
A New Species of Large Duck (Aves: Anatidae) from the Miocene of New Zealand †

 , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
3. Results
| SYSTEMATICS |
| Class Aves Linnaeus, 1758 |
| Order Anseriformes Wagler, 1831 |
| Family Anatidae Leach, 1819 |
| Subfamily Tadorninae Reichenbach, 1849: Shelducks |
| Tribe Tadornini Reichenbach, 1849 |
| Genus Miotadorna Worthy, Tennyson, Jones, McNamara and Douglas, 2007 |
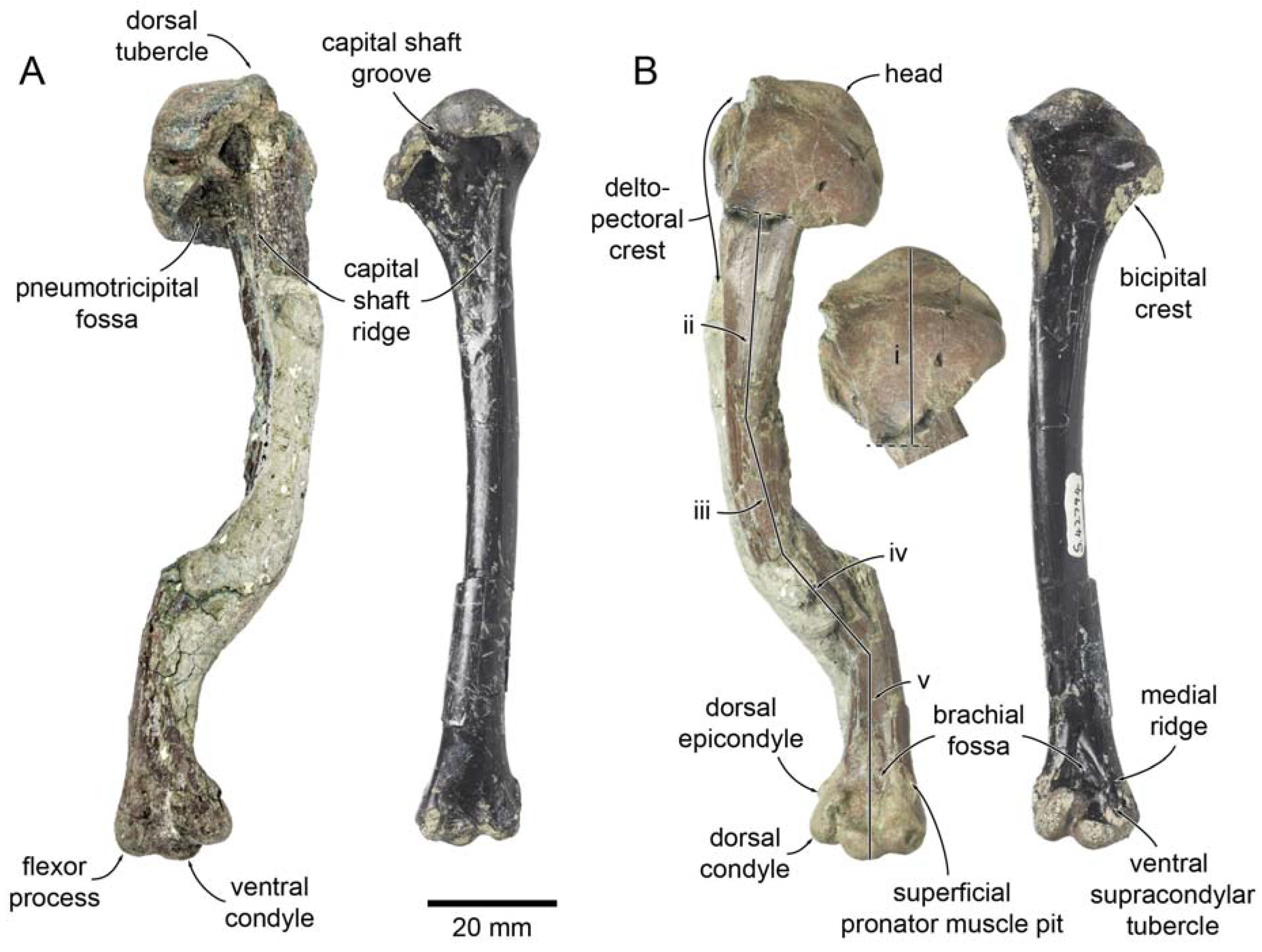
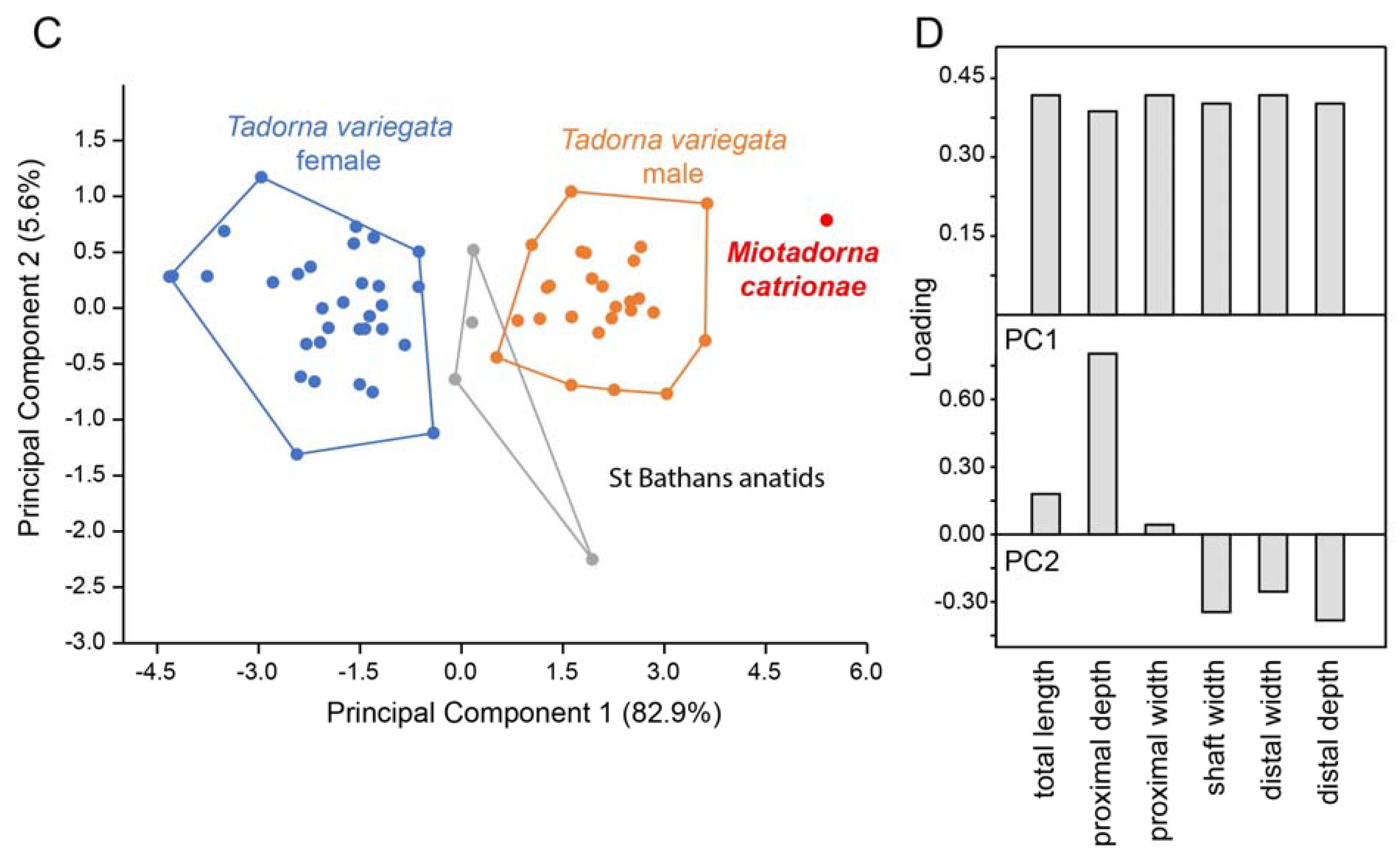
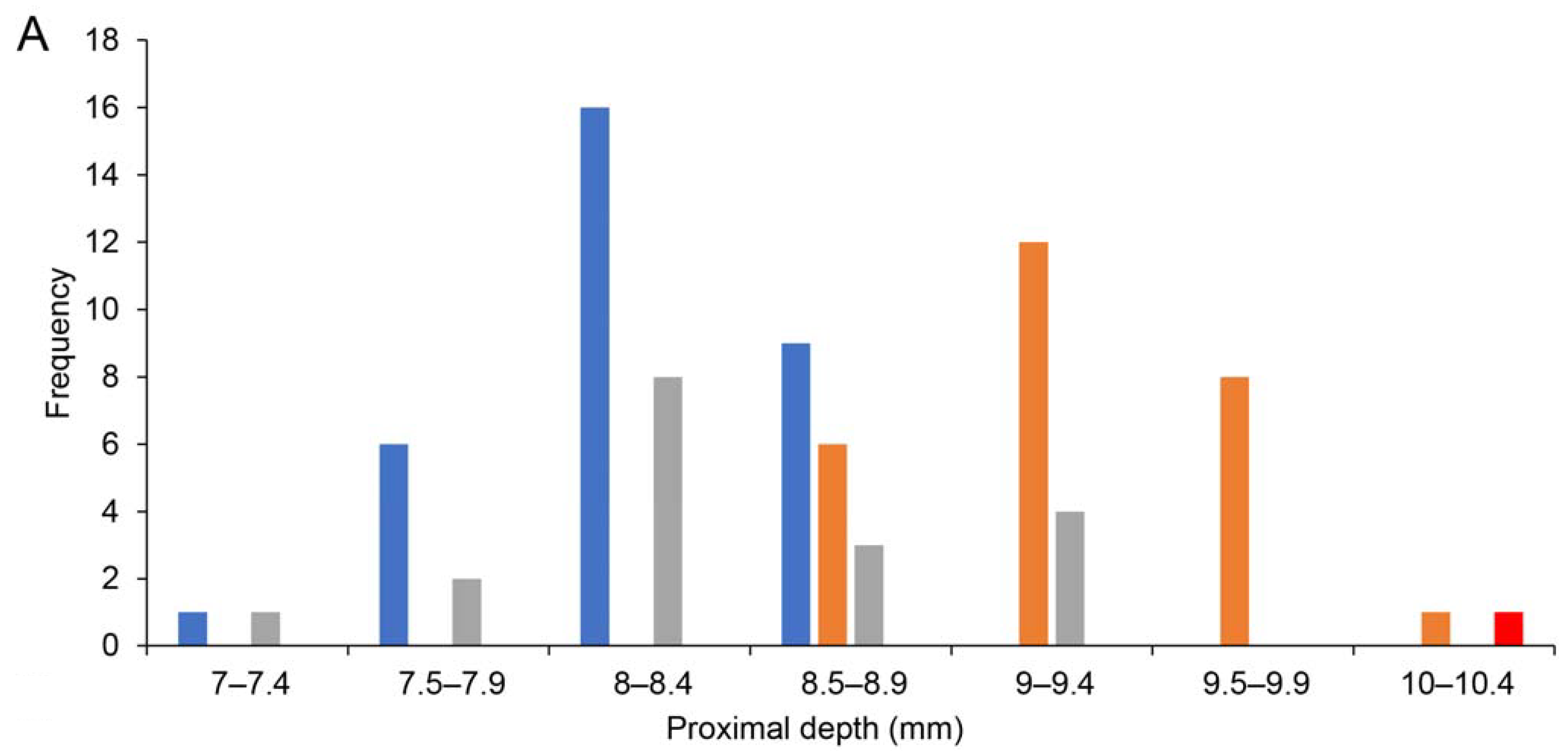
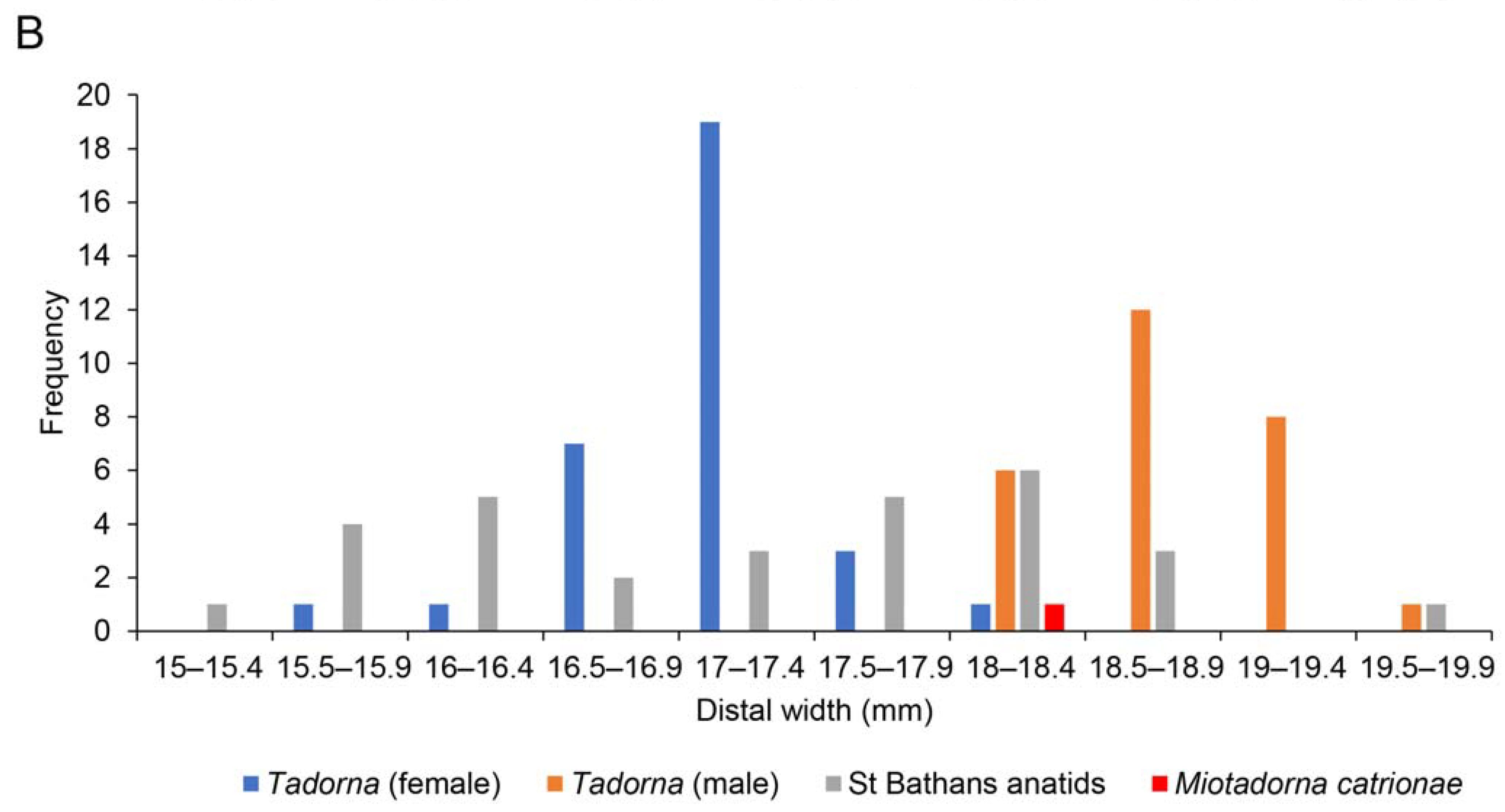
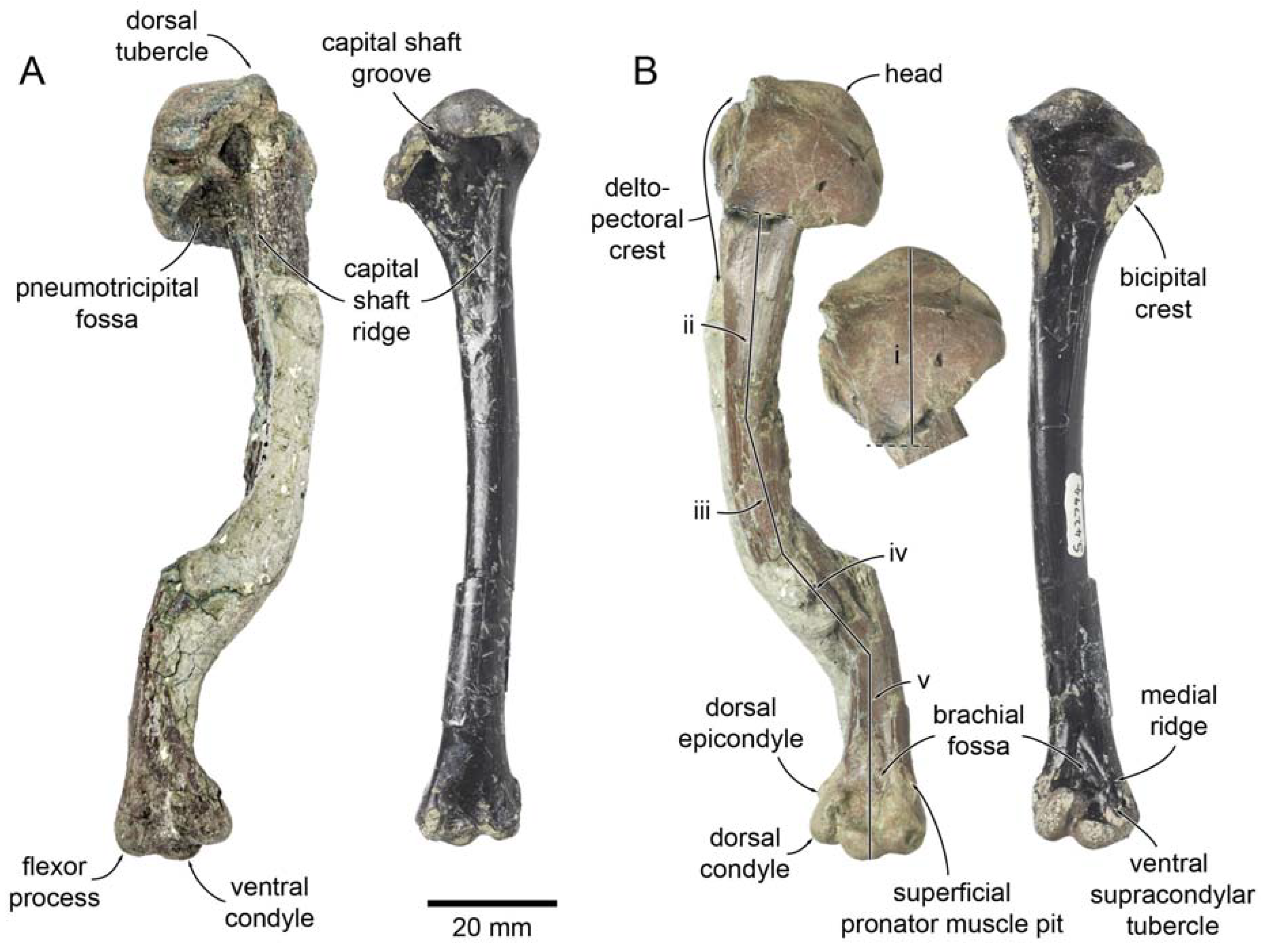
| Miotadorna catrionaesp. nov. Tennyson, Greer, Lubbe, Marx, Richards, Giovanardi and Rawlence |
| (Figure 1) |
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
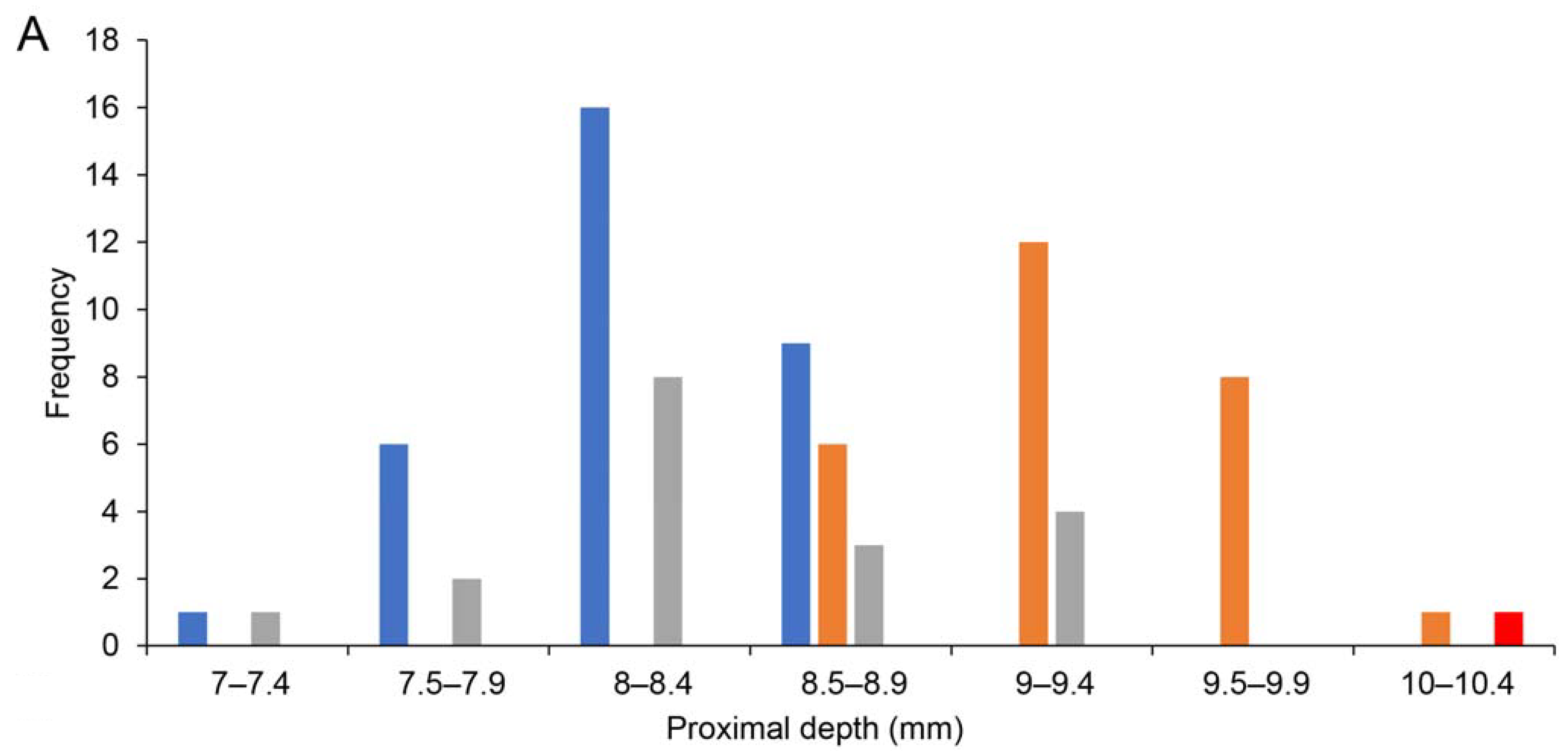
| Taxon | Age/Sex | Reg. No. | Location | Hum TL | pHum Wid | pHum Dep | Shaft Wid | dHum Wid | dHum Dep |
|---|---|---|---|---|---|---|---|---|---|
| St Bathans anatid ** | - | S.42234 | Croc L1 | 125.7 | 24.7 | 8.2 | 10.8 | 19.9 | 10.9 |
| St Bathans anatid ** | - | S.42272 | Croc L1 | 114 | - | 8.4 | 9.5 | - | - |
| St Bathans anatid ** | - | S.42273 | Croc L1 | 119.4 | - | 7.2 | 9.9 | 16.5 | - |
| St Bathans anatid ** | - | S.42274 | Croc L1 | - | 23.8 | 8.3 | - | - | - |
| St Bathans anatid ** | - | S.42275 | Croc L1 | - | - | - | - | 18.1 | 10.2 |
| St Bathans anatid ** | - | S.42313 | HH1a | - | - | - | - | 15.9 | 9.1 |
| St Bathans anatid ** | - | S.42497 | Jim’s Mata | - | - | - | - | 17.1 | 10.5 |
| St Bathans anatid ** | - | S.42558 | HH1a | - | - | - | - | 16.4 | 9.1 |
| St Bathans anatid * | - | S.42794 | HH1a | 122.8 | 25.3 | 8.3 | 10.1 | 17.3 | 10.3 |
