Abstract
Zamia magnifica (Zamiaceae), a new species endemic to Sierra Norte, Oaxaca, Mexico, is described. Zamia magnifica is characterized by having a rupicolous habit, pendent leaves bearing leaflets that are densely tomentose and pink to caramel in color when emerging, and broad oblong leaflets that are coriaceous in texture with few non-prominent denticulations and ovulate strobili with short (<4 cm) peduncles. It is compared to Z. furfuracea and Z. meermanii, the Mesoamerican Zamia species to which it shares the closest morphological resemblance.
1. Introduction
Cycads represent an ancient lineage of seed plants that now forms the second most diverse group of gymnosperms, just after the conifers [1]. The modern cycads are classified into ten genera, four of which (Zamia, Dioon, Ceratozamia and Microcycas) occur exclusively in the New World [2]. Within the New World, cycads are distributed into three distinct biogeographic regions [2]: (i) the Caribbean region which hosts two genera (Microcycas and Zamia) and nine species, (ii) the South America and Central American Isthmus region which hosts only a single genus (Zamia) with 49 species, and (iii) the Mesoamerican region (here referred to as ‘Mesoamerica’ and including the Mesoamerican Dominion and Mexican Transition Zone [3], which hosts three genera and 83 species [4].
Because the Mesoamerican region has the highest species and genus richness in the New World, it is considered a global hotspot for cycad diversity. Specifically, the region includes the entirety of the genera Ceratozamia (39 spp.) and Dioon (18 spp.) and almost a third of the genus Zamia (26 out of 84 total species). The latter is the most species rich, morphologically variable, ecologically diverse, and broadly distributed, with the highest species richness occurring in the South America and Central American Isthmus region [5]. Although the taxonomy of the genus Zamia remains in flux, the majority of discoveries in recent years have occurred in South America, including the description of nine new species, e.g., [6,7,8,9,10,11,12], and numerous taxonomic and nomenclatural clarifications, e.g., [13,14,15]. In contrast, the taxonomy of Mesoamerican Zamia species has remained relatively stable in recent years. A taxonomic revision of the genus in Mesoamerica was published in 2009 [16], and since then, only four new species [17,18,19,20] and a few taxonomic clarifications [21,22,23] have been published. In this contribution, we add to the body of knowledge of Mesoamerican Zamia by describing a novel rupicolous species that was recently discovered in the karstic mountainous area of Sierra Norte, Oaxaca, Mexico and comparing it to the two species to which it bears the closest morphological resemblance: the Belizean species Z. meermanii Calonje and the Mexican species Z. furfuracea L.f.
2. Materials and Methods
Fieldwork focused on investigating the putative new species was conducted in the August of 2022 in Sierra Norte, Oaxaca, Mexico (Figure 1). This included the collection of morphometric data, plant material (12 leaves from different adult plants, two seed cones, and four pollen cones), herbarium specimens, and photographs. This amount of collected material was decided by the local people, who actively protect the plants and who guided us to the location where the plants occur. A review of our extensive collective database of herbarium specimen data and images confirmed that the species of interest was unique and had not been collected previously. The collected material was subjected to diagnosis, and we found that the most morphologically similar species were the Belizean species Z. meermanii and the Mexican species Z. furfuracea (Figure 1). Thus, we examined qualitative traits that could be used to identify the differences between these species and conducted morphometric analyses to compare them (Table 1). For Z. meermanii, we used morphometric data collected in August of 2008 in Soldier Creek, Belize District, Belize, during the course of the fieldwork that resulted in the original species description [17]. For Z. furfuracea, in situ morphometric data were collected in Alvarado, Veracruz, Mexico in March of 2023. A total of 12 individuals of the putative new species, 15 of Z. meermanii, and 9 of Z. furfuracea were compared morphometrically utilizing twelve traits typically used in species delimitation in cycads, e.g., [24,25,26,27] (Table 1). All statistical tests were performed in the PAST v3.14 software [28]. To estimate the global differentiation among species, we used the ANOVA and Kruskal–Wallis tests. This approach deals with the differences in the sample size variation among species. We also used a Linear Discriminant Analysis (LDA) to estimate how much the three species can be distinguished when considering the multivariate data.
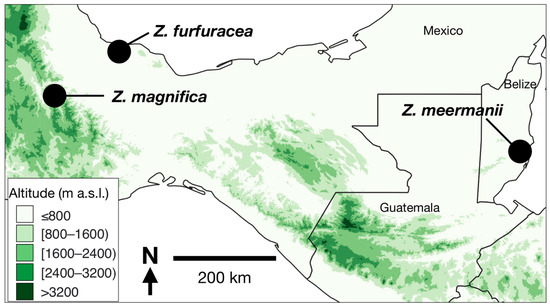
Figure 1.
Locations of the three species that we compared in this study: Zamia magnifica from La Chinantla, Sierra Norte, Oaxaca, Mexico; Z. furfuracea from Alvarado, Veracruz, Mexico; Z. meermanii from Soldier Creek, Belize Department, Belize.

