments), and the GTR+G+I model of evolution.
2.4. Bioacoustics
We recorded the advertisement call of Pristimantis clarae sp. nov., male CORBIDI 21669 at the type locality on 18 October 2019 at an air temperature of 12.2 °C and relative humidity of 88% (taken with a digital thermometer-hygrometer to the nearest 0.1 °C and 1%). We used a digital recorder (Marantz PMD661MK2) and unidirectional microphone (Sennheiser ME64) for field recording at 48 kHz, 24-bit, WAV format, and Raven Pro version 1.5 (Cornell Laboratory of Ornithology, Ithaca, NY, USA) to analyze call variables. We analyzed 15 calls of P. clarae and 25 calls of P. albertus, a morphologically similar and sympatric species. The following variables were measured from oscillograms: note and call duration and rate, interval between notes or calls, and presence of amplitude modulation. Variables measured from spectrograms included dominant frequency, and presence of frequency modulation or harmonics. Spectral parameters were calculated through fast Fourier transform (FFT) set at a length of 512 points (Hann window, 50% overlap). Averages are reported ± SD.
2.5. Species Delimitation
The taxonomic conclusions of this study are based on the observation of morphological features, color patterns, and bioacoustic traits of the advertisement calls, as well as the inferred phylogenetic relationships [29,30,31,32]. Information that is considered as species delimitation criteria following a general lineage or unified species concept [29,30].
3. Results
3.1. Systematics
Our molecular phylogeny of the 16S gene fragment suggests that P. clarae is closely related to P. bounides Lehr, von May, Moravec, and Cusi, 2017, and P. humboldti Lehr, von May, Moravec, and Cusi, 2017 (Figure 1), two species distributed at higher elevations in the same region of Satipo in the eastern Andean slopes of central Peru (Figure 2) [18]. Pristimantis clarae thus appears to be a low-elevation relative of these two “Phrynopus-like” Pristimantis species, i.e., Pristimantis that superficially resemble Phyronopus because of their compact body shape, relatively short limbs, and reduced toe pads. The sister taxon to this hypothetical clade of three species (node support is 0.81) is P. aniptopalmatus Duellman and Hedges, 2005, from Provincia de Oxapampa, Departamento de Pasco (Figure 2) [2].
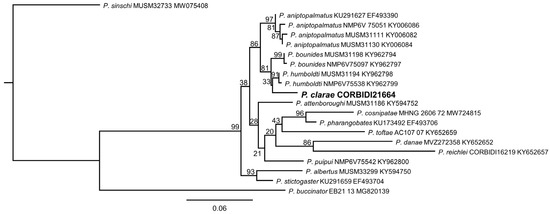
Figure 1.
Maximum Likelihood consensus tree for Pristimantis clarae and related species included in this study based on a 542-bp fragment of the 16S rRNA gene in IQ-TREE (posterior probabilities are indicated at each node). Consensus tree constructed from 1000 bootstrap trees (log-likelihood of consensus tree: −343.9, SE 104.7). Branch lengths are optimized by maximum likelihood on original alignment.
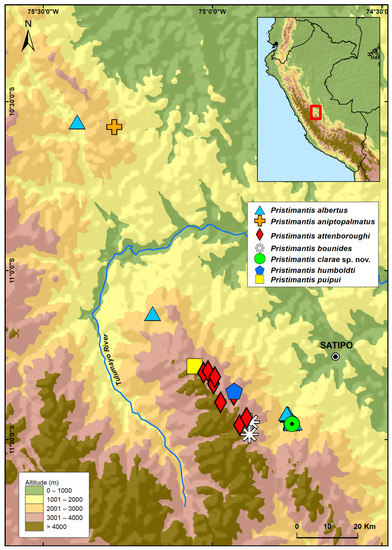
Figure 2.
Map showing the distribution of Pristimantis clarae sp. nov. and similar species of the P. danae species group. The green circle with a dot corresponds to the type locality.
Uncorrected p-distances for 16S (Supplementary S1 in Supplemental Materials) are lower than 3% for the species P. humboldti, P. attenboroughi Lehr and von May, 2017, P. aniptopalmatus and P. bounides (ranked from smallest to largest p-distance), supporting the relationships inferred from the maximum likelihood phylogeny. Although the genetic distances are relatively low compared to other pairwise distances among species of Pristimantis, even the species with smallest p-distance of 1.09%, P. humboldti is easily distinguishable from P. clarae on the basis of diagnostic meristic traits.
3.2. Taxonomy
Family Strabomantidae Hedges, Duellman and Heinicke 2008
Genus Pristimantis Jiménez de la Espada 1871
Pristimantis clarae sp. nov. Venegas, García-Ayachi, Marchelie, Ormeño and Catenazzi, 2023
LSID: urn:lsid:zoobank.org:act:11D8DF83-673E-458D-B1D6-42DD0ED1D938

Figure 3.