| St Bathans anatid ** | - | S.42796 | HH1a | - | - | - | - | 18.1 | 10.8 |
| St Bathans anatid ** | - | S.42810 | Croc L1 | - | - | - | - | 18.7 | 10.6 |
| St Bathans anatid ** | - | S.42931 | Croc L3 | - | - | - | - | 17.9 | 10.9 |
| St Bathans anatid ** | - | S.42932 | Croc L3 | - | - | - | - | 16.6 | - |
| St Bathans anatid ** | - | S.43069 | HH1a | - | - | - | - | - | - |
| St Bathans anatid ** | - | S.43131 | HH1a | - | - | 9.0 | - | - | - |
| St Bathans anatid ** | - | S.43974 | HH1a | - | - | - | - | 15.8 | 8.6 |
| St Bathans anatid ** | - | S.44162 | HH1a | 122.4 | 24.8 | 9.1 | 9.9 | 18.4 | 9.8 |
| St Bathans anatid ** | - | S.44221 | HH1a | - | 25.7 | 8.6 | 9.9 | - | - |
| St Bathans anatid ** | - | S.44352 | Below HH1d | - | 24.3 | 9.3 | - | - | - |
| St Bathans anatid *** | - | S.47273 | Mata 9 | 142.5 | 28.8 | 10.2 | 11.1 | 18.4 | 11.3 |
| St Bathans anatid | - | S.49406 | Mata 9 | - | - | - | - | 17.7 | - |
| St Bathans anatid | - | S.49415 | Mata 9 | - | 25.0 | 8.9 | 9.5 | - | - |
| St Bathans anatid | S.49448 | Croc L1 | - | - | 7.6 | - | - | - | |
| St Bathans anatid | - | S.50045 | HH1a | - | - | - | - | 15.6 | 8.6 |
| St Bathans anatid | - | S.50082 | HH1a | - | - | - | - | 17.0 | 9.7 |
| St Bathans anatid | - | S.50106 | HH1a | - | - | - | - | 17.5 | 10.1 |
| St Bathans anatid | - | S.50117 | HH1a | - | - | - | - | 16.4 | 9.4 |
| St Bathans anatid | - | S.50122 | HH1a | - | - | 7.6 | - | 17.7 | - |
| St Bathans anatid | - | S.50189 | Trench (HH1b) | - | - | 8.2 | - | - | - |
| St Bathans anatid | - | S.50957 | Trench (HH1b) | - | - | - | - | 15.6 | - |
| St Bathans anatid | - | S.50959 | Trench (HH1b) | - | - | - | - | 16.0 | 9.6 |
