
New Techniques for Seed Shape Description in Silene Species


 ,
,  and
and 
Abstract
:1. Introduction
2. Experimental Section
2.1. Seeds of Silene Analyzed
2.2. Seed Images
2.3. General Morphological Description
2.4. Obtention of an Average Silhouette
2.5. Comparison with Geometric Models: Calculation of the J Index
2.6. Statistical Analysis
3. Results
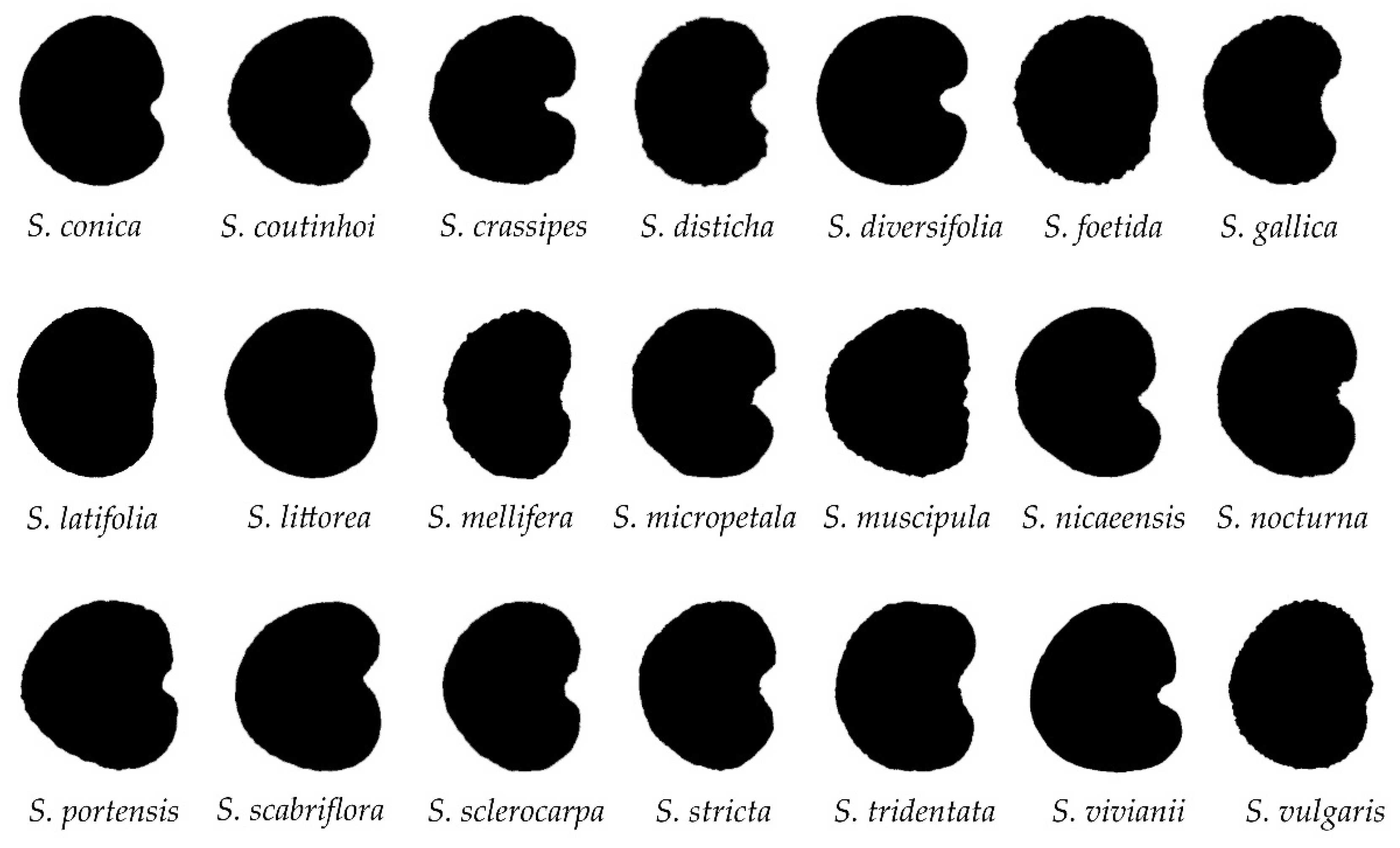
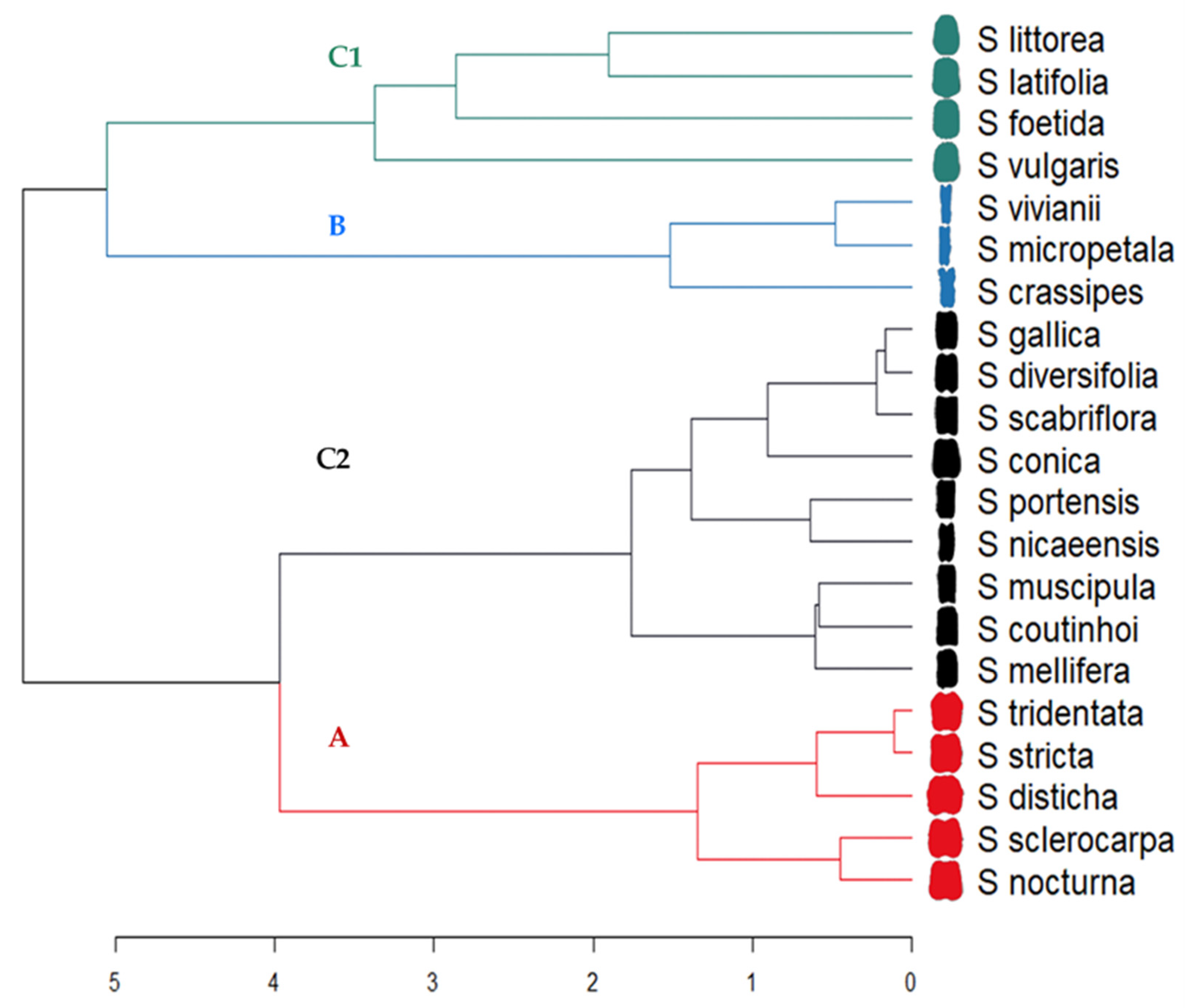