Pristimantis clarae sp. nov. in life: (A) dorsolateral, (B) dorsal, and (C) ventral views of CORBIDI 21664; (D) dorsolateral, (E) dorsal, and (F) ventral views of CORBIDI 21670; (G) dorsolateral, (H) dorsal and (I) ventral views of male paratype CORBIDI 21666; (J) dorsolateral, (K) dorsal, and (L) ventral views of male paratype CORBIDI 21669. Photographs by A. Marchelie.

Figure 4.
Preserved specimens of Pristimantis clarae sp. nov. (holotype) in dorsal view (A), ventral view (B), palm (C), sole (D), and head in lateral view (E). Scale 5 mm. Photographs by L.A. García-Ayachi.
3.2.1. Holotype
CORBIDI 21664, an adult female, from Fundo Palmapata, Distrito de Pampa Hermosa, Provincia de Satipo, Departamento de Junín, Perú (11.45329 S, 74.76541 W, 2199 m a.s.l) obtained on 19 October 2019 by L.A. García-Ayachi, A. Marchelie, J. Ormeño, and P.J. Venegas.
3.2.2. Paratypes
Thirteen total, all from Fundo Palmapata: CORBIDI 21663, CORBIDI 21665–66, adult males, CORBIDI 21667, an adult female, and CORBIDI 21668–74, adult males, collected with the holotype. CORBIDI 21620–21, a juvenile and adult male, respectively, (11.45006 S, 74.76752 W, 2117 m a.sl.) collected on 18 October 2019 by L.A. García-Ayachi, A. Marchelie, J. Ormeño, and P.J. Venegas.
3.2.3. Diagnosis
A new species of Pristimantis assigned to the Pristimantis danae species Group Padial, Grant and Frost, 2014, having the following combination of characters: (1) Skin on dorsum smooth to shagreen with scattered round and conical tubercles; flanks finely tuberculate; skin on venter weakly areolate or areolate; discoidal fold present or absent and thoracic fold present; dorsolateral folds present; (2) tympanic membrane and tympanic annulus present, distinct; (3) snout short, rounded or truncate in dorsal view and rounded in profile; (4) upper eyelid bearing some low conical tubercles; EW shorter than IOD; cranial crests absent; (5) dentigerous processes of vomers present, distinct, oblique; (6) males with vocal slits and small subgular vocal sac; nuptial pads absent; (7) Finger I shorter than Finger II; discs of digits expanded, truncated; (8) fingers bearing narrow lateral fringes; (9) minute conical ulnar tubercles present or absent and tarsal tubercles present, low, conical; (10) heel bearing one or two low conical tubercles; inner tarsal fold present, weakly defined, short; (11) inner metatarsal tubercle ovoid, prominent, 2 or 3 times as large as outer; outer metatarsal tubercle subconical, distinct; supernumerary plantar tubercles indistinct, scattered; (12) toes bearing narrow lateral fringes; basal toe webbing absent; Toe V longer than Toe III; toe discs slightly smaller than those on fingers; (13) in life (Figure 3), dorsum tan or brown with dark brown marks as a bar or blotch on the top of snout, interorbital bar, a conspicuous canthal stripe, labial bars, supratympanic stripe, triangular blotches on dorsum, diagonal streaks on flanks and diagonal bars on limbs; groins dusty brown with or without scattered cream minute flecks, anterior and posterior surface of thighs dark brown with minute pale cream flecks and with or without some scattered pale cream blotches; ventral surface on throat, belly and hind limbs dirty cream with a dense dark brown mottling, usually with a V-shaped mark below the chin; iris coppery, with the upper half bright and the lower dark, bearing flecks, speckles or vermiculation irradiating from the pupil; sclerotic ring whitish gray; (14) SVL 12.9–15.6 mm (n = 11) in males and 17.6–19.3 mm (n = 2) in females.
3.2.4. Comparison with Other Species
Pristimantis clarae can be distinguished from other species of the genus by the following combination of characters: (i) a rounded or truncate snout in dorsal view, (ii) presence of dorsolateral folds, (iii) tympanic membrane and annulus distinct, (iv) absence of flash marks on the hidden surfaces of flanks and hindlimbs, and (v) a small size (maximum SVL 15.6 mm in males and 19.3 mm in females).
Compared to other species of Pristimantis from the Cordillera Oriental of central Peru, the new species is most similar to the sympatric P. albertus. Both species were common at the type locality, are similar in size (SVL 12.9–15.6 mm in males and 17.6–19.3 mm in females of P. clarae versus 12.9–19.5 in males and 19.7–24.2 in females of P. albertus; Duellman and Lehr [2], Moravec, Lehr and Kodejs [17]), and share the presence of dorsolateral folds. Moreover, both species belong to the P. danae species group. However, the new species differs from P. albertus (state of characters in parenthesis) by having low conical tubercles on hells and outer edge of tarsus (absent in P. albertus) and usually a V-shaped mark below the chin (absent). Additionally, the throat and vocal sac of adult males of P. albertus is yellow, yellowish cream, greenish yellow, or orange with brown or cream flecks or reticulations; while it is tan, brown or cream with dark brown speckles or reticulations in the new species. The two species have very different advertisement calls (Figure 5), consisting of a long thrill of up to 46 notes, ~50–60 ms in duration in P. albertus, and a shorter call of 4–7 notes, <10 ms long, in P. clarae, despite both species having similar peak frequencies.