| St Bathans anatid | - | S.51151 | Trench (HH1b) | - | - | - | - | 16.2 | 9.1 |
| St Bathans anatid | - | S.51220 | Trench (HH1b) | - | - | - | - | 18.0 | - |
| St Bathans anatid | - | S.51284 | Trench (HH1b) | - | - | - | - | 15.1 | 8.4 |
| St Bathans anatid | - | S.51411 | HH1a | - | - | - | - | 18.2 | 10.5 |
| St Bathans anatid | - | S.51592 | HH1a | - | - | 8.0 | - | - | - |
| St Bathans anatid | - | S.51728 | Trench (HH1b) | - | - | - | - | 18.9 | 11.0 |
| St Bathans anatid | - | S.52562 | HH1a | - | - | - | - | 16.1 | 9.4 |
| St Bathans anatid | - | S.52934 | Trench (HH1b) | - | - | - | - | 18.0 | 10.0 |
| St Bathans anatid | - | S.53148 | Trench (HH1b) | - | 24.7 | 8.0 | - | - | - |
| St Bathans anatid | - | S.53038 | Trench (HH1b) | - | - | 9.0 | 11.0 | - | - |
| St Bathans anatid | - | S.53569 | Mata 9 | 121.4 | 27.2 | 8.4 | 10.0 | 17.8 | 9.7 |
| St Bathans anatid | - | OU.21957 | Vinegar Hill | - | 26.3 | 8.6 | - | 18.6 | 10.1 |
| Tadorna variegata | ad F | OR.16471 | Gisborne | 120.7 | 23.3 | 8.1 | 9.9 | 17.3 | 9.1 |
| Tadorna variegata | ad M | OR.16472 | Gisborne | 130.8 | 26.5 | 10.3 | 11.2 | 19.1 | 10.4 |
| Tadorna variegata | ad M | OR.16473 | Gisborne | 128.7 | 25.3 | 9.5 | 10.5 | 18.7 | 10.2 |
| Tadorna variegata | ad F | OR.16501 | Gisborne | 119.4 | 24.1 | 8.0 | 9.8 | 17.3 | 10.0 |
| Tadorna variegata | ad F | OR.16590 | Gisborne | 120.0 | 24.1 | 8.8 | 10.0 | 17.7 | 9.8 |
| Tadorna variegata | ad F | OR.24559 | Wellington | 118.5 | 24.0 | 8.8 | 9.4 | 16.8 | 9.8 |
| Tadorna variegata | ad M | OR.25139 | Wellington | 130.2 | 25.8 | 9.5 | 10.5 | 19.5 | 10.7 |
| Tadorna variegata | ad M | OR.25669 | Wairarapa | 130.1 | 25.9 | 9.2 | 11.5 | 19.0 | 10.6 |
