1. Introduction
Seed shape diversity observed in
Silene has long been recognized as a valuable source of information for infrageneric classification. Seed morphological characters have been used with diverse applications in the taxonomy of this genus [
1,
2,
3,
4,
5,
6,
7,
8,
9,
10,
11]. In relation to general seed shape, the description is traditionally based on adjectives such as
reniformia [
1,
2], reniform, reniform-circular [
3,
4,
5,
6,
7,
8]; half-rounded, flabellate [
7]; round-reniform, symmetrical-reniform, cordate-reniform, asymmetrical-reniform, semicircle-reniform [
8]; and ovate or winged [
9]. Recently, a relationship between the surface structure and the overall outline seed morphology has been described [
12,
13]. Based on the seed surface outline of 52 species of
Silene and two closely related genera (
Atocion and
Viscaria), a classification in four groups was proposed (smooth, rugose, echinate, and papillose) [
13]. This seed classification showed a certain association with the taxonomic treatment of the currently defined subgenera. Hence, the group of smooth seeds was mostly composed of species of
S. subg.
Silene while the majority of the echinate seeds corresponded to a species of
S. subg.
Behenantha.
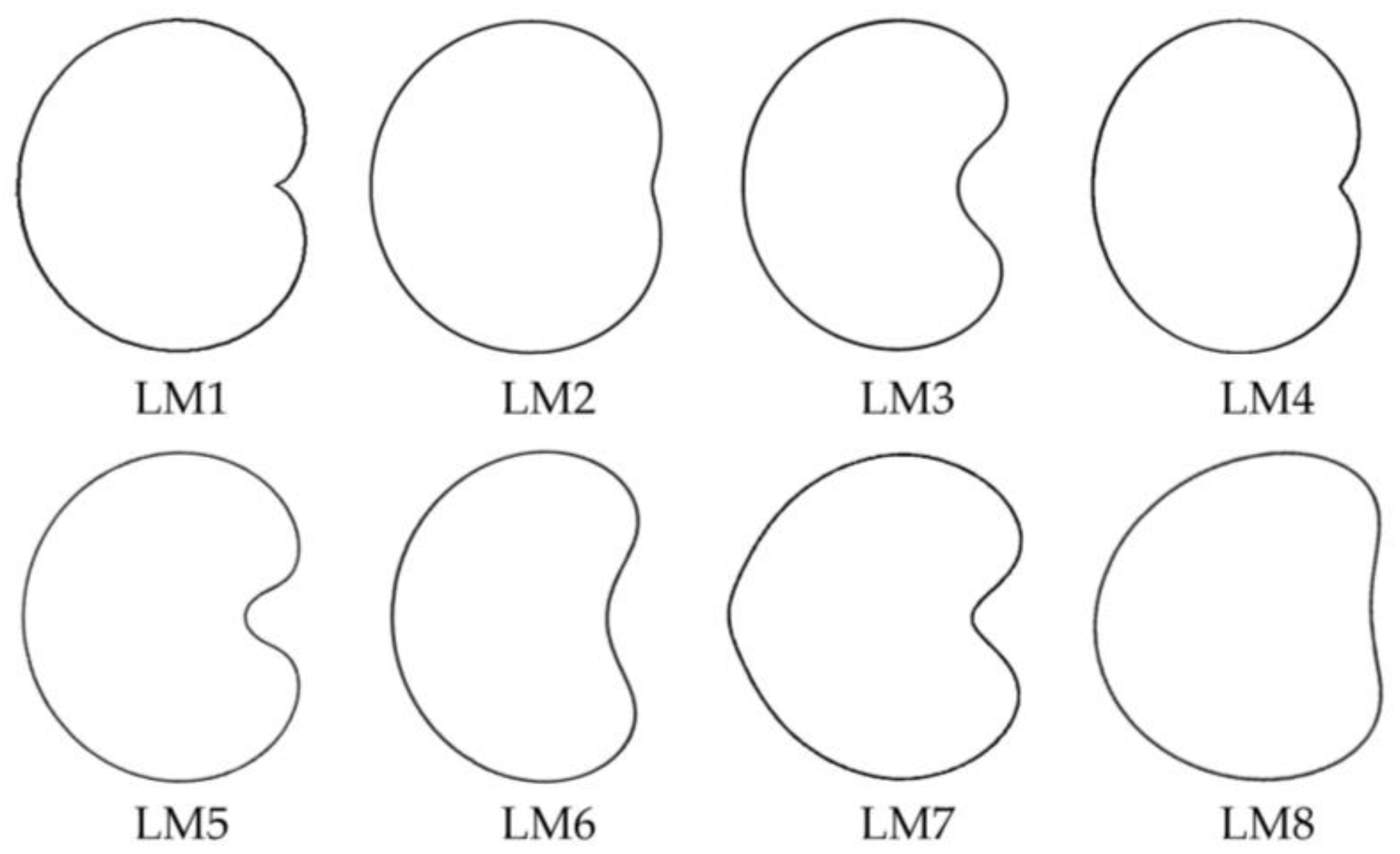
As a new morphological approach, we have described the lateral and dorsal views of
Silene seeds comparing the seed contour with algebraically defined geometric figures, such as the cardioid, modified cardioid, or diverse ellipses. We provided a
J index, a numerical value indicating the percentage of similarity between a defined seed silhouette and a given geometric figure, taken as a model [
14,
15,
16,
17,
18]. The comparison of lateral seed views in geometric models was applied for the first time in
Silene for 21 species with the basic model LM1 (the cardioid) and the values of the
J index were over 90 in 11 species of
Silene subg.