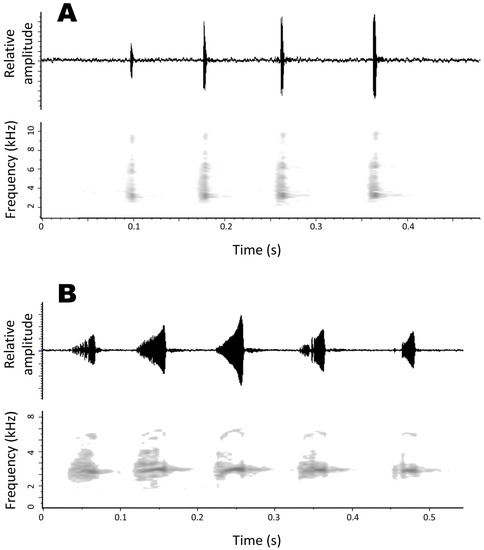
Figure 5.
Advertisement calls of two species of Pristimantis from the Cordillera Oriental of central Peru. (A) advertisement call (short call) of male CORBIDI 21669 (SVL 15.6 mm), paratype of Pristimantis clarae sp. nov., recorded on 18 October 2019 (Tair = 12.2 °C), illustrating amplitude modulation and last note interval being longer than the interval among the preceding notes. (B) Advertisement call of Pristimantis albertus CORBIDI 21658 (SVL 14.1 mm), recorded at the type locality on 18 October 2019 (Tair = 12.2 °C).
Another sympatric species that possesses dorsolateral folds, P. bipunctatus Duellman and Hedges, 2005, is larger (SVL 22.6–28.8 mm in males and 32.4–41.5 mm in females) and lacks conical tubercles on eyelids, heels and outer surface of tarsus. Moreover, the ventral surface is cream or grayish white in P. bipunctatus, whereas it is dirty cream or tan with a dense dark brown mottling or reticulations in the new species.
Other species of Pristimantis from the Cordillera Oriental of central and southern Peru [2] that share with P. clarae the presence of dorsolateral folds and belong to the P. danae species group are: P. aniptopalmatus Duellman and Hedges, 2005, P. attenboroughi, P. bounides, P. danae Duellman, 1978, P. humboldti, P. ornatus Lehr, Lundberg, Aguilar, and von May, 2006, P. pharangobates Duellman, 1978, P. puipui Lehr, von May, Moravec, and Cusi, 2017, P. rhabdolaemus Duellman, 1978, and P. stictogaster Duellman and Hedges, 2005. Of which, P. aniptopalmatus, P. danae, P. ornatus, and P. stictogaster differ from P. clarae by lacking conical tubercles on heels and outer edge of tarsus. Pristimantis pharangobates and P. rhabdolaemus share with P. clarae the presence of tubercles on eyelids, heels and the outer edge of tarsus. Nevertheless, both species can be distinguished from P. clarae by having the snout long and subacuminate in P. pharangobates, and narrowly rounded in P. rhabdolaemus (short and rounded in the new species).
Pristimantis attenboroughi and P. puipui can be easily distinguished from P. clarae by lacking tympanic annulus and membrane, and by having the tips of digits narrow, rounded and lacking circumferential groves. The new species possesses a distinct tympanic annulus and membrane and the tips of digits bearing expanded discs with well-defined circuferential grooves. The new species differs from P. bounides (state of characters in parentheses) by having the skin on flanks finely tuberculate (tuberculate), discs of digits expanded and truncated (sligthly expanded and round), dorsolateral fold continuous (irregular and discontinuous), and basal toe webbing absent (present). Pristimantis humboldti can be readily distinguished from P. clarae by having the groins blackish brown with orange, white or pale gray spots, while in the new species are dusty brown with or without scattered cream minute flecks. Additionally, discs on digits are truncate in P. clarae and rounded in P. humboldti.
3.2.5. Description of the Holotype
An adult female (Figure 4A,B) with the head slightly narrower than body, slightly wider than long; head length 32% of SVL; head width 37% of SVL; cranial crests absent; snout moderately short, rounded in dorsal and lateral views; eye-nostril distance 71% of eye diameter; nostrils not protuberant, directed dorsolaterally; canthus rostralis slightly curved in profile, concave in dorsal view; loreal region concave; lips rounded; upper eyelid each with several small round tubercles, heterogenous in size; upper eyelid width 38% of IOD; occipital and scapular region lacking folds; supratympanic fold distinct, short, extending from the posterior corner of the eye curving toward the arm insertion, concealing the dorsal and posterior edge of tympanum; tympanic membrane and annulus present, distinct; several postrictal tubercles present, round, with two distinctly enlarged bilaterally. Choanae large, oval, not concealed by palatal shelf of maxilla; dentigerous processes of vomers present, distinct, oblique, with several minute teeth each; tongue longer than wide, barely notched posteriorly, covering most of floor of mouth, posterior third free.