| Tadorna variegata | ad M | OR.26562 | Bay of Plenty | 132.0 | 26.7 | 9.5 | 11.0 | 19.4 | 10.8 |
| Tadorna variegata | ad F | OR.26563 | Bay of Plenty | 122.2 | 24.1 | 8.2 | 10.1 | 17.3 | 9.3 |
| Tadorna variegata | ad F | OR.29041 | Wellington | 119.3 | 23.8 | 8.3 | 9.2 | 17.2 | 9.9 |
| Tadorna variegata | ad F | OR.29042 | Wellington | 122.8 | 23.5 | 8.2 | 10.0 | 17.1 | 9.6 |
| Tadorna variegata | ad M | OR.29052 | Canterbury | 131.7 | 25.9 | 9.2 | 10.5 | 18.8 | 10.2 |
| Tadorna variegata | ad F | OR.29053 | Wellington | 119.4 | 24.4 | 8.4 | 10.3 | 17.4 | 9.7 |
| Tadorna variegata | ad F | OR.29054 | Wellington | 118.8 | 23.4 | 7.7 | 9.4 | 17.2 | - |
| Tadorna variegata | ad F | OR.29061 | Wellington | 117.2 | 22.3 | 8.7 | 8.9 | 16.8 | 9.3 |
| Tadorna variegata | imm F | OR.29133 | Canterbury | 117.5 | 23.5 | 8.2 | 9.9 | 17.0 | 9.5 |
| Tadorna variegata | F | OR.30054 | Otago | 117.2 | 24.1 | 8.3 | 10.4 | 18.0 | 10.2 |
| Tadorna variegata | F | OR.30205 | Wellington | 119.2 | 23.5 | 8.9 | 10.1 | 17.0 | 9.5 |
| Tadorna variegata | juv F | OR.30312 | Northland | 120.4 | 23.6 | 7.9 | 9.7 | 17.1 | 9.4 |
| Tadorna variegata | juv M | OR.30349 | Northland | 130.2 | 25.3 | 8.8 | 10.0 | 18.3 | 10.4 |
| Tadorna variegata | juv F | OR.30350 | Northland | 118.2 | 22.7 | 7.7 | 9.0 | 16.5 | 8.9 |
| Tadorna variegata | ad M | OR.30365 | Northland | 132.2 | 26.2 | 9.6 | 10.8 | 18.8 | 10.2 |
| Tadorna variegata | juv F | OR.30366 | Northland | 116.0 | 22.2 | 7.7 | 8.8 | 15.8 | 9.2 |
| Tadorna variegata | juv F | OR.30367 | Northland | 115.6 | 22.0 | 7.7 | 8.9 | 16.1 | 9.0 |
| Tadorna variegata | juv M | OR.30368 | Northland | 131.7 | 24.7 | 9.1 | 10.0 | 18.7 | 9.9 |
