
Phylogenetic Position of African Punctoid Snails (Stylommatophora, Punctoidea, Trachycystinae)


Abstract
:1. Introduction
2. Material and Methods
2.1. Taxon Sampling
2.2. DNA Extraction, Amplification and Sequencing
2.3. Sequence Assembly and Alignment
2.4. Phylogenetic Analysis
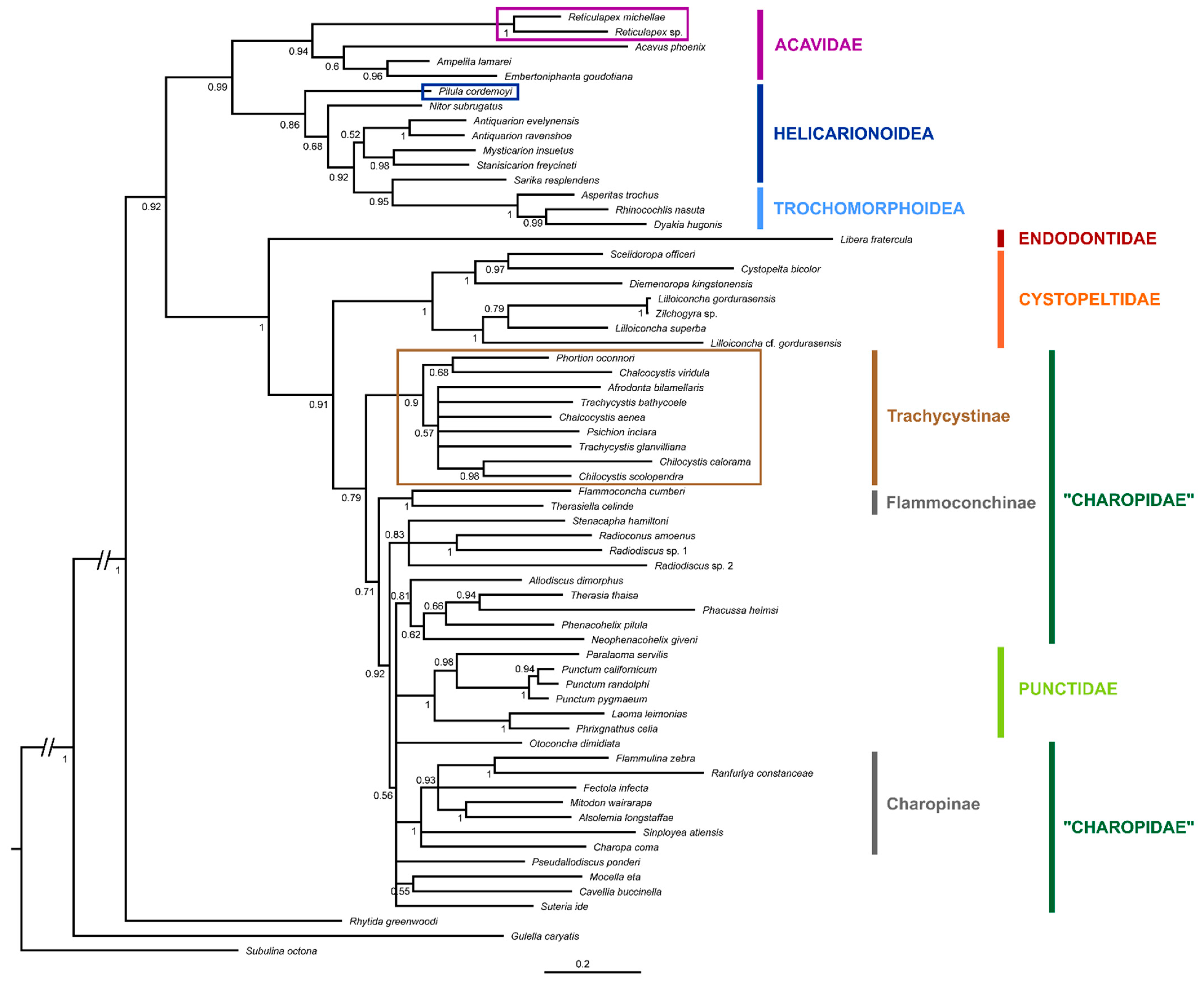
3. Results
4. Discussion
5. Conclusions
| Superfamily Rhytidoidea Pilsbry, 1893 |
| Family Acavidae Pilsbry, 1895 |
| Genus Reticulapex Emberton & Pearce, 2000 |
| Superfamily Punctoidea Morse, 1864 |
| Family Endodontidae Pilsbry, 1895 |
| Family Cystopeltidae Cockerell, 1891 |
| Family Punctidae Morse, 1864 |
| Family “Charopidae” Hutton, 1884 |
| Subfamily Charopinae Hutton, 1884 |
| Subfamily Flammoconchinae Schileyko, 2001 |
| Genus Flammoconcha Dell, 1952 |
| Genus Therasiella Powell, 1948 |
| Subfamily Trachycystinae Schileyko, 1986 |
| Genus Afrodonta Melvill & Ponsonby, 1908 |
| Genus Chalcocystis H. Watson, 1934 |
| Genus Chilocystis H. Watson, 1934 |
| Genus Phortion Preston, 1910 |
| Genus Psichion Gude, 1911 |
| Genus Trachycystis Pilsbry, 1893 |
| Genus Xerocystis H. Watson, 1934 |
| Superfamily Helicarionoidea Bourguignat, 1877 |
| Helicarionoidea incertae sedis |
| Genus Pilula E. von Martens, 1898 |
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salvador, R.B.; Brook, F.J.; Shepherd, L.D.; Kennedy, M.; Brook, F.J.; Shepherd, L.D.; Kennedy, M. Molecular phylogenetic analysis of Punctoidea (Gastropoda, Stylommatophora). Zoosystematics Evol. 2020, 96, 397–410. [Google Scholar] [CrossRef]
- Herbert, D.G.; Kilburn, D. Field Guide to the Land Snails and Slugs of Eastern South Africa; Natal Museum: Pietermaritzburg, South Africa, 2004; p. 336. [Google Scholar]
- Emberton, K.C.; Pearce, T.A. Charopid snails of Mounts Mahermana, Ilapiry, and Vasiha, southeastern Madagascar, with description of a new genus and with conservation statuses of nine species. Veliger 2000, 43, 248–264. [Google Scholar]
- Schileyko, A.A. Treatise on Recent terrestrial pulmonate molluscs. Part 7: Endodontidae, Thyrophorellidae, Charopidae. Ruthenica 2001, Suppl. 2, 881–1034. [Google Scholar]
- Burnup, H.C. On Afrodonta Melv. and Pons., with descriptions of new species. Ann. Natal Mus. 1912, 2, 333–343. [Google Scholar]
- Herbert, D.G. Revision of the aperturally dentate Charopidae (Gastropoda: Stylommatophora) of southern Africa—Genus Afrodonta s. lat., with description of five new genera, twelve new species and one new subspecies. Eur. J. Taxon. 2020, 629, 1–55. [Google Scholar] [CrossRef]