Skin on dorsum shagreen sprinkled with enlarged round and subconical tubercles, including on the head, and a hairline mid-dorsal weakly defined fold from the tip of snout to the upper margin of cloacal sheath, flanks tuberculate, dorsolateral folds present; skin on throat, chest and belly weakly areolate; discoidal fold visible laterally and thoracic folds present, distinct; cloacal sheath short.
Outer ulnar surface with two low conical tubercles; palmar tubercle deeply divided; thenar tubercle ovoid, prominent; subarticular tubercles prominent, round in ventral and lateral views; supernumerary tubercles present, few, most similar in size to the subarticular tubercles; fingers bearing narrow lateral fringes; Finger I shorter than Finger II; discs on Finger I narrow, round; disc on Finger II slightly expanded, round; disc on Finger III and IV expanded, truncate, about twice the size of digit proximal to it (Figure 4C).
Hind limbs long, slender, tibia length 55% of SVL; foot length 47% of SVL; upper surfaces of hind limbs covered by round tubercles, heterogenous in size; inner and posterior surface of thighs smooth, ventral surfaces of thighs weakly areolate; knees with several minute round tubercles; heels bearing one low conical tubercles; outer surface of tarsus with some low, small, conical tubercles; inner tarsal fold present, weakly defined, short; inner metatarsal tubercle distinct, ovoid, three times the size of distinct, round outer metatarsal tubercle; subarticular tubercles distinct, round in ventral and lateral views; plantar supernumerary tubercles weak, about one third or half the size of subarticular tubercles; toes bearing narrow lateral fringes; basal webbing absent; discs on digit of toes about as large as those on fingers, truncate; relative length of toes: 1 < 2 < 3 < 5 < 4; distal edge of disc on Toe III reaching to proximal margin of distal subarticular tubercle on Toe IV, distal edge of disc on Toe V extends to the middle of distance between distal and hiperdistal subarticular tubercle on Toe IV (Figure 4D).
Measurements (in mm) of the holotype: SVL 17.5; tibia length 9.7; foot length 8.3; head length 5.5; head width 6.6; eye diameter 2.5; tympanic diameter 0.9, IOD 2.1; upper eyelid width 0.8; internarial distance 1.89; eye-nostril distance 1.8.
Coloration of holotype: In life (Figure 3A–C), dorsum dark cinnamon brown with a dark brown interorbital bar and a faint brown triangular blotch of the middle on back (both marks with thin cream borders), and dorsolateral folds tan; sides of head, flanks and limbs dusty brown; sides of head with a conspicuous dark brown canthal stripe, labial bars, and supratympanic mark, tympanic membrane tan with coppery hue; flanks with dark brown diagonal streaks bearing cream borders, groins dark brown with few minute cream flecks; hind limbs with dark brown diagonal bars bearing thin cream borders and posterior surface of thighs dark brown with faint cream flecks; ventrally, throat, chest belly and hind limbs dirty cream with a dense dark brown mottling; forelimbs, palms and soles dark brown with dirty cream flecks or speckles; iris coppery, with the upper half bright and the lower dark, bearing dark brown speckles and vermiculation irradiating from the pupil; sclerotic ring whitish gray.
In ethanol 70% (Figure 4), the dorsum turns tan, the interorbital bar disappears and the triangular blotch of the middle of black is faint, and the dorsolateral folds are paler than dorsum; sides of head, flanks and limbs are dark brown with its marks darker; posterior surface of thighs dark brown with whitish cream flecks and speckles; ventral surface brown with a dirty cream mottling, discs grayish cream, palms and soles with cream blotches.
3.2.6. Intraspecific Variation
Sexual dimorphism is noticeable in size, males possess vocal slits and are smaller (SVL = 12.8–15.6 mm) than females (SVL = 17.5–19.3 mm), and both sexes differ slightly in some proportions (Table 1).

Table 1.
Variation of measurements (in mm) and proportions of the type series of Pristimantis clarae sp. nov. Mean ± SD is given with range in parentheses. See text for abbreviations.
The dorsal background coloration varies from creamy tan (CORBIDI 21665) to cinnamon brown (CORBIDI 21668) and dark brown (CORBIDI 21669). The dorsal marks on the back are faint or darker than background coloration (CORBIDI 21666), and the marks can also have dirty cream contrasting borders (see CORBIDI 21669). The dorsal pattern varies from one to two triangular dorsal mark (CORBIDI 21666), or chevrons (CORBIDI 21663) or one quadrangular mark (CORBIDI 21665). Dorsolateral folds are usually distinctly paler than dorsal background coloration, are tan (CORBIDI 21670) or dirty cream (CORBIDI 21669), but also can lacking contrast (CORBIDI 2162). Ventrally, some individuals possess a dark brown V-shaped mark below the chin (CORBIDI 21666).