| Tadorna variegata | ad F | OR.30376 | Northland | 117.6 | 22.6 | 8.1 | 9.0 | 16.5 | 9.0 |
| Tadorna variegata | ad F | OR.30377 | Northland | 116.1 | 23.1 | 8.1 | 9.4 | 17.1 | 9.8 |
| Tadorna variegata | juv M | OR.30378 | Northland | 131.0 | 25.4 | 9.3 | 10.5 | 18.6 | 10.3 |
| Tadorna variegata | juv M | OR.30379 | Northland | 128.8 | 26.0 | 8.6 | 10.6 | 18.8 | 10.2 |
| Tadorna variegata | juv M | OR.30380 | Northland | 129.6 | 26.4 | 9.4 | 10.6 | 19.2 | 10.4 |
| Tadorna variegata | ad M | OR.30427 | Northland | 128.9 | 24.7 | 8.8 | 9.8 | 18.3 | 10.5 |
| Tadorna variegata | juv M | OR.30428 | Northland | 130.4 | 25.5 | 9.0 | 10.3 | 18.3 | 10.5 |
| Tadorna variegata | ad M | OR.30429 | Northland | 130.5 | 25.4 | 9.7 | 10.8 | 18.9 | 10.4 |
| Tadorna variegata | juv F | OR.30436 | Northland | 118.0 | 23.0 | 7.4 | 10.0 | 17.0 | 9.6 |
| Tadorna variegata | ad M | OR.30437 | Northland | 128.9 | 25.1 | 9.5 | 10.3 | 18.8 | 10.3 |
| Tadorna variegata | ad F | OR.30438 | Northland | 121.6 | 23.8 | 8.4 | 9.4 | 17.4 | 9.7 |
| Tadorna variegata | ad M | OR.30444 | Northland | 134.5 | 25.6 | 9.1 | 10.5 | 19.0 | 10.3 |
| Tadorna variegata | juv M | OR.30445 | Northland | 132.8 | 25.5 | 8.8 | 10.5 | 18.8 | 10.8 |
| Tadorna variegata | juv F | OR.30448 | Northland | 122.8 | 24.1 | 8.4 | 9.7 | 17.3 | 9.6 |
| Tadorna variegata | ad F | OR.30449 | Northland | 120.9 | 24.4 | 8.9 | 10.0 | 17.4 | 9.7 |
| Tadorna variegata | juv M | OR.30450 | Northland | 128.4 | 25.7 | 9.0 | 10.0 | 18.6 | 10.2 |
| Tadorna variegata | juv M | OR.30451 | Northland | 130.4 | 26.3 | 8.9 | 10.6 | 19.2 | 10.0 |
| Tadorna variegata | ad M | OR.30456 | Northland | 127.3 | 25.9 | 9.5 | 10.7 | 19.0 | 10.6 |