- Thiele, J. Handbuch der Systematichen Weichtierkunde, Erster Band; Gustav Fischer: Jena, Germany, 1931; p. 778. [Google Scholar]
- Watson, H. Natalina and other South African snails. Proc. Malacol. Soc. Lond. 1934, 21, 150–196. [Google Scholar]
- Connolly, M. A monographic survey of South African non-marine Mollusca. Ann. S. Afr. Mus. 1939, 33, 1–660. [Google Scholar]
- Zilch, A. Gastropoda von Wilhelm Wenz. Teil 2. Euthyneura. In Handbuch der Paläozoologie; Band 6, Teil 2; Schindewolf, O.H., Ed.; Gebrüder Borntrager: Berlin, Germany, 1959; pp. 1–835. [Google Scholar]
- Solem, A. The land snail genus Afrodonta (Mollusca: Pulmonata: Endodontidae). Ann. Natal Mus. 1970, 20, 341–364. [Google Scholar]
- Solem, A. Fam. Charopidae. Annales du Musée Royal de l’Afrique Central. Sci. Zool. 1977, 4, 521–533. [Google Scholar]
- Solem, A. Endodontoid Land Snails from Pacific Islands (Mollusca: Pulmonata: Sigmurethra). Part II. Families Punctidae and Charopidae, Zoogeography; Field Museum: Chicago, IL, USA, 1983; p. 336. [Google Scholar]
- Schileyko, A.A. O filogeneticheskikh sviaziakh Trigonochlamydidae (Gastropoda: Pulmonata). Sb. Tr. Zool. Muzeia 1986, 24, 187–196. [Google Scholar]
- Vaught, K.C. A Classification of the Living Mollusca; American Malacologists: Melbourne, FL, USA, 1989; p. 195. [Google Scholar]
- Bouchet, P.; Rocroi, J.-P.; Frýda, J.; Hausdorf, B.; Ponder, W.; Valdés, Á.; Warén, A. Classification and nomenclator of gastropod families. Malacologia 2005, 47, 1–397. [Google Scholar]
- Bouchet, P.; Rocroi, J.-P.; Hausdorf, B.; Kaim, A.; Kano, Y.; Nützel, A.; Parkhaev, P.; Schrödl, M.; Strong, E.E. Revised classification, nomenclator and typification of gastropod and monoplacophoran families. Malacologia 2017, 61, 1–526. [Google Scholar] [CrossRef]
- MolluscaBase. Available online: http://www.molluscabase.org (accessed on 31 March 2022).
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and compilation of Conserved Polymerase Chain Reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Wade, C.M.; Mordan, P.B. Evolution within the gastropod molluscs: Using the ribosomal RNA gene cluster as an indicator of phylogenetic relationships. J. Molluscan Stud. 2000, 66, 565–570. [Google Scholar] [CrossRef] [Green Version]