3.2.7. Advertisement Call
We recorded 15 calls of CORBIDI 21669 on 18 October 2019 at 12:20 (Figure 5A). At a Tair = 12.2 °C, the advertisement call averaged 421 ± 24 ms in duration (range 277–634 ms, n = 15) and consisted of 4–7 notes (11 calls with four notes, two calls with five notes, and one call each with six and seven notes) produced at a rate of 0.21–0.48 calls/s (across two recordings, 10 calls in ~21 s, and 5 calls in ~24 s), and at intervals of 2950 ± 1195 ms (range 1243–17184 ms, n = 13). The notes averaged 9.5 ± 0.3 ms in duration (range 5–16 ms, n = 68 notes), emitted at intervals of 75 ± 6 ms (range 32–140 ms, n = 50 note intervals). Intervals among notes tend to increase towards the end of the call, with last interval averaging 99 ms ± 22 ms (range 32–261 ms, n = 13 intervals). Note amplitude generally increases throughout the call. Peak frequency averaged 3156 ± 17 Hz (range 3070–3281 Hz, n = 15). No harmonics were visible in the recordings.
3.2.8. Distribution and Ecology
Pristimantis clarae is currently only known for its type locality, a cloud forest in the Pampa Hermosa River basin at elevations from 2117 to 2199 m, Department of Junín, in the Amazonian slopes of the Cordillera Oriental of central Peru (Figure 2). Following the terrestrial ecoregions of the world by Olson, et al. [33], the new species inhabits the Peruvian Yungas ecoregion. All individuals were collected on ferns, Chusquea spp. branches, and herbaceous vegetation at heights from 10 cm to 130 cm. The habitat where P. clarae was collected is a primary forest with a canopy about 5 to 8 m high on a steep slope. The dominant vegetation is the mountain bamboo, Chusquea spp. (Poaceae), but arboreal ferns are abundant too. Sympatric anurans were P. albertus, P. bipunctatus, and Pristimantis sp. Individuals of P. albertus and P. clarae were abundant at the type locality by night. Males of both species were found calling on ferns and herbaceous vegetation up to about 50 cm from the ground. Males of P. albertus were calling while perched on fern leaves or on top of herbaceous vegetation, whereas males of P. clarae were calling hidden in the herbaceous vegetation.
3.2.9. Etymology
The specific name “clarae” is for Clara Alexandrova and is treated as a latinized feminine noun in the genitive case. Clara Alexandrova is a young frog enthusiast that together with her family support the conservation of cloud forest ecosystems in Peru through the nonprofit organization Rainforest Partnership, Austin, TX, USA. The cloud forests harbor the greatest diversity of Pristimantis and are one of the most threatened ecosystems in the world.
3.3. Advertisement Call of Pristimantis albertus
We recorded the call of CORBIDI 21658 on 18 October 2019 at 12:20 (Figure 5B), at an Tair = 12.2 °C. This male emitted two types of advertisement calls, a long call nearly 6 s long and composed of 46 notes (single recording), and a short call composed of up to 8 notes (n = 24 calls). Except for the number of notes, the peak frequency and note duration were similar for both types of calls. The notes of the long call averaged 52 ± 2 ms in duration (range 32–75 ms, n = 46), emitted at intervals of 79 ± 2 ms (range 53–112 ms, n = 45). The short call averaged 287 ± 111 ms in duration (range 149–779 ms, n = 24) and consisted of 2–8 notes (16 calls with three notes, 4 calls with four notes, 2 calls with two notes, and one call each with 5 and 8 notes) produced at a rate of 0.13 calls/s (24 calls in ~188 s). The notes of the short call averaged 61 ± 2 ms in duration (range 27–96 ms, n = 24), emitted at intervals of 44 ± 3 ms (range 16–91 ms, n = 24). Peak frequency across notes averaged 3312 ± 8 Hz (range 3094–3375 Hz, n = 46 notes of one call) in the long call, and 3319 ± 23 Hz (range 3094–3563 Hz, n = 24 calls) in the short calls. In addition to the band of peak amplitude around 3316–3366 Hz, several notes have up to three harmonics around ~6912, 10,444, and 13,824 Hz.
4. Discussion
We used an integrative taxonomy approach that combined molecular, morphological and bioacoustics traits of the advertisement calls data to describe a new species of Pristimantis from the Andean montane forests of central Peru. We used the gene 16S to infer the phylogenetical relationships of P. clarae and to calculate genetic distances. Our comparison with close relatives supports the separation of P. clarae from its most similar species like the sympatric P. albertus and P. aniptopalmatus. The 16S fragment as well as the gene COI are commonly used in amphibian DNA barcoding [34] and to explore cryptic diversity [35].