3.1. Description of the Lateral Views of Seeds. Comparison between Silene Species
3.2. Description of the Dorsal Views of Seeds. Comparison between Silene Species
3.3. Comparison of the Lateral Views with the Cardioid. J Index Values
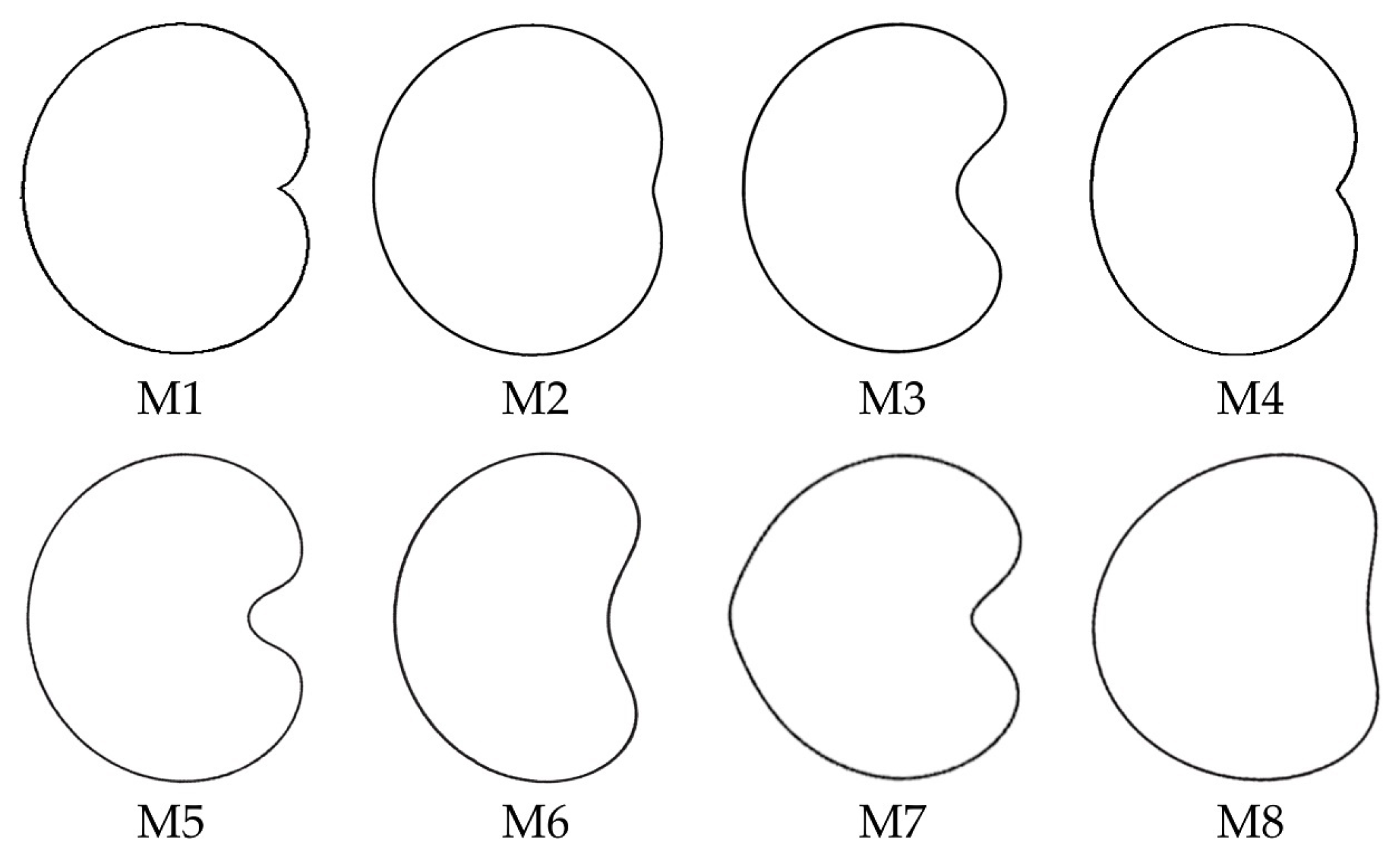
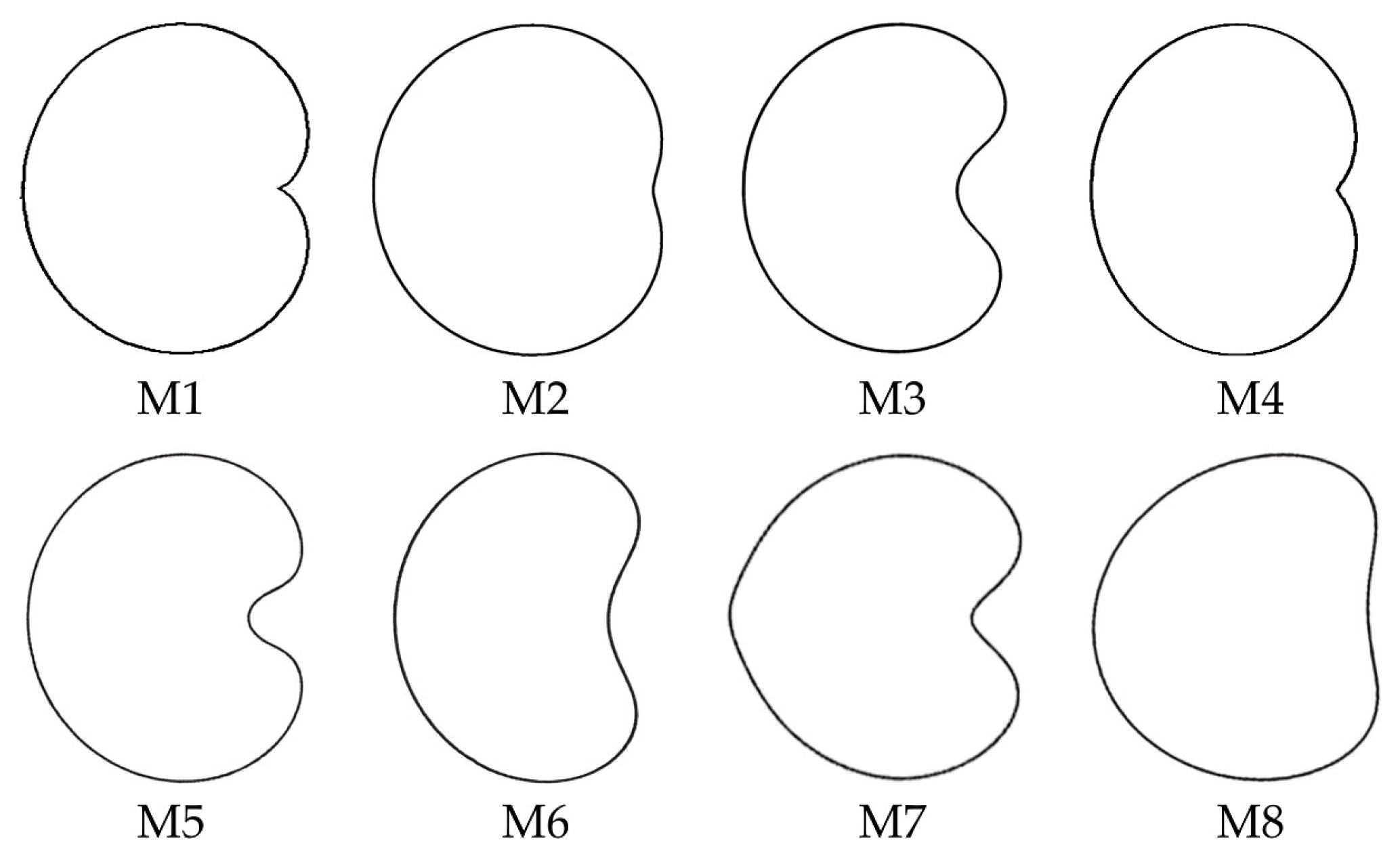
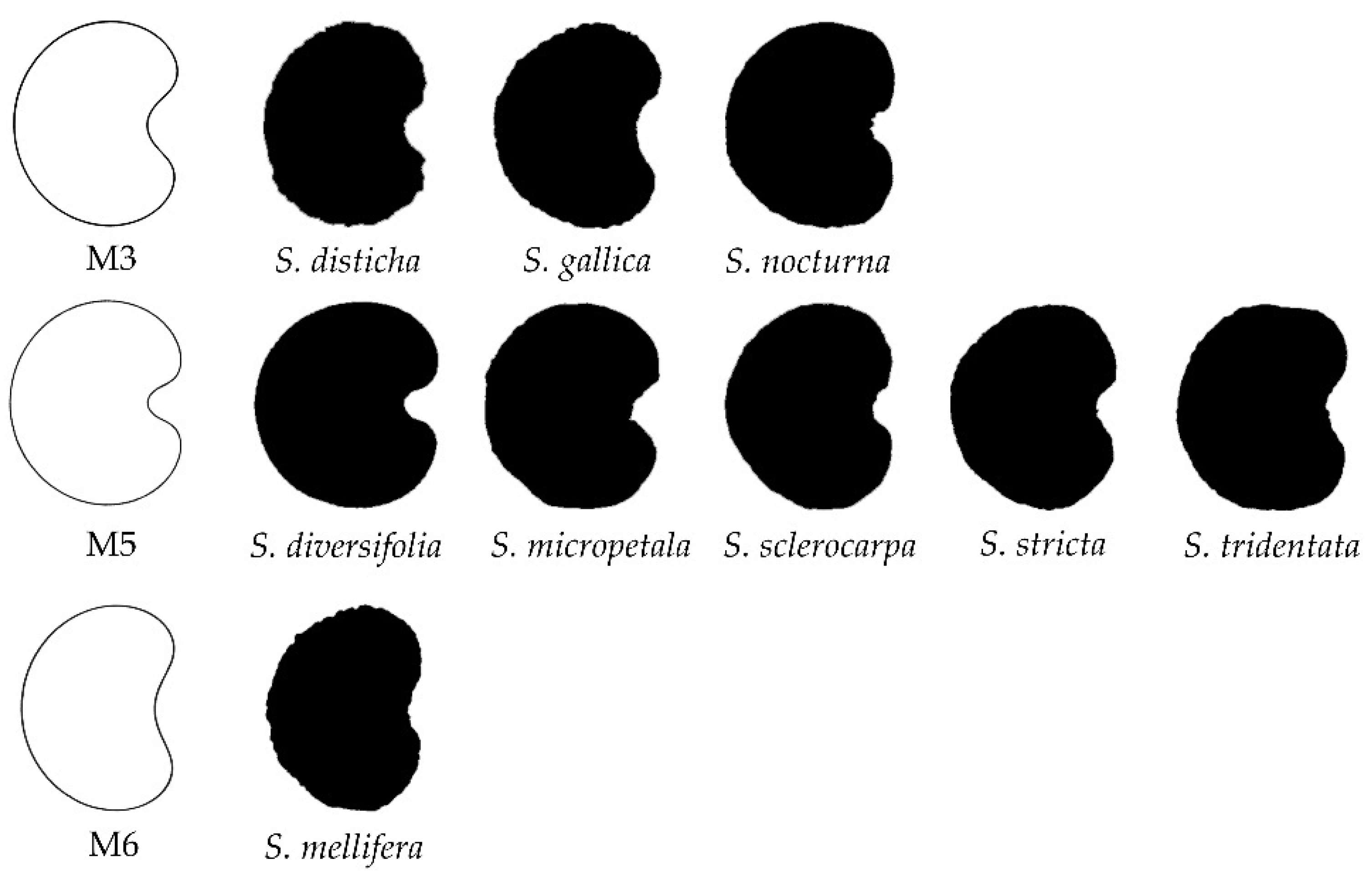
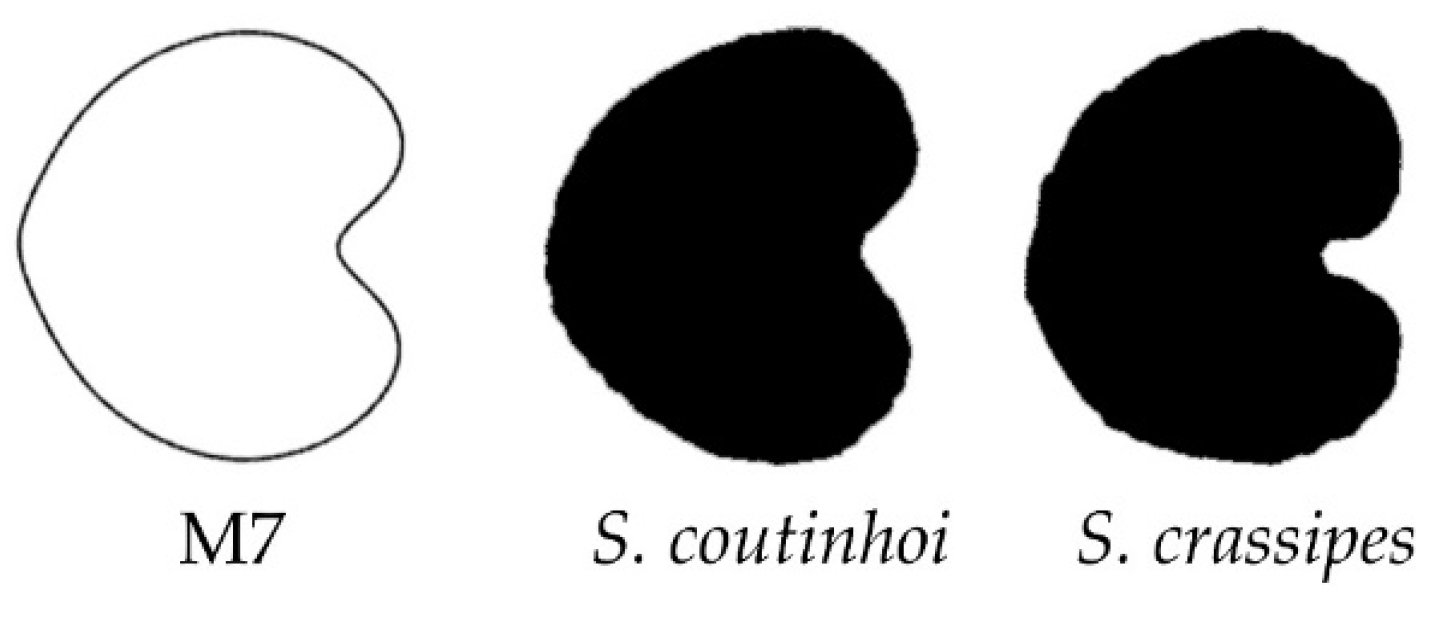
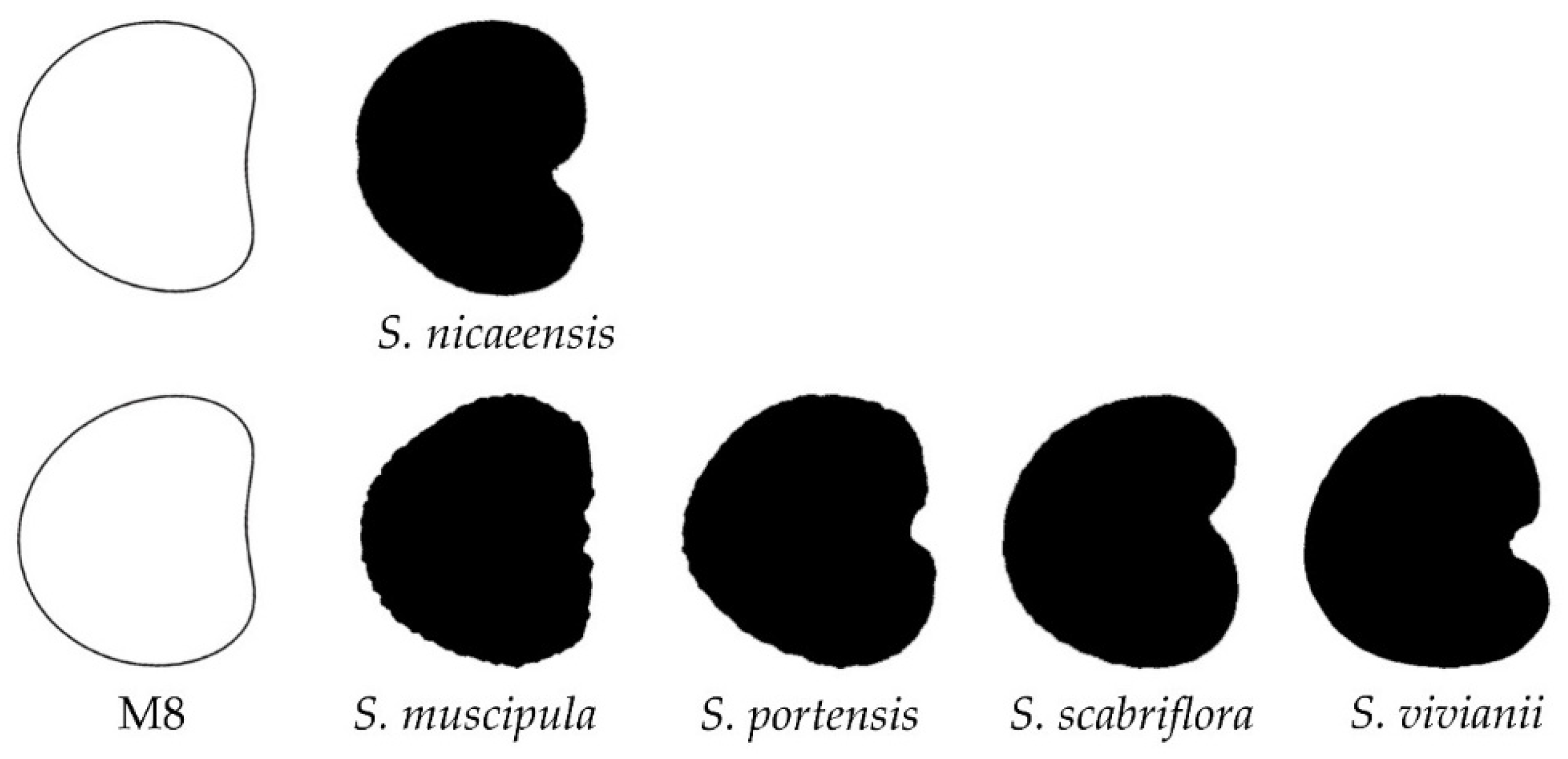