Table 1.
List of 12 morphometric traits evaluated in Z. magnifica and two closely related species: Z. meermanii and Z. furfuracea. Overall differentiation estimated as Welch’s F in ANOVA and p-values are indicated. D.f. = degrees of freedom.
3. Results
3.1. Morphometric Analysis
Welch’s ANOVA demonstrated that the 12 quantitative traits have overall significantly different mean values among the three compared species (Table 1). However, in pairwise comparisons, the number of leaflets did not show significant Tukey’s Q values (Table 2, Figure 2), suggesting that it is not useful as a diagnostic trait. In contrast, all the remaining 11 traits were significantly differentiated in any of the three pairwise comparisons. In particular, the number of leaves, length of petioles, length of basal leaflets, length of median leaflets, length of apical leaflets, width of median leaflets, width of apical leaflets, and distance between median leaflets were the traits that are significantly differentiated among the three species (their mean values do not overlap). The length of rachis, the width of basal leaflets, and the width of articulations of median leaflets were significantly differentiated between Z. magnifica and Z. meermanii, and between Z. magnifica and Z. furfuracea, but not between Z. meermanii and Z. furfuracea.
Table 2.
Pairwise Tukey’s Q values of differentiation between the new species (mag = Z. magnifica) and the other two species examined species (fur = Z. furfuracea and mee = Z. meermanii) of 12 morphometric traits. Abbreviations: * p < 0.05; *** p < 0.001; ns, no significant.
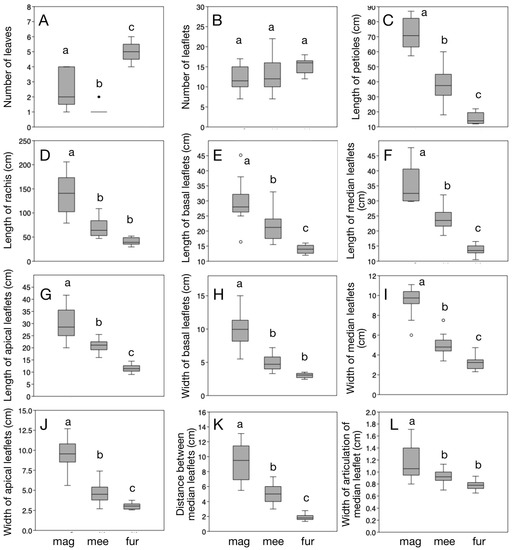
Figure 2.
Morphological variation and differentiation of 12 traits in Zamia magnifica (mag), Z. meermanii (mee), and Z. furfuracea (fur). Panels (A–L) correspond to the traits listed in Table 1 and Table 2. Different lowercase letters above the bars indicate significant differentiations, as obtained in the Tukey’s Q pairwise estimations. Dots and open dots above or below the boxplots indicate outliers.
The LDA summarized the total variation among species in two axes, each representing 67.97% (axis 1), 32.03% (axis 2) of the total variation (Figure 3). The biplots C and D suggest that the length of petioles and rachides were the main traits that dispersed the three groups. The confusion matrix obtained from LDA (Table 3) confirmed that 100% of the individuals evaluated were classified correctly, suggesting a complete sorting among species. The squared Mahalanobis distances (Table 4) were significantly high in all pairwise comparisons (p < 0.5), also suggesting that the overall morphometric variation is completely sorted among the examined taxa, confirming that the Z. magnifica represents a different species.
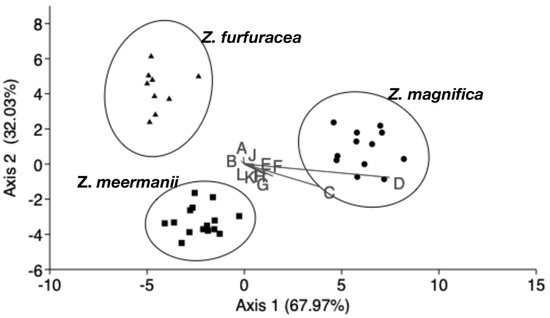
Figure 3.
Linear Discriminant Analysis plot. The two main axes summarize 100% of the total variation. The 95% ellipses of species do not overlap, indicating a clear overall differentiation among the three species: Zamia magnifica (circles), Z. furfuracea (triangles), Z. meermanii (squares). The biplots A–L correspond to the traits listed in Table 1. The length and direction of the biplots indicate the relative importance of each trait in representing the variation among species.
Table 3.
Confusion matrix constructed with the Linear Discriminant Analysis (LDA). Rows indicate given groups (number of individuals considered Z. magnifica, Z. meermanii, or Z. furfuracea) and columns indicate groups predicted by the LDA.
Table 4.
Squared Mahalanobis distances (below diagonal) and p-values (above diagonal) among the three analyzed species. All values are significant, suggesting complete sorting among species.