Behenantha (
S. acutifolia Link ex Rohrb.,
S. conica L.,
S. diclinis (Lag.) M.Laínz,
S. dioica (L.) Clairv.,
S. latifolia Poir.,
S. noctiflora L.,
S. pendula L.,
S. uniflora Roth,
S. viscosa (L.) Pers.,
S. vulgaris (Moench) Garcke and
S. zawadskii Herbich), and 7 species of
S. subg.
Silene (
S. gallica L.,
S. italica (L.) Pers.,
S. nutans L.,
S. otites (L.) Wibel,
S. portensis L.,
S. saxifraga L., and
S. vivianii Steud.) [
14]. Nevertheless, seeds of
S. diclinis and
S. latifolia had higher
J index values in the additional models LM2 and LM4 (derived from the cardioid, LM1), than in the cardioid model. Seeds of
S. gallica had the highest value in another geometric model LM3 [
14].
The research continued with the description of new models for the lateral view (LM5 to LM8) based on the analysis of 20
Silene species. From these 20 species, 16 (
S. coutinhoi Rothm. and P.Silva,
S. crassipes Fenzl.,
S. disticha Willd.,
S. diversifolia Otth.,
S. foetida Link ex Spreng.,
S. littorea Brot.,
S. micropetala Lag.,
S. muscipula L.,
S. nicaeensis All.,
S. nocturna L.,
S. portensis L.,
S. scabriflora Brot.,
S. sclerocarpa Dufour,
S. stricta L.,
S. tridentata Desf., and
S. vivianii) were analyzed for the first time, extending the analysis to a total of 37
Silene species [
15]. This work revealed the potential existence of four defined seed groups according to the aspect ratio in their dorsal views. The new geometric models gave the best results with
S. diversifolia and
S. tridentata (LM5),
S. coutinhoi (LM7) and
S. nicaeensis,
S. portensis, and
S. scabriflora (LM8). In addition, a preliminary correspondence was found between lateral models and the convexity values in the dorsal views. Those convex seeds in their dorsal views adjusted well to models LM2 and LM4, while seeds that presented concavities in their dorsal views adjusted better to other lateral models (LM3, LM5, and LM6).