3.4. New Models Obtained to Adjust Seed Shape in Silene Species
3.5. J Index Values Obtained with Models 2 to 8 with Seeds of Silene Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jafari, F.; Zarre, S.; Gholipour, A.; Eggens, F.; Rabler, R.K.; Oxelman, B. A new taxonomic backbone for the infrageneric classification of the species-rich genus Silene (Caryophyllaceae). Taxon 2020, 69, 337–368. [Google Scholar] [CrossRef]
- Boissier, E. Flora Orientalis; Georg, H., Ed.; Bibliopolam: Basel, Switzerland, 1867; Volume 1, pp. 567–656. [Google Scholar]
- Rohrbach, P. Monographic der Gattung Silene; Verlag von Engelmann: Leipzig, Germany, 1869; pp. 1–249. [Google Scholar]
- Oxelman, B.; Rautenberg, A.; Thollesson, M.; Larsson, A.; Frajman, B.; Eggens, F.; Petri, A.; Aydin, Z.; Töpel, M.; Brandtberg-Falkman, A. Sileneae Taxonomy and Systematics. 2013. Available online: http://www.sileneae.info (accessed on 11 September 2021).
- Fawzi, N.M.; Fawzy, A.M.; Mohamed, A.A.A. Seed Morphological Studies on Some Species of Silene L. (Caryophyllaceae). Int. J. Bot. 2010, 6, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Ullah, F.; Ayaz, A.; Saqib, S.; Zaman, W.; Butt, M.A.; Ullah, A. Silene conoidea L.: A review on its systematic ethnobotany and phytochemical profile. Plant Sci. 2019, 6, 373–382. [Google Scholar] [CrossRef]
- Bojňanský, V.; Fargašová, A. Atlas of Seeds and Fruits of Central and East-European Flora: The Carpathian Mountains Region; Springer Science & Business Media: Berlin, Germany, 2007; pp. 1–954. [Google Scholar]
- Yildiz, K.; Cirpici, A. Seed morphological studies of Silene L. from Turkey. Pak. J. Bot. 1998, 30, 173–188. Available online: https://www.researchgate.net/publication/286978973 (accessed on 22 November 2021).