- Wade, C.M.; Mordan, P.B.; Naggs, F. Evolutionary relationships among the Pulmonate land snails and slugs (Pulmonata, Stylommatophora). Biol. J. Linn. Soc. 2006, 87, 593–610. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Schwartz, T.; Pickett, B.E.; He, S.; Klem, E.B.; Scheuermann, R.H.; Passarotti, M.; Kaufman, S.; O’Leary, M.A. A RESTful API for access to phylogenetic tools via the CIPRES Science Gateway. Evol. Bioinform. 2015, 11, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; van der Mark, P.; Huelsenbeck, J.P. Bayesian phylogenetic analysis using MrBayes. In The Phylogenetic Handbook: A Practical Approach to Phylogenetic Analysis and Hypothesis Testing; Lemey, P., Salemi, M., Vandamme, A.-M., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 210–266. [Google Scholar]
- Van Bruggen, A.C. Land molluscs. Monogr. Biol. 1978, 31, 877–923. [Google Scholar]
- Van Bruggen, A.C. Studies on the land molluscs of Zululand with notes on the distribution of land molluscs in southern Africa. Zool. Verh. 1969, 103, 1–116. [Google Scholar]
- Pickford, M. Land snails from the Early Miocene Legetet Formation, Koru, Kenya. Münchner Geowiss. Abh. A 2019, 49, 1–70. [Google Scholar]
- Kensley, B. A second assemblage of Pliocene invertebrate fossils from Langebaanweg, Cape. Ann. South Afr. Mus. 1977, 72, 189–210. [Google Scholar]
- Malan, J.A. The Stratigraphy and Sedimentology of the Bredasdorp Group, Southern Cape Province; University of Cape Town: Cape Town, South Africa, 1990; p. 197. [Google Scholar]


{kind=link}
| Species | COI | 16S | ITS2+28S | Voucher | Provenance |
|---|---|---|---|---|---|
| African Charopidae | |||||
| Afrodonta bilamellaris Melvill & Ponsonby, 1908 | ON365470 | ON374086 | ON376035 | NMSA-Mol 0W9618 | South Africa, KwaZulu-Natal, Port Shepstone, Four Man’s Hill |
| Chalcocystis viridula (Connolly, 1939) | — | ON374088 | ON376037 | NMSA-Mol 0W9407 | South Africa |
| Chilocystis calorama (Melvill & Ponsonby, 1899) | ON365472 | ON374089 | ON376038 | NMSA-Mol 0W9636 | South Africa, KwaZulu-Natal, Port Shepstone, Simuma Hill |
| Chilocystis scolopendra (Melvill & Ponsonby, 1903) | ON365473 | ON374090 | ON376039 | NMSA-Mol 0W7837 | South Africa, KwaZulu-Natal, Port Shepstone, Hlokohloko Valley |
| Phortion oconnori (Preston, 1912) | ON365475 | ON374092 | ON376041 | NMSA-Mol 0W8889 | South Africa, Western Cape, Bainskloof Pass |
| Psichion inclara (Morelet, 1889) | ON365478 | ON374095 | ON376043 | NMSA-Mol 0W9163 | South Africa, Eastern Cape, Woody Cape Nature Reserve |
| Trachycystis bathycoele (Melvill & Ponsonby, 1892) | ON365481 | — | — | NMSA-Mol 0P0178 | South Africa, Limpopo, Entabeni Forest |
| Trachycystis glanvilliana (Ancey, 1890) | ON365482 | ON374097 | ON376047 | NMSA-Mol 0W7149 | South Africa, KwaZulu-Natal, Karkloof Falls |
| Trachycystis tollini (Benson, 1856) | — | — | ON376048 * | NMSA-Mol 0W9407 | South Africa, Western Cape, Platbos Forest Reserve |