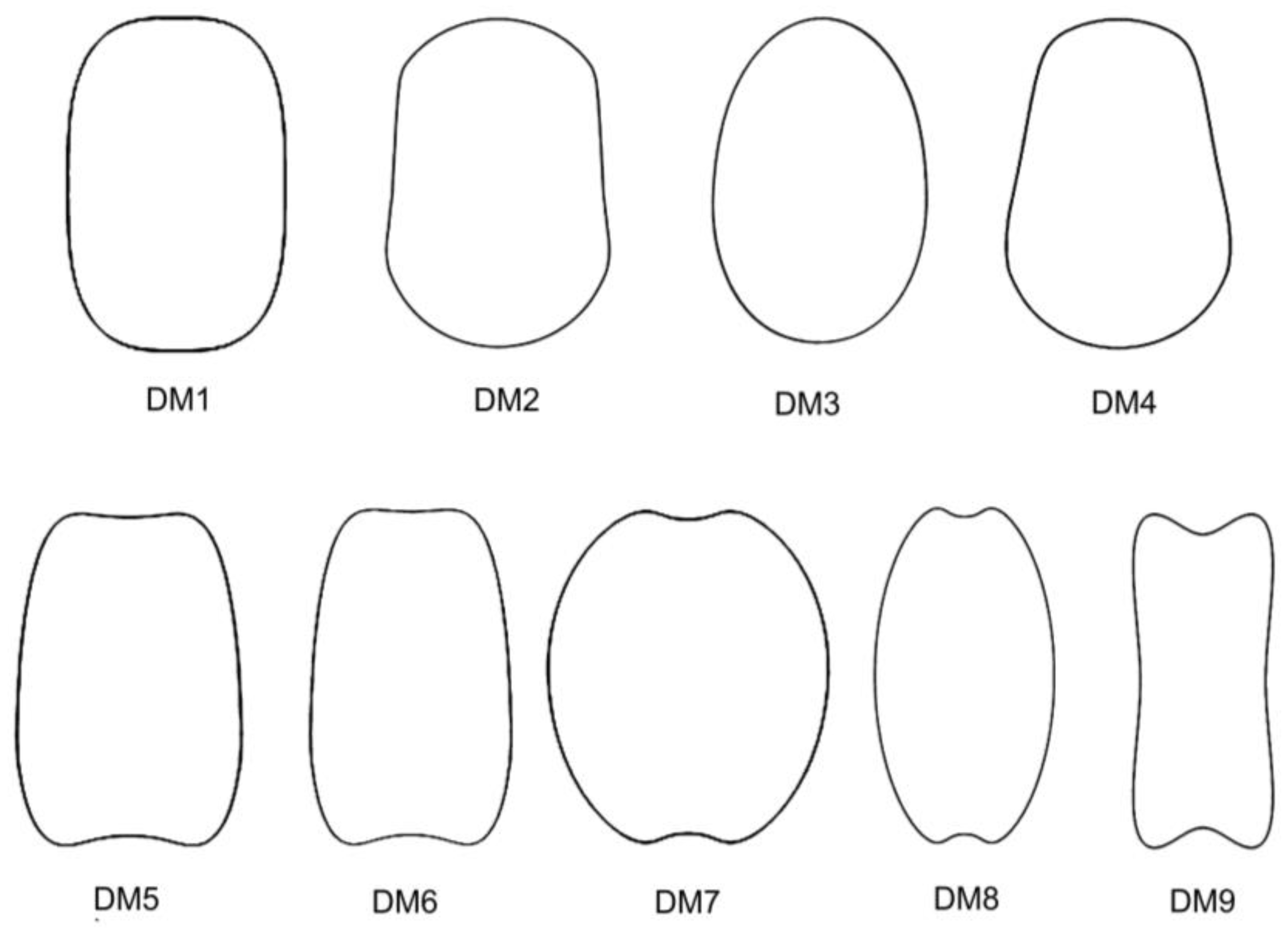
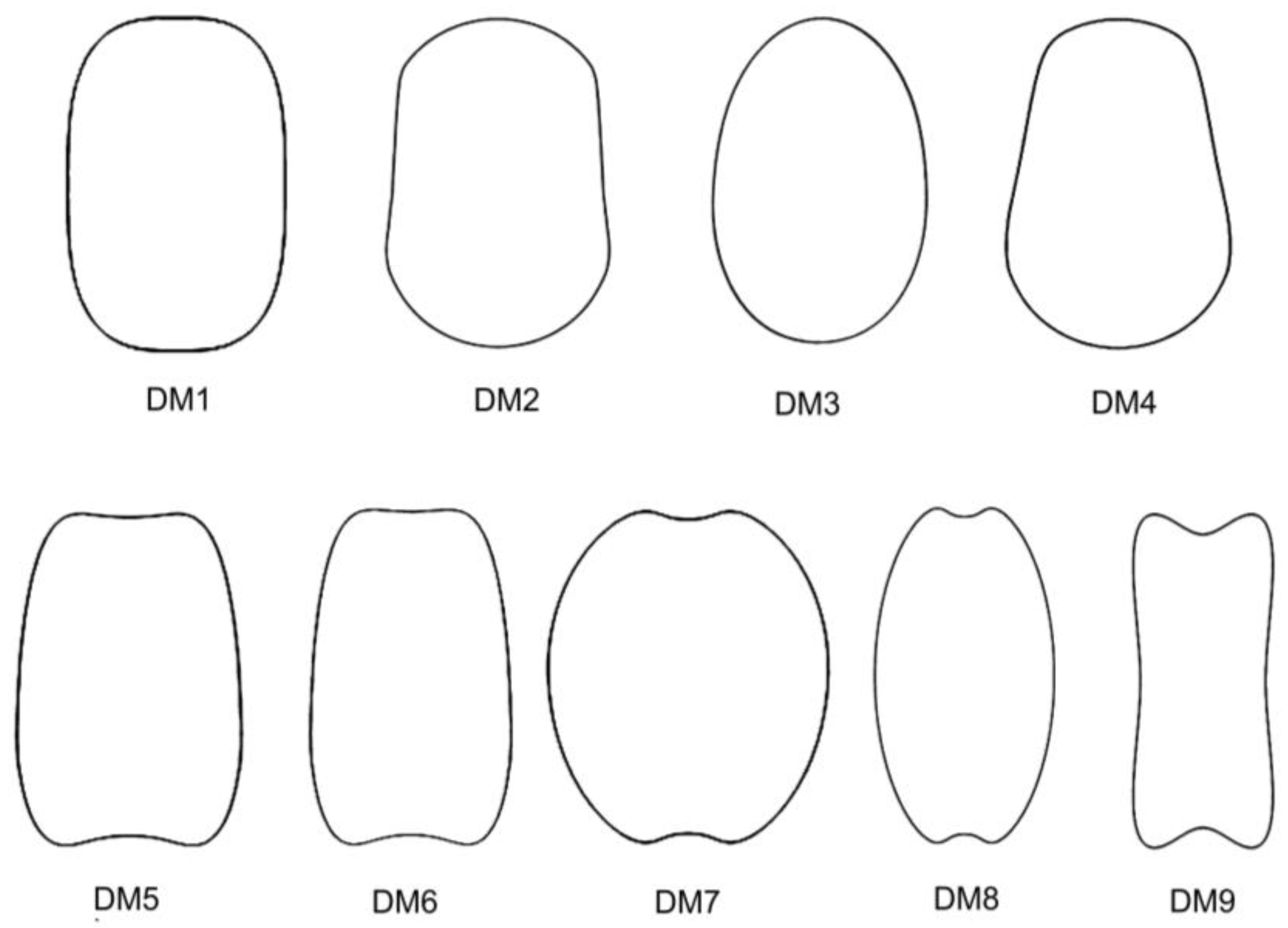
A new approach was recently conducted based on dorsal views and the relationship between their morphology and the geometric models. Rodríguez-Lorenzo et al. [
16] reported new models for dorsal views of seeds, together with new data from the
J index for the lateral views of the species
S. inaperta L.,
S. pseudoatocion Desf., and
S. ramosissima Desf. in the lateral geometric model LM1. Dorsal seed views adjusted to nine models (DM1–DM9), which corresponded to convex (DM1–DM4) and non-convex (DM5–DM9) model types. The species related to convex models were
S. diclinis,
S. dioica,
S. foetida,
S. latifolia, and
S. vulgaris; those with non-convex models were
S. conica,
S. coutinhoi,
S. inaperta, and
S. ramosissima.
Based on the data set out above, the description and quantification of seed shape with geometric models revealed a certain level of differentiation among species, and this morphological tool constitutes a promising technique for the taxonomy of species, especially for
Silene. An example of the description and taxonomical use of lateral and dorsal models was reported by Martin-Gómez et al. [
17] for the
S. mollissima (L). Pers. aggregate. The images of seeds of this group of species adjusted to three lateral (LM1, LM4, and LM5) and two dorsal models (DM5 and DM6) [
17]. The authors reported that the degree of similarity of the seeds to the model LM5 revealed the identification of two groups of species with different geographic origin. This work highlighted the stability of seed shape measured by comparison with a geometric models in contrast to other common morphological measurements (e.g., seed size).
The objectives of the present work were (i) to apply a seed description based on geometric models to non-previously analyzed Silene species, (ii) to identify the differences between these species, and (iii) to establish a correspondence between the groups based on their surface and geometric models describing the overall shape in the lateral and dorsal views of these Silene species. We hypothesize that the obtained information may provide new useful knowledge to understand the relationships among species as well as providing new comparative information for their characterization and classification using this morphological approach.