- Atazadeh, N.; Keshavarzi, M.; Sheidai, M.; Gholipour, A. Seed morphology of Silene commelinifolia Boiss. Complex (Caryophyllaceae Juss.). Mod. Phytomorphol. 2017, 11, 5–13. [Google Scholar]
- Keshavarzi, M.; Mahdavinejad, M.; Sheidai, M.; Gholipour, A. Seed and pollen morphology of some Silene species in Iran. Phytol. Balc. 2015, 21, 7–12. [Google Scholar]
- Perveen, A. Seed morphology of the genus Silene: Caryophyllaceae from Pakistan and Kashmir. Int. J. Biol. Biotech. 2009, 6, 219–227. [Google Scholar]
- Cervantes, E.; Martín, J.J.; Ardanuy, R.; de Diego, J.G.; Tocino, Á. Modeling the Arabidopsis seed shape by a cardioid: Efficacy of the adjustment with a scale change with factor equal to the Golden Ratio and analysis of seed shape in ethylene mutants. J. Plant Physiol. 2010, 67, 408–410. [Google Scholar] [CrossRef] [PubMed]
- Martín-Gómez, J.J.; Tocino, Á.; Ardanuy, R.; de Diego, J.G.; Cervantes, E. Dynamic analysis of Arabidopsis seed shape reveals differences in cellulose mutants. Acta Physiol. Plant. 2014, 36, 1585–1592. [Google Scholar] [CrossRef]
- Cervantes, E.; Martín, J.J.; Chan, P.K.; Gresshoff, P.M.; Tocino, Á. Seed shape in model legumes: Approximation by a cardioid reveals differences in ethylene insensitive mutants of Lotus japonicus and Medicago truncatula. J. Plant Physiol. 2012, 169, 1359–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervantes, E.; Saadaoui, E.; Tocino, Á.; Martín-Gómez, J.J. Seed shape quantification in the model legumes: Methods and applications. In The Model Legume Medicago Truncatula; de Brujin, F., Ed.; Wiley: Hoboken, NJ, USA, 2020. [Google Scholar] [CrossRef]
- Cervantes, E.; Martín-Gómez, J.J. Seed Shape Description and Quantification by Comparison with Geometric Models. Horticulturae 2019, 5, 60. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, E.; Martín-Gómez, J.J.; Saadaoui, E. Updated Methods for Seed Shape Analysis. Scientifica 2016, 2016, 5691825. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, E.; Martín Gómez, J.J.; Gutiérrez del Pozo, D.; Silva Dias, L. An Angiosperm Species Dataset Reveals Relationships between Seed Size and Two-Dimensional Shape. Horticulturae 2019, 5, 71. [Google Scholar] [CrossRef] [Green Version]
- Martín-Gómez, J.J.; Rewicz, A.; Rodríguez-Lorenzo, J.L.; Janoušek, B.; Cervantes, E. Seed Morphology in Silene Based on Geometric Models. Plants 2020, 9, 1787. [Google Scholar] [CrossRef] [PubMed]
- Martín-Gómez, J.J.; Del Pozo, D.G.; Ucchesu, M.; Bachetta, G.; De Santamaría, F.C.S.; Tocino, A.; Cervantes, E. Seed Morphology in the Vitaceae Based on Geometric Models. Agronomy 2020, 10, 739. [Google Scholar] [CrossRef]
- Ferreira, T.; Rasband, W. ImageJ User Guide-IJ1.46r. 2012. Available online: http://imagej.nih.gov/ij/docs/guide (accessed on 22 November 2021).
- Zdilla, M.J.; Hatfield, S.A.; McLean, K.A.; Cyrus, L.M.; Laslo, J.M.; Lambert, H.W. Circularity, solidity, axes of a best fit ellipse, aspect ratio, and roundness of the foramen ovale: A morphometric analysis with neurosurgical considerations. J. Craniofac. Surg. 2016, 27, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, E.; Martín-Gómez, J.J.; Espinosa-Roldán, F.E.; Muñoz-Organero, G.; Tocino, Á.; Cabello-Sáenz de Santamaría, F. Seed Morphology in Key Spanish Grapevine Cultivars. Agronomy 2021, 11, 734. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Cervantes, E.; Martín-Gómez, J.J.; Gutiérrez del Pozo, D.; Tocino, Á. Seed Geometry in the Vitaceae. Plants 2021, 10, 1695. [Google Scholar] [CrossRef]
- Ullah, F.; Papini, A.; Shah, S.N.; Zaman, W.; Sohail, A.; Iqbal, M. Seed micromorphology and its taxonomic evidence in subfamily Alsinoideae (Caryophyllaceae). Microsc. Res. Tech. 2019, 82, 250–259. [Google Scholar] [CrossRef]
- Hoseini, E.; Ghahremaninejad, F.; Assadi, M.; Edalatiyan, M.N. Seed micromorphology and its implication in subgeneric classification of Silene (Caryophyllaceae, Sileneae). Flora 2017, 228, 31–38. [Google Scholar] [CrossRef]
- Ghazanfar, S.A. Seed characters as diagnostic in the perennial sections of the genus Silene (Family Caryophyllaceae). Pak. J. Bot. 1983, 15, 7–12. [Google Scholar]
- Dadandi, M.Y.; Yildiz, K. Seed morphology of some Silene L. (Caryophyllaceae) species collected from Turkey. Turk. J. Bot. 2015, 39, 280–297. [Google Scholar] [CrossRef]
- Justice, O.L.; Bass, L.N. Principles and Practices of Seed Storage, Agriculture Handbook, No. 506; U.S. Department of Agriculture: Washington, DC, USA, 1978.