3.2. Taxonomic Treatment
Zamia magnifica Pérez-Farrera, Gutiérrez-Ortega, and Calonje sp. nov. (Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9 and Figure 10).

Figure 4.
(A) Zamia magnifica sp. nov. in habitat. (B) Leaves of Z. magnifica (next to first author) are among the largest in the genus.

Figure 5.
(A) Leaf of adult Zamia magnifica. (B) Details of base of petiole.

Figure 6.
(A) Close up view of leaflet of Zamia magnifica. Note the visible veins. (B) Trunk of juvenile. (C) Seedling with eophyll.

Figure 7.
Pollen cone of Zamia magnifica. (A) Fertile region of strobilus (without peduncle). (B) Distal portion of strobilus. (C) Cross section showing microsporophylls. (D) Microsporophylls, abaxial (top) and adaxial (bottom) surfaces.

Figure 8.
Seed cone of Zamia magnifica. (A) Entire strobilus with peduncle. (B) Apex. (C) Apical view. (D) Basal view.

Figure 9.
Zamia magnifica in habitat. (A) Emergent leaf showing inflexed ptyxis. (B) Emergent leaf expanding, showing dense cover of white tomentum. (C) Immature leaf color change. (D) Mature leaf. (E) Articulation of leaflets and tomentose rachis. (F) Median leaflet.

Figure 10.
Zamia magnifica in habitat. (A) Mature leaf hanging from cliff. (B) Close up view of immature leaf. (C) Rupicolous habit of Z. magnifica.
Holotype.—MEXICO. Oaxaca, Sierra Norte, Santiago Tuxtepec, 80 m, Pérez-Farrera 4285, and Mauricio G. Martínez-Mtz, Diego Villar Morales, male, 3 August 2022. (HEM!). Isotypes: (XAL!, MEXU!).
Zamia magnifica is distinguished from its congeners by its rupicolous habit, pendent leaves bearing leaflets that are densely tomentose and pink to caramel in color when emerging, and has broad oblong leaflets that are coriaceous in texture with few non-prominent denticulations, and ovulate strobili with short (<4 cm) peduncles.
Additional specimens examined.—MEXICO. Oaxaca, Sierra Norte, Santiago Tuxtepec, 80 m a.s.l., Pérez-Farrera 4286, and Mauricio G. Martínez-Mtz, Diego Villar Morales, male, 3 August 2022. (HEM!); Mauricio G. Martínez-Mtz 9, 10, 11, 12 and Pérez-Farrera, Diego Villar Morales, 3 August 2022. (HEM!).
Description. Stem pachycaulous, hypogeous, erect, or decumbent, solitary to profusely branching, 10–38 cm in height, 10–17 cm in diameter. Cataphylls persistent, brown, and densely tomentose at emergence. Leaves 3–4 per stem, pendent, 160.8–324.0 cm long, 38.7–60.0 cm wide, densely tomentose and pink in the emergence changing to caramel color, transitioning finally to dark green and sparsely tomentose at maturity. Petiole terete, 57.3–87.0 cm long, sparsely armed with very few minute prickles diminishing from the petiole to rachis. Rachis terete, 93–206 cm, unarmed or sparsely armed with few minute prickles in the proximal third. Leaflets 10–17 pairs, oblong to oblanceolate, subcoriaceous, alternate in the basal part of the rachis, subopposite in the median, opposite in the apical part of the rachis, apex acute to rounded, crenulately toothed on the distal fourth, asymmetric or symmetric, base attenuate; median leaflets 29.8–47.7 × 6.0–11.1 cm, inter-leaflet distance 5.0–13.1 cm; articulations green, 0.8–1.7 cm wide. Eophylls with two leaflets. Pollen strobili 1–3, cylindrical, erect, 10.75–15.00 × 1.90–2.33 cm, light green turning tan to light brown or light orange color at maturity; peduncle tomentose, 3.2–5.5 × 1.2–1.5 cm. Microsporophylls spirally arranged in 18–21 orthostichies of 32–37 sporophylls each, cuneiform, distal face 5.7–7.9 mm long, 4.0–5.6 mm wide, abaxial surface bearing 18–24 microsporangia aggregated into a single group. Ovulate strobili solitary, cylindrical, erect, 9.1–15.8 cm, 7.2–7.8 cm in diameter, yellowish at maturity; apex acute, 1.6–2.0 cm long, peduncle tomentose, light yellow at maturity, 9.1–10 × 1.4–2.7 cm. Megasporophylls arranged in 7–8 orthostichies of 9–10 sporophylls each, cuneiform, peltate, distal end hexagonal truncate, 24.8–37 mm short axis, 23.2–32.5 mm wide axis, distal face brown pubescent, yellowish when immature, green at maturity. Seeds ovate, sarcotesta reddish orange, 18.6–23.5 mm long, 12.5–15.2 mm in diameter.