| Tadorna variegata | ad M | OR.30457 | Northland | 131.6 | 25.5 | 9.2 | 10.5 | 18.7 | 10.6 |
| Tadorna variegata | ad F | OR.30458 | Northland | 117.9 | 24.0 | 8.3 | 9.6 | 17.4 | 9.8 |
| Tadorna variegata | juv F | OR.30460 | Northland | 117.1 | 22.7 | 8.2 | 8.9 | 16.9 | 9.7 |
| Tadorna variegata | juv M | OR.30461 | Northland | 127.2 | 25.0 | 8.5 | 9.7 | 18.2 | 10.5 |
| Tadorna variegata | juv F | OR.30462 | Northland | 120.2 | 23.8 | 8.5 | 9.9 | 17.4 | 9.7 |
| Tadorna variegata | ad M | OR.30463 | Northland | 133.8 | 25.4 | 9.3 | 10.5 | 19.1 | 10.5 |
| Tadorna variegata | juv F | OR.30464 | Northland | 119.9 | 23.4 | 8.0 | 10.0 | 17.7 | 9.6 |
| Tadorna variegata | juv F | OR.30465 | Northland | 117.6 | 23.7 | 7.8 | 9.8 | 16.9 | 9.6 |
| Tadorna variegata | ad F | OR.30466 | Northland | 118.3 | 23.7 | 8.5 | 9.6 | 17.5 | 10.1 |
| Tadorna variegata | juv F | OR.30467 | Northland | 118.9 | 23.7 | 8.7 | 9.7 | 17.1 | 9.5 |
| Tadorna variegata | juv F | OR.30468 | Northland | 115.3 | 23.4 | 8.5 | 9.3 | 17.1 | 9.6 |
| Tadorna variegata | juv F | OR.30521 | Northland | 118.9 | 23.5 | 8.2 | 8.9 | 16.9 | 9.7 |
| Tadorna variegata | ad M | OR.30709 | Wellington | 129.2 | 25.2 | 9.1 | 10.4 | 18.4 | 10.5 |
| Tadorna variegata | ad M | OR.30742 | Wairarapa | 129.3 | 25.1 | 9.1 | 10.3 | 18.4 | 10.2 |
| Tadorna variegata | ad M | OR.30746 | Wairarapa | 130.3 | 25.5 | 9.6 | 10.2 | 18.5 | 10.0 |
References
- Reichgelt, T.; Kennedy, E.M.; Conran, J.G.; Mildenhall, D.C.; Lee, D.E. The early Miocene paleolake Manuherikia: Vegetation heterogeneity and warm-temperate to subtropical climate in southern New Zealand. J. Paleolimnol. 2015, 53, 349–365. [Google Scholar] [CrossRef]