4. Discussion
The similarity of lateral views of
Silene seeds with the cardioid led to the application of geometric models in the quantification of seed shape. In general, the mean value of the
J index obtained with the cardioid (LM1) in seeds of species of
S. subg.
Behenantha was higher than in seeds of
S. subg.
Silene [
14]. Nevertheless, the description of other, additional geometric models derived from the cardioid gave better
J index values when applied to the species of
S. subg.
Silene (e.g., LM3 for
S. gallica, LM6 for
S. mellifera, LM5 for
S. tridentata). The same trend applied to the species of
S. subg.
Behenantha (e.g., LM2 for
S. latifolia, LM4 for
S. diclinis) [
14,
15].
In general, the combination of models LM2 to LM8 may be more discriminant than the cardioid (LM1) itself, because they narrow better the morphological characteristics of seeds (open or closed in the hilum region). Geometric models LM2, LM4, and LM8 corresponded to those seeds with a more plane and closed region around the hilum, characterized by low partial concavities and high solidity [
12]. Meanwhile, models LM3, LM5, LM6, and LM7 presented an open region (partial concavity) in the hilum region [
14,
15]. Thus, both groups represent alternative morphologies. In addition, the dorsal view of seeds was also analyzed to provide quantitative data about shape and morphology from this view [
16]. The application of new models for the dorsal views showed that convex models adjusted better to seed images of
S. subg.
Behenantha [
16]. On the contrary, we have seeds with partial concavities in the upper and lower sides of their dorsal views due to a channel running through the profile of the seed. These are known as
dorso canaliculata [
1,
2,
3] and correspond to non-convex models in their dorsal views, such as
S. apetala,
S. colorata Poir.,
S. inaperta, or
S. ramosissima. They are frequently present in
S. subg.
Silene, and less common or absent in some sections of
Silene subg.
Behenantha, such as
Silene sect.
Melandrium [
16].
These results suggest that the comparison with geometric figures might be a useful tool to apply morphological traits in plant taxonomy. Hence, an important question for the consideration of the taxonomic value of these morphological characters based on geometric models concerns their stability. Although these geometric models revealed levels of stability for certain complex taxonomic groups of
Silene [
12,
13,
14,
15,
16,
17,
18], more data are still needed to fully support these previous statements.