Habitat description. The vegetation type where this cycad is found is tropical rainforest, according to the classification of Breedlove [29]. Zamia magnifica is sympatric with Dioon spinulosum. Both species occur in karst tropical rainforest and gallery forest in lowland forest in the Sierra de Juarez, Oaxaca dominated by: Swietenia macrophylla King, Inga sp., Cedrela odorata L., Trophis sp., Bursera simaruba (L.) Sarg., Oreopanax sp., Dendropanax sp., Attalea butyracea (Mutis ex L. f.) Wess. Boer, Chamaedorea elatior Mart., C. tepejilote Liebm., C. oblongata Mart., C. pinnatifrons (Jacq.) Oerst., C. metallica O.F. Cook ex H.E. Moore, C. sartorii Liebm., Cecropia peltata L., Bactris mexicana Mart., Cnidoscolus sp., Piper sp., Heliconia sp., epiphytes and hemiepiphytes as Monstera sp., Philodendron sp., Syngonium neglectum Schott, Tillandsia sp., Maxillaria sp., Anthurium schlechtendalii Kunth, Spathiphyllum sp. The region is characterized by sedimentary and metamorphic rocks as well as lephosols, cambisols, fluvisol, and humic acrisol soils. There are also sedimentary rocks from the Cretaceous period of the Mesozoic era, generally on abruptly steep slopes of about 40–45° and it is found within the range of 80–100 m a.s.l. [30,31].
Etymology. The epithet ‘magnifica’, derived from the Latin magnificus (‘magnificent’), refers to the strikingly beautiful aspect of the species due to its large and spectacular pendent leaves.
Uses. The local people mentioned that they use the seeds of Z. magnifica as an insecticide. They grind the seeds and add sugar or honey and deposit the mix in an open recipient. Flies eventually eat from the mix and die due to the toxic compounds.
Distribution and conservation status. The species should be classified as Critically Endangered (CR) according to the IUCN (2001), due to its restricted distribution (La Chinantla, Sierra Norte, Oaxaca) and its limited number of populations (only one population) with around 2000 individuals (considering seedlings, juveniles, and adults). The area where the population occurs is approximately 4477 ha (44.8 km2) in size, and plants are actively protected by the local people, making studies and collection of them impossible without their consent and permission. However, much of the surrounding habitat is being transformed into pastures for raising cattle and for coffee and corn cultivation. As is the case with many cycad species in Mexico, its conservation both ex situ and in situ is urgent.
4. Discussion
Zamia magnifica occurs in one locality in the La Chinantla region, in Sierra Norte, Oaxaca. La Chinantla region is an area that has been suggested to be a Neogene refuge [32], where a high floristic richness and great endemism of ancient and rare taxa occur [33,34]. About 1021 species have been reported in this locality [33,35,36]. The plant communities that are below 1000 m altitude in the Sierra Norte have been greatly altered due to human activities [33], being converted to grazing areas or pastures for cattle, and cultivation areas for corn and coffee. However, some hills consist of rich karstic tropical forests that contain great levels of endemism. Other notable species such as Beaucarnea recurvate, Chamaedorea metallica, Ceratozamia whitelockiana, C. aurantiaca, Dioon spinulosum, D. rzedowskii, Gaussia gomez-pompae, and Licaria chinanteca also live in these forests.
Zamia magnifica is one of four obligate cliff-dwelling Zamia species in Mesoamerica, the others being Z. cremnophila Vovides, Schutzman, and Dehgan from Mexico, Z sandovalii C.Nelson from Honduras, and Z. meermanii from Belize. The species is easily distinguished from Z. cremnophila and Z. sandovalii by its leaflets alone, which are oblong to oblanceolate and crenulately toothed at the apex, compared to those of the other two species which are lanceolate and prominently toothed at the distal half. Among the rupicolous Mesoamerican species, Z. magnifica most closely resembles the Belizean species Z. meermanii, as both species share pendent leaves that are densely tomentose and colorful at emergence, bearing obovate to oblanceolate leaflets that are inconspicuously toothed at the apex. However, the leaflets of Z. meermanii are concave and strongly coriaceous in texture, whereas those of Z. magnifica are distinctly convex and moderately coriaceous. The new leaves of Z. magnifica are whitish pink, and the color changes to caramel with white to tan tomentum. The leaves of Z. meermanii are cream-colored, and covered with saffron-orange tomentum (Table 5). Morphometrically, Z. magnifica can be distinguished from Z. meermanii in 11 out of 12 traits we evaluated (all traits except the number of leaves) (Table 2). Zamia magnifica attains much larger dimensions overall than Z. meermanii, typically bearing much longer leaves (142–290 cm vs. 71–147 cm) and median leaflets that are much longer (29–48 cm vs. 18–32.0 cm) and broader (6–11 vs. 3.4–7.5 cm). Furthermore, Z. magnifica has microsporophylls with 18–24 microsporangia aggregated into a single group, whereas Z. meermanii has 22–28 microsporangia occurring in two distinctly separate groups. In addition, the stems of Z. meermanii are typically solitary, whereas in Z. magnifica they can be profusely branching on older individuals (see Figure 4A).