- Worthy, T.H.; Scofield, R.P.; Salisbury, S.W.; Hand, S.J.; De Pietri, V.L.; Blokland, J.C.; Archer, M. A new species of Manuherikia (Aves: Anatidae) provides evidence of faunal turnover in the St Bathans fauna, New Zealand. Geobios 2021, in press. [Google Scholar] [CrossRef]
- Douglas, B.J. Manuherikia Group of Central Otago, New Zealand: Stratigraphy, Depositional Systems, Lignite Resource Assessment and Exploration Models. Ph.D. Thesis, University of Otago, Dunedin, New Zealand, 1986. [Google Scholar]
- Schwarzhans, W.; Scofield, R.P.; Tennyson, A.J.D.; Worthy, J.P.; Worthy, T.H. Fish remains, mostly otoliths, from the non-marine early Miocene of Otago, New Zealand. Acta Palaeontol. Pol. 2012, 57, 319–350. [Google Scholar] [CrossRef] [Green Version]
- Worthy, T.H.; Tennyson, A.J.D.; Hand, S.J.; Scofield, R.P. A new species of the diving duck Manuherikia and evidence for geese (Aves: Anatidae: Anserinae) in the St Bathans Fauna (Early Miocene), New Zealand. J. R. Soc. N. Z. 2008, 38, 97–114. [Google Scholar] [CrossRef]
- Worthy, T.H.; Tennyson, A.J.D.; Jones, C.; McNamara, J.A.; Douglas, B.J. Miocene waterfowl and other birds from Central Otago, New Zealand. J. Syst. Palaeontol. 2007, 5, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Baumel, J.J.; Witmer, L.M. Osteologia. In Handbook of Avian Anatomy: Nomina Anatomica Avium; Baumel, J.J., King, A.S., Breazile, J.E., Evans, H.E., Vanden Berge, J.C., Eds.; Nuttall Ornithological Club: Cambridge, MA, USA, 1993; pp. 45–132. [Google Scholar]
- Livezey, B.C.; Zusi, R.L. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. Zool. J. Linn. Soc. 2007, 149, 1–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pole, M.; Douglas, B. A quantitative palynostratigraphy of the Miocene Manuherikia Group, New Zealand. J. R. Soc. N. Z. 1998, 28, 405–420. [Google Scholar] [CrossRef]