Based on our phylogram, we assign the new species described herein to the P. danae species group. The P. danae species group is composed of 14 species (i.e., P. albertus, P. aniptopalmatus, P. attenboroughi, P. bounides, P. cuneirostris Duellman and Pramuk, 1999, P. danae, P. humboldti, P. ornatus, P. pharangobates, P. puipui, P. reichlei Padial and De la Riva, 2009, P. rhabdolaemus, P. sagittulus Lehr, Aguilar and Duellman, 2004, P. stictogaster, and P. toftae Duellman, 1978), most of them distributed in the Andean montane forest of central and southern Peru [18]. Although we used only the 16S gene, the clade composed of the species of P. danae species group and P. clarae has strong support (See Figure 1). Additionally, the presence of dorsolateral folds in P. clarae, a morphological character common in the P. danae species group, and its distribution supports its assignation to this monophyletic group.
Our phylogram shows that P. clarae is closely related to P. bounides, P. humbodti, and P. aniptopalmatus (Figure 1), the two first species distributed at higher elevations (above 3300 m) in the same region of Satipo, like P. attenboroughi and P. puipui; and P. aniptopalmatus in the region of Oxapampa 120 km NW of the type locality of P. clarae (See Figure 2). Although the genetic distances are low (<3%), and as low as 1.09% with P. humboldti (see Supplementary S1 in Supplemental Materials), P. clarae can be readily distinguished from these species by morphological traits (e.g., snout shape, presence and condition of dorsolateral folds, presence or absence of tympanic membrane and annulus distinct, prescense of turbercles on eyelids, heels and outer edge of tarsus, presence or absence of flash marks on the hidden surfaces of flanks and hindlimbs) that are commonly used in the description of new species of Pristimantis, e.g., [6,15,18,20]. Moreover, species such as P. attenboroughi, P. bounides, P. humboldti, and P. puipui that are phylogenetically and geographically closest (see Figure 2) to P. clarae, superficially resemble the high-elevation genus Phyronopus because they have a compact body shape, relatively short limbs, and reduced toe pads.
With 107 new species named, Peru is currently the country with the third highest number of new species of Pristimantis described since 2010, following Ecuador and Colombia with 212 and 168 species, respectively [36]. However, the accumulation curve of described species is far from reaching saturation. The number of new species of Pristimantis described in the Cordillera Oriental of central Peru has reached 34 species in the last two decades [6,7,8,9,10,11,12,13,14,15,16,17,18] and the amount of information of each described species is constantly improving [10,14,17]. However, most species are currently known from few specimens and localities (e.g., P. adiastolus Duellman and Hedges, 2007, P. aniptopalmatus, P. eurydactylus Hedges and Schlüter, 1992, P. boucephalus Lehr, Moravec, Cusi, and Gvoždík, 2017, P. minutulus Duellman and Hedges, 2007, and P. rhabdocnemus Duellman and Hedges, 2005).
Moravec et al. [17] provided additional data about morphological variation and systematics of P. albertus, a poorly known species described from the Yanachaga-Chemillen National Park [2,37]. Herein, we describe for the first time two types of advertisement calls of P. albertus. The species was common in our surveys at Pampa Hermosa River at elevations from 1800 to 2199 m. Following these recent contributions, P. albertus is now the Pristimantis species of central Peru best known with respect to taxonomy and distribution. Such knowledge is crucial for amphibian conservation in countries such as Peru, where amphibian diversity and distribution patterns are not yet well understood.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/taxonomy3020019/s1, Supplementary S1. Genetic distances (uncorrected p-distances) estimated from 542 bp of the non-coding 16S rRNA mitochondrial fragment between Pristimantis clarae and related taxa of the genus Pristimantis (Strabomantidae).
Author Contributions
Field data recollection, A.M., J.R.O., L.A.G.-A. and P.J.V.; conceptualization, P.J.V. and L.A.G.-A.; methodology, A.C. and P.J.V.; software, A.C. and L.A.G.-A.; validation, P.J.V., L.A.G.-A. and A.C.; formal analysis, P.J.V. and A.C.; investigation, P.J.V. and L.A.G.-A.; resources, P.J.V.; data curation, L.A.G.-A.; writing—original draft preparation, P.J.V.; writing—review and editing, A.C. and L.A.G.-A.; visualization, P.J.V.; supervision, A.C.; project administration, P.J.V.; funding acquisition, P.J.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Rain Forest Partnership through Hollomon Price Foundation.