Table 5.
Morphological variation among Zamia magnifica and species affine: (Z. furfuracea and Z. meermanii).
Zamia magnifica also shares some morphological resemblance to the Veracruz endemic species Z. furfuracea, as both species have leaves that are densely tomentose at emergence, bearing leaflets that are similar in shape (oblong to oblanceolate), and are inconspicuously toothed at the apex. However, Z. furfuracea is a terrestrial species (vs. rupicolous) with spreading (vs. pendent) leaves, the emerging leaves in Z. furfuracea are ochre to light brown (vs. pink), and seed strobili with longer and thinner peduncles (Table 5) (14–16 vs. 9.1–10 cm). Eleven out of 12 vegetative traits were significantly differentiated between the two species (all traits except the number of leaves) (Table 2). Zamia furfuracea is a much smaller plant overall, having much smaller leaves (48–66 cm vs. 142–290 cm) and leaflets that are much shorter (10.5–16.5 vs. 29–48 cm) than Z. magnifica.
Definitely, Z. magnifica is the Mexican Zamia species with the largest leaves (up to 290 cm). The second Mexican Zamia would be Z. splendens Schutzman, with leaves of around 2 m long. Some plants of Z. variegata Warsz. in Belize may also produce large leaves (at more than 2.5 m long), especially in plants living in areas with frequent flooding (personal obs.) or in cultivation [1]. Furthermore, it is notable that Z. magnifica produces some of the largest seeds among the Mexican Zamia; we found seeds 1.9–2.4 cm long and 1.1–1.5 cm wide, whereas the Mexican Zamia with the largest seeds is Z. soconuscensis (2.2–2.6 cm long, 1.4–1.9 cm wide).
Recent phylogenetic work in the genus Zamia [5] has helped clarify the relationships between the Mesoamerican species of Zamia, with four major groups evident: (i) The Fischeri clade, being sister to the rest of the mainland American species (excluding Z. integrifolia L.f.), consists of the three species from northeastern Mexico: Z. fischeri Miq., Z. vazquezii D.W.Stev., Sabato and De Luca, and Z. inermis Vovides, J.D.Rees and Vázq.Torres. (ii) The Furfuracea subclade, being a broadly distributed group (Mexico, Guatemala, Belize, and Honduras), consists of seven species: Z. herrerae S.Calderón and Standl., Z. paucijuga Wieland, Z. prasina W.Bull, Z. variegata, Z. spartea A.DC., Z. furfuracea, and Z. loddigesii Miq. (iii) The Purpurea clade consists of six species that are associated mostly with the Northern Area of the Chiapas Highlands biogeographic province [3]: Z. lacandona Schutzman and Vovides, Z. grijalvensis Pérez-Farr., Vovides and Mart.-Camilo, Z. purpurea Vovides, J.D.Rees and Vázq.Torres, Z. cremnophila, Z. splendens, and most likely Z. katzeriana (Regel) E.Rettig (see below). (iv) The Tuerckheimii subclade is restricted to evergreen tropical rainforests in Guatemala, Belize, and Honduras, and consists of seven species: Z. decumbens Calonje, Meerman, M.P.Griff. and Hoese, Z. onan-reyesii C.Nelson and Sandoval, Z. tuerckheimii Donn.Sm., Z. meermanii, Z. sandovalii, Z. standleyi Schutzman, and most likely Z. oreillyi C.Nelson (see below). Calonje et al. [5] did not include Z. katzeriana or Z. oreillyi C.Nelson in their molecular phylogenetic analysis, but the former is believed to belong to the Purpurea clade [23], and the latter, based on its morphological characteristics and geographical distribution, is likely in the Tuerckheimii subclade. Although Z. magnifica has yet to be included in phylogenetic analyses, we hypothesize the species most likely belongs to the Furfuracea subclade due to its geographic distribution and morphology. Zamia magnifica shares a similar leaflet morphology to Z. furfuracea (member of the Furfuracea clade), as well as the densely tomentose new leaves that characterize that group. Geographically, Z. magnifica occurs within the geographic distribution range of the Furfuracea clade, and because there is a strong congruence between geographic distribution and phylogenetic relationships in Zamia [5], the distribution is a piece of evidence that favors the idea of Z. magnifica being in the Furfuracea clade. Based on the distribution ranges of the Zamia clades, an alternative might be that Z. magnifica belongs to the Purpurea clade. However, the Purpurea clade is limited to the east of the Isthmus of Tehuantepec (whereas Z. magnifica occurs to the west). This geographic disjunction is notable, as the Isthmus of Tehuantepec has been recognized as a formidable geographic barrier affecting the distribution and diversification of Dioon [37] and Ceratozamia [38], which disfavors this alternative hypothesis of Z. magnifica in the Purpurea clade. Future phylogeographic studies will contribute to untangle the relationships among Zamia species in these geographic regions.