| Xerocystis capensis (L. Pfeiffer, 1841) | — | — | ON376049 * | NMSA-Mol 0W8893 | South Africa, Western Cape, De Hoop Nature Reserve |
| African taxa, not Charopidae | |||||
| Pilula cordemoyi (G. Nevill, 1870) | ON365476 | ON374093 | — | UF 415477 | Réunion Island, Basse Valley |
| Reticulapex michellae K.C. Emberton et al., 2010 | ON365479 | — | ON376044 | UF 421124 | Madagascar, Andriantantely Massif |
| Reticulapex sp. | — | — | ON376045 | UF 421119 | Madagascar, Andriantantely Massif |
| New Zealand Charopidae | |||||
| Cavellia buccinella (Reeve, 1852) | ON365471 | ON374087 | ON376036 | OZD Cabuc-1 | New Zealand, Northland, Kaipara |
| Flammoconcha cumberi (Powell, 1941) | ON365474 | ON374091 | ON376040 | OZD FB29 | New Zealand, West Coast, Grange Ridge |
| Pseudallodiscus ponderi Climo, 1971 | ON365477 | ON374094 | ON376042 | OZD Ppond-1 | New Zealand, Auckland, Pahiatua Hill Scenic Reserve |
| Therasiella celinde (Gray, 1850) | ON365480 | ON374096 | ON376046 | OZD FB116 | New Zealand, Waikato, Pokeno |
| Species | COI | 16S | ITS2+28S | Provenance of Voucher |
|---|---|---|---|---|
| Charopidae | ||||
| Allodiscus dimorphus (Reeve, 1852) | MN792581 | MN756708 | MN782439 | New Zealand, Auckland, Waitakere Ranges, Titirangi, Atkinson Track |
| Alsolemia longstaffae (Suter, 1913) | MN792582 | MN756709 | MN759313 | New Zealand, Southland, Colac Bay |
| Chalcocystis aenea (F. Krauss, 1848) | MN792590 | MN756717 | MN782447 | South Africa, KwaZulu-Natal, Hluhluwe |
| Charopa coma (Gray, 1843) | MN792591 | MN756718 | MN782448 | New Zealand, Auckland, Waitakere Ranges, Titirangi, Paturoa Stream |
| Fectola infecta (Reeve, 1852) | MN792600 | MN756727 | MN782457 | New Zealand, Waikato, Coromandel Peninsula, Port Charles |
| Flammulina zebra (Le Guillou, 1842) | MN792601 | MN756728 | MN782458 | New Zealand, Tasman, Lake Daniells |
| Mitodon wairarapa (Suter, 1890) | MN792607 | MN756732 | MN782464 | New Zealand, Southland, Stewart Island, Mason Bay, Gutter |
| Mocella eta (Pfeiffer, 1853) | MN792608 | MN756733 | MN782465 | New Zealand, Northland, Umuheke Bay |
| Neophenacohelix giveni (Cumber, 1961) | MN792609 | MN756743 | MN782466 | New Zealand, Northland, Whangarei, Coronation Reserve |
| Otoconcha dimidiata (L. Pfeiffer, 1853) | MN792614 | MN756738 | MN782471 | New Zealand, Northland, Whangarei, Bream Head |
| Phacussa helmsi (Hutton, 1882) | MN792618 | MN756742 | MN782475 | New Zealand, West Coast, Greymouth, Point Elizabeth |