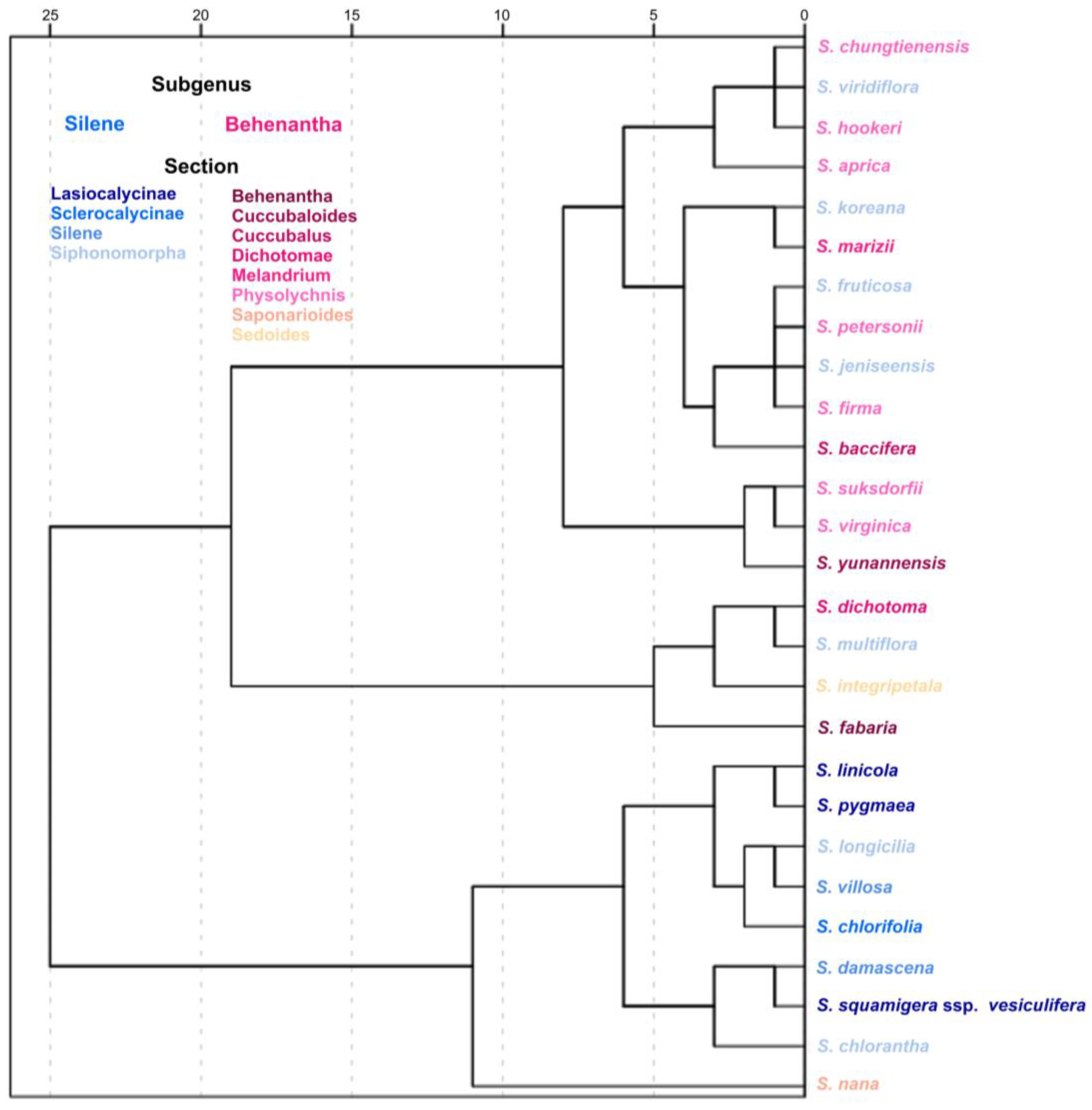
In this work, we have classified 51 species of
Silene from diverse geographic provenances. The classification was according to their highest scores of
J index values with specific geometric models for their lateral and dorsal views. The analysis started with the values of the
J index in the average silhouettes for the lateral and dorsal view of each species. Average silhouettes gave a preliminary approximation to overall seed shape in both views, lateral and dorsal [
15,
16,
17]. They provide information about aspects of seed morphology such as general symmetry, aspect ratio, and convexity. For example, the seeds of eight species resembling model LM5 show positive correspondence in DM5 to DM9, but it is negative in models DM1–DM4. A total of 10 species giving strong similarity in their average silhouettes for the dorsal views with convex models (DM1–DM4) belong to the group of high similarity with convex models LM2 and LM4. Likewise, non-convex models DM8 and DM9 are linked to the group of 17 species which are elongated and gave low results of
J indexes in the tests based on mean values of 20 seeds. All of them, except
S. yunnanensis, were discarded from further study. This might indicate that these seeds require more specific models or that their seeds were quite heterogenous. Nevertheless, elevated values of aspect ratio (elongated seeds) could be a symptom of dehydration due to prolonged storage or storage in adverse conditions [
22].
We found an association between the J index in the average silhouettes and the J index as the mean value of 20 individual seeds. The group of species whose average silhouettes gave higher scores in LM2, LM4, and LM8 contains all the species giving the best scores in these models when estimated as means of 20 seeds. Similarly, most species giving elevated scores of J index as the mean value of 20 individual silhouettes in LM5 (e.g., S. linicola, S. longicilia, S. pygmaea, and S. damascena) also had high values in this model in their average silhouettes. Values obtained with average silhouettes, although indicative of seed shape in a group of seeds, could give overestimations because the analyses with the average silhouettes concentrate more on conserved regions, and hence, the shape of many seeds can differ from their corresponding average silhouette. Thus, the final analysis including J index values measured as the mean of 20 seeds results in much more precise estimations.