Institutional Review Board Statement
The Florida International University IACUC (protocols #IACUC-18-009 and #IACUC-21-017) reviewed and approved animal capture, handling and collecting techniques used in this study.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful with the residents of the San Antonio village who allowed our herpetological research. We are indebted with Juan Mayta for loan us his inn and Jhordy Orcuhuaranga for his logistic support during the expedition.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
The following specimens were examined: Pristimantis albertus. PERU: DEPARTAMENTO JUNÍN: Pampa Hermosa: Juan de Dios, 1600–1900 m: CORBIDI 21507, 21510, 21512, 21513, 21517, 21535-38, 21541, 25143-44; Fundo La Palma (Juan Mayta), 1876–2174 m: CORBIDI 21634-35, 21647, 21658-61. DEPARTAMENTO PASCO: Oxapampa: Concesión de Conservacion La Suiza—Ulcumano Ecolodge, 2220 m; CORBIDI 21766-77; Pristimantis aniptopalmatus, DEPARTAMENTO PASCO: Oxapampa: Abra Esperanza (Parque Nacional Yanachaga Chemillen): 2748 m: CORBIDI 10292, 11619, 11620, 11632-35, 11639, 11640-42; Pristimantis bipunctatus, DEPARTAMENTO PASCO: Oxapampa: Concesión de Conservación La Suiza- Ulcumano Ecolodge, 2220 m; CORBIDI 21757, 21759, 21783-84, 21794, 21800; DEPARTAMENTO JUNÍN: Pampa Hermosa: Fundo La Palma (Juan Mayta), 1876–2174 m: CORBIDI 21576, 21622-23, 21642-43, 21645, 21655, 21657, 21757, 21759, 21783-84, 21794, 21800; Pristimantis rhabdolaemus, DEPARTAMENTO AYACUCHO: La Mar: Pampa Aurora, 1246 m: CORBIDI 10775; San Antonio, 1246 m: CORBIDI 10808; Chiquintirca, 2635 m: CORBIDI 10813, 10815-16; Pristimantis stictogaster, DEPARTAMENTO PASCO: Oxapampa: Parque Nacional Yanachaga Chemillen: Oso Playa, 2500 m: CORBIDI 7426-27; Abra Esperanza 2808 m: CORBIDI 10287-88; Comunidad Campesina Chacos, 2901 m: CORBIDI 3826, 3850.
References
- AmphibiaWeb. Available online: https://amphibiaweb.org (accessed on 8 February 2023).
- Duellman, W.E.; Lehr, E. Terrestrial Breeding Frogs (Strabomantidae) in Peru; Natur und Tier-Verlag GmbH: Munster, Germany, 2009; p. 382. [Google Scholar]
- Hedges, S.B.; Duellman, W.E.; Heinicke, M.P. New World direct-developing frogs (Anura: Terrarana): Molecular phylogeny, classification, biogeography, and conservation. Zootaxa 2008, 1737, 1–182. [Google Scholar] [CrossRef]
- Campbell, J.A.; Lamar, W.W. The Venomous Reptiles of the Western Hemisphere; Comstock Pub. Associates: Ithaca, NY, USA, 2004; 1032p, Volume 1. [Google Scholar]
- Lehr, E. Amphibien und Reptilien in Peru: Die Herpetofauna Entlang des 10. Breitengrades von Peru: Arterfassung, Taxonomie, Okologische Bemerkungen und Biogeographische Beziehungen; Natur und Tier-Velag: Munster, Germany, 2002; p. 208. [Google Scholar]
- Duellman, W.E.; Hedges, S.B. Eleutherodactyline frogs (Anura: Leptodactylidae) from the Cordillera Yanachaga in central Peru. Copeia 2005, 2005, 526–538. [Google Scholar] [CrossRef]
- Duellman, W.E.; Hedges, S.B. Three new species of Pristimantis (Lissamphibia, Anura) from montane forests of the Cordillera Yanachaga in Central Peru. Phyllomedusa 2007, 6, 119–135. [Google Scholar] [CrossRef]
- Boano, G.; Mazzotti, S.; Sindaco, R. A new peculiar frog species of the genus Pristimantis from Yanachaga-Chemillén National Park, Peru. Zootaxa 2008, 1674, 51–57. [Google Scholar] [CrossRef]
- Chávez, G.; Catenazzi, A. A new species of frog of the genus Pristimantis from Tingo Maria National Park, Huanuco Department, central Peru (Anura, Craugastoridae). Zookeys 2016, 610, 113–130. [Google Scholar] [CrossRef] [PubMed]
- Chávez, G.; García-Ayachi, L.A.; Catenazzi, A. Beauty is in the eye of the beholder: Cruciform eye reveals new species of direct-developing frog (Strabomantidae, Pristimantis) in the Amazonian Andes. Evol. Syst. 2021, 5, 81–92. [Google Scholar] [CrossRef]
- Duellman, W.E.; Chaparro, J.C. Two distinctive new species of Pristimantis (Anura: Strabomantidae) from the Cordillera Oriental with a distributional synopsis of strabomantids in Central Peru. Zootaxa 2008, 1918, 12–25. [Google Scholar] [CrossRef]
- Lehr, E.; Aguilar, C.; Duellman, W.E. A striking new species of Eleutherodactylus from Andean Peru (Anura: Leptodactylidae). Herpetologica 2004, 60, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Lehr, E.; Lundberg, M.; Aguilar, C.; Von May, R. New species of Eleutherodactylus (Anura: Leptodactylidae) from the eastern Andes of central Peru with comments on central Peruvian Eleutherodactylus. Herpetol. Monogr. 2006, 20, 105–128. [Google Scholar] [CrossRef]