| Phenacohelix pilula (Reeve, 1852) | MN792619 | MN756744 | MN782476 | New Zealand, Northland, Whangaruru North Head |
| Radioconus amoenus (Thiele, 1927) | MN792623 | MN756749 | MN782481 | Brazil, Santa Catarina, Florianópolis, Gruta do Triângulo |
| Radiodiscus sp. | MN792625 | MN756751 | MN782483 | Brazil, Bahia, Ilhéus |
| Radiodiscus sp. | MN792626 | MN756752 | MN782484 | Chile, Chiloé, Chiloé National Park, Chepu |
| Ranfurlya constanceae Suter, 1903 | MN792627 | MN756753 | MN782485 | New Zealand, Auckland Islands, Adams Island |
| Sinployea atiensis (Pease, 1870) | MN792628 | MN756754 | MN782486 | Cook Islands, Rarotonga, Tupapa Valley |
| Stenacapha hamiltoni (Cox, 1868) | MN792629 | MN756755 | MN782487 | Australia, Tasmania, Central Plateau, Viormy |
| Suteria ide (Gray, 1850) | MN792630 | MN756756 | MN782488 | New Zealand, Manawatu-Wanganui, Bushy Park |
| Therasia thaisa Hutton, 1883 | MN792631 | MN756757 | MN782489 | New Zealand, Southland, Clifden, Clifden Limestone Cave System |
| Cystopeltidae | ||||
| Cystopelta bicolor Petterd & Hedley, 1909 | MN792592 | MN756719 | MN782449 | Australia, Tasmania, Bronte Park |
| Diemenoropa kingstonensis (Legrand, 1871) | MN792616 | MN756740 | MN782473 | Australia, Tasmania, Skullbone Plains, Kenneth Lagoon |
| Lilloiconcha cf. gordurasensis (Thiele, 1927) | MN792604 | MN756731 | MN782461 | Brazil, Alagoas, Pedra Talhada Biological Reserve |
| Lilloiconcha gordurasensis (Thiele, 1927) | MN792605 | — | MN782462 | Brazil, São Paulo, São Paulo, Burle Marx Park |
| Lilloiconcha superba (Thiele, 1927) | MN792606 | — | MN782463 | Brazil, Alagoas, Pedra Talhada Biological Reserve |
| Scelidoropa officeri (Legrand, 1871) | MN792617 | MN756741 | MN782474 | Australia, Tasmania, Flinders Island, Brougham Sugarloaf |
| Zilchogyra sp. | MN792632 | — | MN782490 | Brazil, São Paulo, Cotia, Morro Grande Reserve |
| Endodontidae | ||||
| Libera fratercula (Pease, 1867) | MN792603 | MN756730 | MN782460 | Cook Islands, Rarotonga, Tupapa |
| Punctidae | ||||
| Laoma leimonias (Gray, 1850) | MN792602 | MN756729 | MN782459 | New Zealand, Northland, Kaihu, Maropiu Road |
| Paralaoma servilis (Shuttleworth, 1852) | MN792615 | MN756739 | MN782472 | New Zealand, Southland, Colac Bay |
| Phrixgnathus celia Hutton, 1883 | MN792620 | MN756745 | MN782477 | New Zealand, Southland, Stewart Island, Mason Bay |
| Punctum californicum Pilsbry, 1898 | MN792621 | MN756746 | MN782478 | USA, California, San Francisco, Presidio, Lincoln Boulevard |
| Punctum pygmaeum (Draparnaud, 1801) | MN812719 | MN756747 | MN782479 | UK, Monmouthshire, Monmouth, Pentwyn Farm |