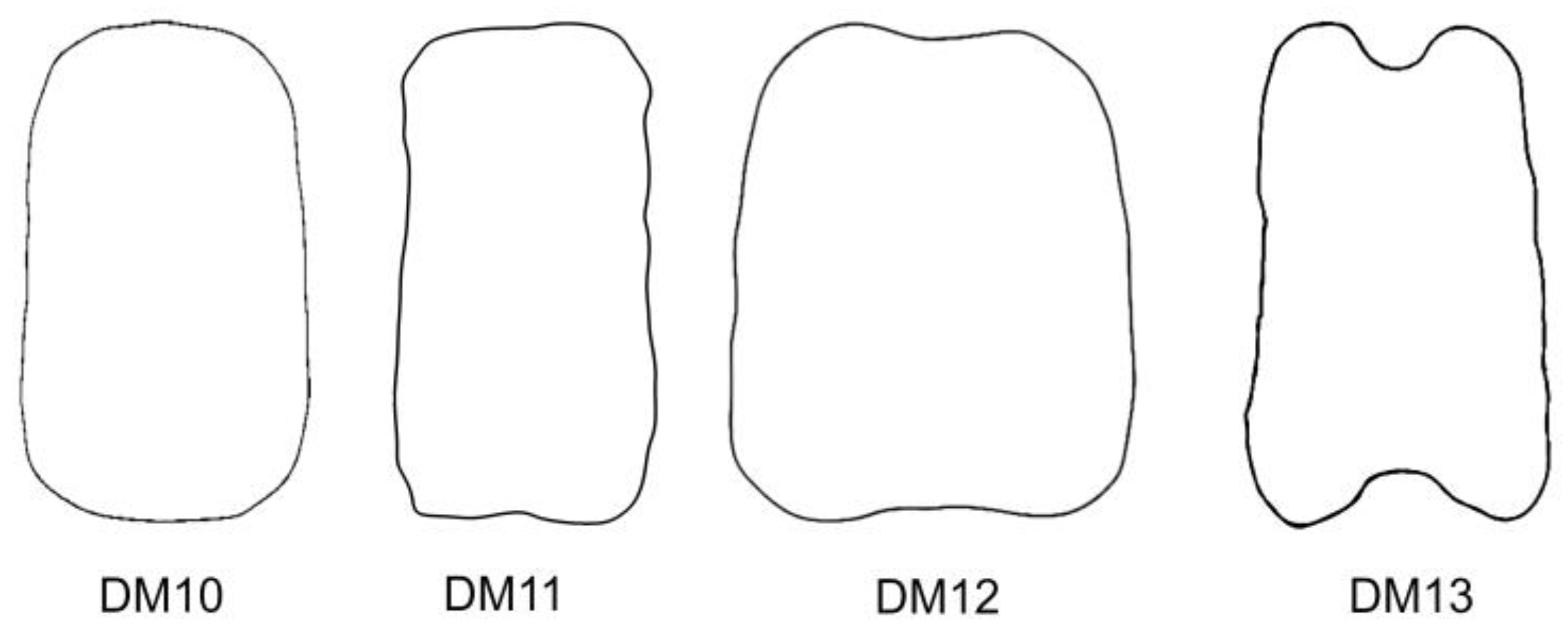
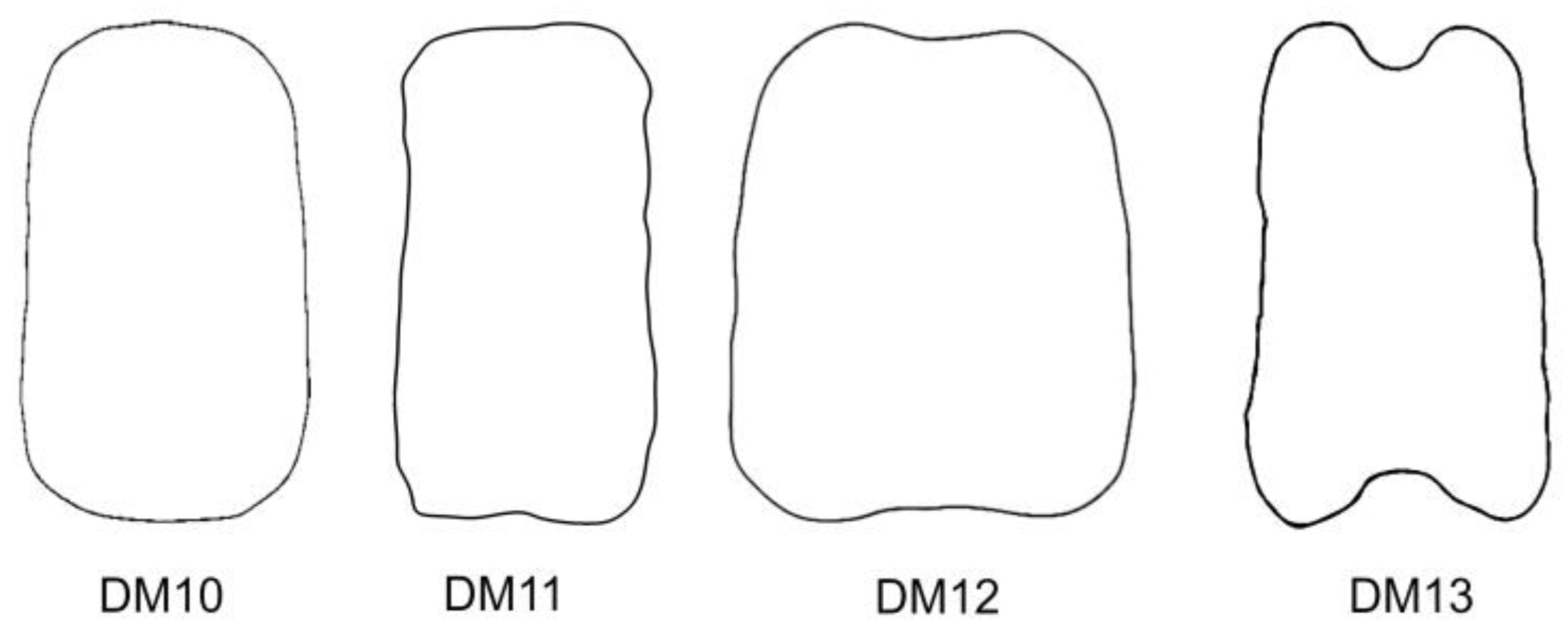
A protocol based on Fourier transform allows us to define models according to their shape, independently of known geometric figures [
18]. The protocol has been applied here to design the new dorsal models DM10 to DM13. Among them, DM10 is convex while the other three models present regions of variable concavities. DM10 fits well for the dorsal view in 11 species. All of them combined with LM2, LM4, or LM8:
S. aprica and
S. koreana (LM2),
S. firma, S. nana, S. chungtienensis,
S. fabaria,
S. suksdorfii, and
S. viridiflora (LM4), and
S. dichotoma,
S. integripetala, and
S. multiflora (LM8). This result would confirm the hypothesis that convex models in the dorsal view are associated with convex models in the lateral view [
16]. Most of these 11 species, whose seeds adjust to convex models, belong to
S. subg.
Behenantha. Model DM11, with small concavity regions, applies also to a variety of species in combination with both convex for LM4 in
S. yunnanensis, and non-convex for LM5 in
S. chlorantha,
S. longicilia, and
S. pygmaea. In contrast to DM10 and DM11, the new models DM12 and DM13 with larger concavity regions are highly specific for
S. linicola and
S. damascena, both combined in LM5, a lateral model presenting a large concavity in the hilum region.
When considering J index values obtained as the mean of 20 measurements, the combination between lateral models closed in the hilum (LM2, LM4, and LM8) and convex models (DM1–DM4, and DM10) was conserved in 18 species: (i) 13 of S. subg. Behenantha: S. aprica, S. baccifera, S. chungtienensis, S. dichotoma, S. fabaria, S. firma, S. hookeri, S. jeniseensis, S. marizii, S. nana, S. petersonii, S. suksdorfii, and