- Lehr, E.; Moravec, J. A new species of Pristimantis (Amphibia, Anura, Craugastoridae) from a montane forest of the Pui Pui Protected Forest in central Peru (Región Junín). Zookeys 2017, 645, 85–102. [Google Scholar] [CrossRef]
- Lehr, E.; Moravec, J.; Cusi, J.C.; Gvoždík, V. A new minute species of Pristimantis (Amphibia: Anura: Craugastoridae) with a large head from the Yanachaga-Chemillén National Park in central Peru, with comments on the phylogenetic diversity of Pristimantis occurring in the Cordillera Yanachaga. Eur. J. Taxon. 2017, 325, 1–22. [Google Scholar] [CrossRef]
- Lehr, E.; von May, R. A new species of terrestrial-breeding frog (Amphibia, Craugastoridae, Pristimantis) from high elevations of the Pui Pui Protected Forest in central Peru. Zookeys 2017, 660, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Moravec, J.; Lehr, E.; Kodejs, K. A new species of Pristimantis (Amphibia, Anura, Strabomantidae) from the Pui Pui Protected Forest (central Peru), with comments on Pristimantis albertus Duellman & Hedges, 2007. Zookeys 2020, 994, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Lehr, E.; Von May, R.; Moravec, J.; Cusi, J.C. Three new species of Pristimantis (Amphibia, Anura, Craugastoridae) from Upper Montane Forests and High Andean Grasslands of the Pui Pui Protected Forest in Central Peru. Zootaxa 2017, 4299, 301–336. [Google Scholar] [CrossRef]
- Lehr, E.; Lyu, S.; Catenazzi, A. A new, critically endangered species of Pristimantis (Amphibia: Anura: Strabomantidae) from a mining area in the Cordillera Occidental of northern Peru (Región Cajamarca). Salamandra 2021, 57, 15–26. [Google Scholar]
- Páez, N.B.; Ron, S.R. Systematics of Huicundomantis, a new subgenus of Pristimantis (Anura, Strabomantidae) with extraordinary cryptic diversity and eleven new species. Zookeys 2019, 868, 1–112. [Google Scholar] [CrossRef]
- Venegas, P.J.; Duellman, W.E. Two syntopic new species of the Pristimantis orestes Group (Anura: Strabomantidae) from Northwestern Peru. Zootaxa 2012, 3249, 47–59. [Google Scholar] [CrossRef]
- Catenazzi, A.; von May, R. Conservation status of amphibians in Peru. Herpetol. Monogr. 2014, 28, 1–23. [Google Scholar] [CrossRef]
- Pinheiro, H.T.; Moreau, C.S.; Daly, M.; Rocha, L.A. Will DNA barcoding meet taxonomic needs? Science 2019, 365, 873–874. [Google Scholar] [CrossRef]
- Scott, N.J. Complete species inventories. In Measuring and Monitoring Biological Diversity: Standard Methods for Amphibians; Heyer, W.R., Donnelly, M.A., McDiarmid, R.W., Hayek, L.C., Foster, M.S., Eds.; Smithsonian Institution Press: Washinton, DC, USA, 1994; pp. 78–84. [Google Scholar]
- Lynch, J.D.; Duellman, W.E. Frogs of the Genus Eleutherodactylus (Leptodactylidae) in Western Ecuador: Systematic, Ecology, and Biogeography; Natural History Museum, The University of Kansas: Lawrence, KS, USA, 1997; p. 236. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- de Queiroz, K. The general lineage concept of species, species criteria, and the process of speciation. In Endless Forms: Species and Speciation; Howard, D.J., Berlocher, S.H., Eds.; Oxford University Press: Oxford, UK, 1998; pp. 57–75. [Google Scholar]
- de Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Frost, D.R.; Kluge, A.G. A consideration of epistomology in Systematic Biology, with special reference to species. Cladistics 1994, 10, 259–294. [Google Scholar] [CrossRef]
- Padial, J.M.; Miralles, A.; De la Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’Amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial ecoregions of the world: A new map of life on earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Crawford, A.J.; Lips, K.R.; Bermingham, E. Epidemic disease decimates amphibian abundance, species diversity, and evolutionary history in the highlands of central Panama. Proc. Natl. Acad. Sci. USA 2010, 107, 13777–13782. [Google Scholar] [CrossRef]
- Ortiz, D.A.; Hoskin, C.J.; Werneck, F.P.; Réjaud, A.; Manzi, S.; Ron, S.R.; Fouquet, A. Historical biogeography highlights the role of Miocene landscape changes on the diversification of a clade of Amazonian tree frogs. Org. Divers. Evol. 2022, 23, 395–414. [Google Scholar] [CrossRef]
- Reyes-Puig, C.; Mancero, E. Beyond the species name: An analysis of publication trends and biases in taxonomic descriptions of rainfrogs (Amphibia, Strabomantidae, Pristimantis). ZooKeys 2022, 1134, 73–100. [Google Scholar] [CrossRef]
- Chavez, G.; Cosmopolis, C.H.; Lujan, L. Annotated checklist and ecological notes of anurans from the southern region of Yanachaga Chemillen National Park, central Andes of Peru. Herpetotropicos 2012, 8, 23–28. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).