- Woolfenden, G.E. Postcranial osteology of the waterfowl. Bulletin of the Florida state museum. Biol. Sci. 1961, 2, 1–129. [Google Scholar]
- Marchant, S.; Higgins, P.J. Handbook of Australian, New Zealand and Antarctic Birds. Vol. 1: Ratites to Ducks; Oxford University Press: Melbourne, Australia, 1990; pp. 1–1400. [Google Scholar]
- Worthy, T.H.; Lee, M.S.Y. Affinities of Miocene waterfowl (Anatidae: Manuherikia, Dunstanetta and Miotadorna) from the St Bathans Fauna, New Zealand. Palaeontology 2008, 51, 677–708. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tennyson, A.J.D.; Greer, L.; Lubbe, P.; Marx, F.G.; Richards, M.D.; Giovanardi, S.; Rawlence, N.J. A New Species of Large Duck (Aves: Anatidae) from the Miocene of New Zealand. Taxonomy 2022, 2, 136-144. https://doi.org/10.3390/taxonomy2010011
Tennyson AJD, Greer L, Lubbe P, Marx FG, Richards MD, Giovanardi S, Rawlence NJ. A New Species of Large Duck (Aves: Anatidae) from the Miocene of New Zealand. Taxonomy. 2022; 2(1):136-144. https://doi.org/10.3390/taxonomy2010011
Chicago/Turabian StyleTennyson, Alan J. D., Liam Greer, Pascale Lubbe, Felix G. Marx, Marcus D. Richards, Simone Giovanardi, and Nicolas J. Rawlence. 2022. "A New Species of Large Duck (Aves: Anatidae) from the Miocene of New Zealand" Taxonomy 2, no. 1: 136-144. https://doi.org/10.3390/taxonomy2010011
APA StyleTennyson, A. J. D., Greer, L., Lubbe, P., Marx, F. G., Richards, M. D., Giovanardi, S., & Rawlence, N. J. (2022). A New Species of Large Duck (Aves: Anatidae) from the Miocene of New Zealand. Taxonomy, 2(1), 136-144. https://doi.org/10.3390/taxonomy2010011