S. virginica; and (ii) 5 of S. subg. Silene: S. fruticosa, S. jeniseensis, S. koreana, S. multiflora, and S. viridiflora. These latter five species belong to Silene sect. Siphonomorpha. On the other hand, the combination of lateral models more open in the hilum (LM5) and dorsal models with concavities is more frequent in species of S. subg. Silene. The six species adjusting to LM5 belong to this group: S. squamigera subsp. vesiculifera, S. chlorantha, S. longicilia, S. pygmaea, S. linicola, and S. damascena. From these, one adjusts to DM6 (S. squamigera subsp. vesiculifera), while five of them adjust to the newly designed models DM11 (S. chlorantha, S. longicilia, S. pygmaea), DM12 (S. linicola), and DM13 (S. damascena).
The cardioid (reniform or kidney-shaped) shape of the seeds is associated with the campylotropous ovule, with most of the morphological axis curved, allowing a great morphological variety in ovules and seeds [
23], which is associated with various types of asymmetries [
14]. Consequently, the similarity of seeds with the cardioid is limited to some
Silene species, while others require specific models for accurate description and quantification. The application of Fourier transform allows the description of new, specific models for these seed images that do not adjust well to defined geometrical figures.
New species of
Silene are regularly being described [
24,
25,
26], and the taxonomy of this genus is being revised [
27,
28,
29]. Having the genus
Silene in mind, we present protocols based on seed morphology, which represent a new technique to contribute to this objective. These protocols may be applied with optical photographic equipment and image analysis programs.


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}