The distribution of Z. magnifica is restricted to certain cliffs where rocks are exposed, and plants are usually very difficult to access. We hypothesize that the cliff-dwelling habit in Z. magnifica might have evolved from a terrestrial habit ancestor, and originated locally in La Chinantla as a response to avoid competition with other terrestrial species. The same might be true for the other cliff-dwelling Zamia species, implying that the habit has multiple origins in the genus, and that each origin might be triggered by distinct or similar local biotic or abiotic factors. However, little is known about this aspect. For example, what are the physiological and anatomical features that may enable these possible cases of adaptation to the cliff-dwelling habit? A recent study on Zamia found little correlation between leaflet anatomical trait variation and phylogeny or macro-environmental variation [39], leaving open the possibility that the effect of micro-environmental variation on selection or biotic factors such as competition are possible causes. In the case of Z. magnifica, it is possible that rodents or other small mammals or birds feeding on the sarcotesta deposit the seeds in the fissures of rocks where plants grow, but which are the exact species that disperse the seeds is still unknown. Nevertheless, we expect that the discovery of Z. magnifica may encourage more researchers to clarify the evolutionary and ecological consequences of the cliff-dwelling habit of cycads.
5. Conclusions
This research presented Z. magnifica, a species for which there are no previous records in herbaria, as far as we know. The discovery of Z. magnifica suggests that the botanical explorations in southern Mexico are still promising, and expeditions to remote places may lead to an even a better comprehension of the cycad diversity. Understanding the full diversity may help us to disentangle the evolutionary history of these plants and contribute to their effective conservation.
Author Contributions
Conceptualization, M.A.P.-F.; methodology, M.A.P.-F., J.S.G.-O., M.G.M.-M. and M.C.; software, M.A.P.-F. and J.S.G.-O.; validation, M.A.P.-F., J.S.G.-O., M.G.M.-M. and M.C.; formal analysis, M.A.P.-F. and J.S.G.-O.; investigation, M.A.P.-F., J.S.G.-O., M.G.M.-M. and M.C.; resources, M.A.P.-F.; data curation, J.S.G.-O.; writing—original draft preparation, M.A.P.-F., J.S.G.-O. and M.C.; writing—review and editing, M.A.P.-F., J.S.G.-O. and M.C.; visualization, J.S.G.-O.; supervision, M.A.P.-F.; project administration, M.A.P.-F.; funding acquisition, M.A.P.-F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Montgomery Botanical Center’s Plant Exploration Fund.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We appreciate the comments that we received form four anonymous reviewers. This work was supported by Montgomery Botanical Center’s Plant Exploration Fund and anonymous donors to MAPF. We thank Diego Villar Morales for his assistance in the field in Oaxaca, Mexico. Fieldwork with Z. meermanii in Belize was funded by the Association of Zoological Horticulture (AZH) and made possible by Jan Meerman, who introduced MC to the species, participated in the fieldwork, and made possible its description. The Belize Forest Department granted the scientific research and collecting permit (CD/60/3/08(45)) supporting research on Belizean Zamiaceae.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Norstog, K.; Nicholls, T.J. The Biology of the Cycads; Cornell University Press: Ithaca, NY, USA, 1997; p. 384. [Google Scholar]
- Calonje, M.; Vovides, A.P.; Gutiérrez-Ortega, J.S. An overview of cycadales in Mesoamerica and the Caribbean: Biology, distribution, and conservation. In Under the Shade of Thipaak: The Ethnoecology of Cycads in Mesoamerica and the Caribbean; Carrasco, M.D., Cibrián Jaramillo, A., Bonta, M., Englehardt, J.D., Eds.; University Press of Florida: Gainesville, FL, USA, 2022; pp. 27–61. [Google Scholar]
- Morrone, J.J.; Escalante, T.; Rodríguez-Tapia, G.; Carmona, A.; Arana, M.; Mercado-Gómez, J.D. Biogeographic regionalization of the Neotropical region: New map and shapefile. An. Acad. Bras. Cienc. 2022, 94, e20211167. [Google Scholar] [CrossRef] [PubMed]
- Calonje, M.; Stevenson, D.W.; Osborne, R. The World List of Cycads, Online Edition. 2023. Available online: http://cycadlist.org (accessed on 23 April 2023).