| Punctum randolphii (Dall, 1895) | MN792622 | MN756748 | MN782480 | Canada, British Columbia, Pemberton, Riverside Trail, Lillooet River |
| Helicarionoidea | ||||
| Antiquarion evelynensis Hyman & Köhler, 2020 | MN654044 | MN654081 | — | Australia, Queensland, Milllaa Millaa |
| Antiquarion ravenshoe Hyman & Köhler, 2020 | MN654053 | MN654089 | — | Australia, Queensland, Tully |
| Mysticarion insuetus Iredale, 1941 | KY662466 | KY662376 | — | Australia, New South Wales, Newcastle |
| Nitor subrugatus (Reeve, 1852) | MH248130 | MH255873 | — | Australia, New South Wales, Tooloom National Park |
| Sarika resplendens (Philippi, 1847) | MT364982 | MT365763 | MT365707 | Thailand |
| Stanisicarion freycineti (Férussac, 1821) | MN073628 | MN073747 | — | Australia, Queensland, Eastern Escarpment Conservation Area |
| Rhytidoidea | ||||
| Acavus phoenix (L. Pfeiffer, 1854) | — | — | AY014083 | Unknown |
| Ampelita lamarei (L. Pfeiffer, 1853) | — | — | KP230504 | Madagascar, Msoala Peninsula |
| Embertoniphanta goudotiana (Férussac, 1839) | — | — | KP230517 | Madagascar, Tsingy Beanka, Belitsaka |
| Rhytida greenwoodi (Gray, 1850) | KT970868 | KT970900 | KP230525 | New Zealand, Waikato, Raglan |
| Trochomorphoidea | ||||
| Asperitas trochus (O. F. Müller, 1774) | MT654630 | MT651546 | MT651601 | Indonesia |
| Dyakia hugonis (L. Pfeiffer, 1864) | MT803064 | MT741748 | MT741915 | Malaysia, Sabah, Segaliud Lokan Forest Reserve |
| Rhinocochlis nasuta (Metcalfe, 1852) | MT803094 | MT741771 | MT741938 | Malaysia, Sarawak, Sibu, Lanjak Entimau |
| Outgroup | ||||
| Gulella caryatis (Melvill & Ponsonby, 1898) | HQ328133 | HQ328323 | GQ330510 | Namibia |
| Subulina octona (Bruguière, 1789) | JX988066 | JX988353 | MF444887 | Palau |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvador, R.B. Phylogenetic Position of African Punctoid Snails (Stylommatophora, Punctoidea, Trachycystinae). Taxonomy 2022, 2, 227-235. https://doi.org/10.3390/taxonomy2020017
Salvador RB. Phylogenetic Position of African Punctoid Snails (Stylommatophora, Punctoidea, Trachycystinae). Taxonomy. 2022; 2(2):227-235. https://doi.org/10.3390/taxonomy2020017
Chicago/Turabian StyleSalvador, Rodrigo Brincalepe. 2022. "Phylogenetic Position of African Punctoid Snails (Stylommatophora, Punctoidea, Trachycystinae)" Taxonomy 2, no. 2: 227-235. https://doi.org/10.3390/taxonomy2020017
APA StyleSalvador, R. B. (2022). Phylogenetic Position of African Punctoid Snails (Stylommatophora, Punctoidea, Trachycystinae). Taxonomy, 2(2), 227-235. https://doi.org/10.3390/taxonomy2020017