- Calonje, M.; Meerow, A.W.; Griffith, M.P.; Salas-Leiva, D.; Vovides, A.P.; Coiro, M.; Francisco-Ortega, J. A time-calibrated species tree phylogeny of the New World cycad genus Zamia L. (Zamiaceae, Cycadales). Int. J. Plant Sci. 2019, 180, 286–314. [Google Scholar] [CrossRef]
- Calonje, M.; Stevenson, D.W.; Calonje, C.; Ramos, Y.A.; Lindstrom, A. A new species of Zamia from Choco, Colombia (Cycadales, Zamiaceae). Brittonia 2010, 62, 80–85. [Google Scholar] [CrossRef]
- Calonje, M.; Esquivel, H.E.; Stevenson, D.; Calonje, C.; Pava, D. A new arborescent species of Zamia from the Central Cordillera of Tolima, Colombia (Cycadales, Zamiaceae), with comments on the Z. poeppigiana species complex. Brittonia 2011, 63, 442–451. [Google Scholar] [CrossRef]
- Calonje, M.; Esquivel, H.E.; Morales, G.; Mora-Lizcano, Y.A.; Stevenson, D. A new arborescent species of Zamia (Cycadales, Zamiaceae) from the department of Huila, Eastern Cordillera of Colombia. Caldasia 2012, 34, 8. [Google Scholar]
- Calonje, M.; Betancur, J.; Lindstrom, A.; Lopez-Gallego, C.; Castro, J.; Castro, C.; Miguel Niño, S.; Barraez, D.C. Zamia orinoquiensis (Zamiaceae, Cycadales), a new species from the western Orinoquía region of Colombia. Phytotaxa 2022, 556, 119–135. [Google Scholar] [CrossRef]
- Lindstrom, A.J.; Idarraga, A. Zamia incognita (Zamiaceae): The exciting discovery of a new gymnosperm from Colombia. Phytotaxa 2009, 2, 6. [Google Scholar] [CrossRef]
- Segalla, R.; Calonje, M. Zamia brasiliensis, a new species of Zamia (Zamiaceae, Cycadales) from Mato Grosso and Rondônia, Brazil. Phytotaxa 2019, 404, 1–11. [Google Scholar] [CrossRef]
- Stevenson, D.W.; López, D.C.; Arboleda, N.C. A new Zamia (Zamiaceae) from Colombia. Brittonia 2018, 70, 364–368. [Google Scholar] [CrossRef]
- Calonje, M.; Taylor-Blake, A.S.; Stevenson, D.; Holzman, G.; Ramos, Y.A. Zamia lindleyi: A misunderstood species from the highlands of western Panama. Mem. N. Y. Bot. Gard. 2012, 106, 419–437. [Google Scholar]
- Calonje, M.; Morales, G.; López-Gallego, C.; Roldán, F. A taxonomic revision of Zamia montana and Zamia oligodonta, with notes on their conservation status. Phytotaxa 2015, 192, 279–289. [Google Scholar] [CrossRef]
- Lindstrom, A.J. Typification of some species names in Zamia L. (Zamiaceae), with an assessment of the status of Chigua D. Stev. Taxon 2009, 58, 265–270. [Google Scholar] [CrossRef]
- Nicolalde-Morejón, F.; Vovides, A.P.; Stevenson, D.W. Taxonomic revision of Zamia in mega-Mexico. Brittonia 2009, 61, 301–335. [Google Scholar] [CrossRef]
- Calonje, M. A new cliff-dwelling species of Zamia (Zamiaceae) from Belize. J. Bot. Res. Inst. Texas 2009, 3, 23–29. [Google Scholar]
- Calonje, M.; Meerman, J.; Griffith, M.P.; Hoese, G. A new species of Zamia (Zamiaceae) from the Maya Mountains of Belize. J. Bot. Res. Inst. Texas 2009, 3, 31–41. [Google Scholar]
- Pérez-Farrera, M.Á.; Vovides, A.P.; Martínez-Camilo, R.; Martínez-Meléndez, N.; Gómez-Domínguez, H.; Galicia-Castellanos, S. Zamia grijalvensis sp. nov. (Zamiaceae, Cycadales) from Chiapas, Mexico with notes on hybridization and karyology. Nord. J. Bot. 2012, 30, 565–570. [Google Scholar] [CrossRef]
- Nicolalde-Morejón, F.; Martínez-Domínguez, L.; Stevenson, D.W.; Vergara-Silva, F. Disentangling the identity of Zamia from Mexican Pacific seaboard, with a description of a new species. Nord. J. Bot. 2019, 37. [Google Scholar] [CrossRef]
- Calonje, M.; Meerman, J. What is Zamia prasina (Zamiaceae: Cycadales)? J. Bot. Res. Inst. Texas 2009, 3, 43–49. [Google Scholar]
- Pérez-Farrera, M.Á.; Vovides, A.; Ruíz Castillejos, C.; Galicia, S.; Cibrian-Jaramillo, A.; López, S. Anatomy and morphology suggest a hybrid origin of Zamia katzeriana (Zamiaceae). Phytotaxa 2016, 270, 161–181. [Google Scholar] [CrossRef]
- Gutiérrez-Ortega, J.S.; Pérez-Farrera, M.A.; López, S.; Vovides, A.P. Demographic history and species delimitation of three Zamia species (Zamiaceae) in south-eastern Mexico: Z. katzeriana is not a product of hybridization. Bot. J. Linn. Soc. 2023, boac062. [Google Scholar] [CrossRef]
- Pérez-Farrera, M.A.; Vovides, A.P.; Martinez-Camilo, R.; Martinez Meléndez, N.; Iglesias, C. A reassessment of the Ceratozamia miqueliana species complex (Zamiaceae) of southeastern Mexico, with comments on species relationships. System. Biodiv. 2009, 7, 433–443. [Google Scholar] [CrossRef]
- Pérez-Farrera, M.A.; Vovides, A.P.; Avendaño, S. Morphology and leaflet anatomy of the Ceratozamia norstogii species complex (Zamiaceae, Cycadales). Int. J. Plant Sci. 2014, 175, 110–121. [Google Scholar] [CrossRef]
- Gutiérrez-Ortega, J.S.; Jiménez-Cedillo, K.; Pérez-Farrera, M.A.; Martínez, J.F.; Molina-Freaner, F.; Watano, Y.; Kajita, T. Species definition of Dioon sonorense (Zamiaceae, Cycadales), and description of D. vovidesii, a new cycad species from northwestern Mexico. Phytotaxa 2018, 369, 107–114. [Google Scholar] [CrossRef]
- Gutiérrez-Ortega, J.S.; Pérez-Farrera, M.A.; Vovides, A.P.; Chávez-Cortázar, A.; López, S.; Santos-Hernández, N.G.; Ruíz-Roblero, S.K. Ceratozamia sanchezae (Zamiaceae): A new cycad species from Chiapas Highlands (Mexico). Phytotaxa 2021, 500, 201–216. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Breedlove, D.E. Flora of Chiapas. Part 1. Introduction to the Flora of Chiapas; California Academy of Sciences: San Francisco, CA, USA, 1981; p. 35. [Google Scholar]
- Ferrusquia-Villafranca, I. La Geología de México: Una Synopsis. In La Biodiversidad de México; Ramamoorthy, T.P., Bye, R., Lot, A., Fa, J., Eds.; UNAM, Instituto de Biología. D.F.: Mexico City, Mexico, 1998; pp. 1–107. [Google Scholar]
- INEGI [Instituto Nacional de Estadística y Geografía]. Prontuario de Información Geográfica Municipal de los Estados Unidos Mexicanos. Available online: https://www.inegi.org.mx/contenidos/app/mexicocifras/datos_geograficos/07/07115.pdf (accessed on 4 May 2023).
- Toledo, V.M. Pleistocene Changes of Vegetation in Tropical Mexico. In Biological Diversification in the Tropics; Prance, G.T., Ed.; Columbia University Press: New York, NY, USA, 1982; pp. 93–111. [Google Scholar]
- Meave, J.A.; Rincón-Gutiérrez, A.; Ibarra-Manríquez, G.; Gallardo-Hernández, C.; Romero-Romero, M.A. Checklist of the vascular flora of a portion of the hyper-humid region of La Chinantla, Northern Oaxaca Range, Mexico. Bot. Sci. 2017, 95, 722–759. [Google Scholar] [CrossRef]
- Villaseñor, J.L.; Meave, J.A. Floristics in Mexico today: Insights into a better understanding of biodiversity in a megadiverse country. Bot. Sci. 2022, 100, 14–33. [Google Scholar] [CrossRef]
- Rzedowski, J. Vegetación de México. 1ra Edición digital, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Limusa, Mexico. 2006. Available online: https://www.biodiversidad.gob.mx/publicaciones/librosDig/pdf/VegetacionMx_Cont.pdf (accessed on 4 May 2023).
- Rzedowski, J.; Palacios-Chávez, R. El bosque de Engelhardtia (Oreomunnea) mexicana en la región de la Chinantla (Oaxaca, México). Una reliquia del Cenozoico. Bot. Sci. 1977, 36, 93–127. [Google Scholar] [CrossRef]
- Gutiérrez-Ortega, J.S.; Salinas-Rodríguez, M.M.; Ito, T.; Pérez-Farrera, M.A.; Vovides, A.P.; Martínez, J.F.; Molina-Freaner, F.; Hernández-López, A.; Kawaguchi, L.; Nagano, A.J.; et al. Niche conservatism promotes speciation in cycads: The case of Dioon merolae (Zamiaceae) in Mexico. New Phytol. 2020, 227, 1872–1884. [Google Scholar] [CrossRef]
- Pérez-Farrera, M.A.; Gutiérrez-Ortega, J.S.; Vovides, A.P.; Calonje, M.; Díaz-Jiménez, P. Ceratozamia dominguezii (Zamiaceae): A New Cycad Species from Southeastern Mexico. Taxonomy 2021, 1, 345–359. [Google Scholar] [CrossRef]
- Glos, R.A.; Salzman, S.; Calonje, M.; Vovides, A.P.; Coiro, M.; Gandolfo, M.A.; Specht, C.D. Leaflet anatomical diversity in Zamia (Cycadales: Zamiaceae) shows little correlation with phylogeny and climate. Bot. Rev. 2022, 88, 437–452. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).