- El-Oqlah, A.A.; Karim, F.M. Morphological and anatomical studies of seed coat in Silene species (Caryophyllaceae) from Jordan. Arab Gulf J. Sci. Res. 1990, 8, 121–139. [Google Scholar]
- Talavera, S. Silene L. In Flora Iberica; Castroviejo, S., Laínz, M., López González, G., Monserrat, P., Muñoz Garmendia, F., Paiva, J., Villar, L., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 1990; Volume 2, pp. 313–406. [Google Scholar]
- Arman, M.; Gholipour, A. Seed morphology diversity in some Iranian endemic Silene (Caryophyllaceae) species and their taxonomic significance. Acta Biol. Szeged. 2013, 57, 31–37. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Population Name | Origin | Subgenus and Section |
|---|---|---|---|
| S. conica L. | Villena | Alicante, Spain | S. subg. Behenantha sect. Conoimorpha |
| S. coutinhoi Rothm. & P.Silva | Larouco | Ourense, Spain | S. subg. Silene sect. Siphonomorpha |
| S. crassipes Fenzl. | Hania | Kriti, Greece | S. subg. Silene sect. Lasiocalycinae |
| S. disticha Willd. | Alcari | Estremadura, Portugal | S. subg. Silene sect. Silene |
| S. diversifolia Otth. | Elda | Alicante, Spain | S. subg. Silene sect. Silene |
| S. foetida Link ex Spreng. | Muiños | Ourense, Spain | S. subg. Behenantha sect. Acutifoliae |
| S. gallica L. | Corse | Corse, France | S. subg. Silene sect. Silene |
| S. latifolia Poir. | Pego | Alicante, Spain | S. subg. Behenantha sect. Melandrium |
| S. littorea Brot. | Melides | Baixo Alentejo, Portugal | S. subg. Behenantha sect. Psammophilae |
| S. mellifera Boiss. & Reut. | Calatayud | Zaragoza, Spain | S. subg. Silene sect. Siphonomorpha |
| S. micropetala Lag. | Nador | Nador, Morocco | S. subg. Silene sect. Silene |
| S. muscipula L. | Granada | Granada, Spain | S. subg. Silene sect. Muscipula |
| S. nicaeensis All. | Benicarló | Castellón, Spain | S. subg. Silene sect. Silene |
| S. nocturna L. | L’Abdet | Alicante, Spain | S. subg. Silene sect. Silene |
| S. portensis L. | Gea | Teruel, Spain | S. subg. Silene sect. Portenses |
| S. scabriflora Brot. | Cavado | Minho, Portugal | S. subg. Silene sect. Silene |
| S. sclerocarpa Dufour | Nador | Nador, Morocco | S. subg. Silene sect. Silene |
| S. stricta L. | Barbate | Cádiz, Spain | S. subg. Silene sect. Muscipula |
| S. tridentata Desf. | Villena | Alicante, Spain | S. subg. Silene sect. Silene |
| S. vivianii Steud. | Saidia | Morocco | S. subg. Silene sect. Silene |
| S. vulgaris (Moench) Garcke | Elda | Alicante, Spain | S. subg. Behenantha sect. Behenantha |
| Species | N | A | P | L | W | AR | C | R |
|---|---|---|---|---|---|---|---|---|
| S. conica | 20 | 0.64 fg | 3.15 f | 0.98 efg | 0.83 efg | 1.18 abcd | 0.81 jk | 0.85 efgh |
| S. coutinhoi | 20 | 0.71 g | 3.46 g | 1.05 g | 0.86 fg | 1.21 bcde | 0.74 def | 0.83 cdef |
| S. crassipes | 20 | 1.02 j | 4.31 k | 1.24 i | 1.04 i | 1.20 abcde | 0.69 ab | 0.84 defgh |
| S. disticha | 20 | 0.26 a | 2.19 a | 0.66 a | 0.51 a | 1.31 fg | 0.70 abc | 0.77 abc |
| S. diversifolia | 20 | 0.57 ef | 3.11 ef | 0.92 def | 0.79 e | 1.17 abc | 0.74 def | 0.86 fgh |
| S. foetida | 20 | 0.86 hi | 3.98 hi | 1.14 h | 0.96 h | 1.18 abcd | 0.69 a | 0.85 efgh |
| S. gallica | 20 | 0.48 cd | 2.88 cd | 0.89 cd | 0.68 c | 1.31 fg | 0.72 cde | 0.77 ab |
| S. latifolia | 20 | 1.17 k | 4.23 jk | 1.32 j | 1.13 j | 1.18 abcd | 0.82 k | 0.85 efgh |
| S. littorea | 20 | 0.58 ef | 2.92 cde | 0.92 de | 0.81 ef | 1.13 ab | 0.86 l | 0.89 gh |
| S. mellifera | 20 | 0.84 h | 3.82 h | 1.20 hi | 0.89 g | 1.37 g | 0.72 bcd | 0.74 a |
| S. micropetala | 20 | 0.94 ij | 3.94 hi | 1.19 hi | 1.00 hi | 1.20 abcde | 0.76 fgh | 0.84 defg |
| S. muscipula | 20 | 0.93 ij | 4.07 ij | 1.19 hi | 1.00 hi | 1.19 abcde | 0.71 abc | 0.84 efgh |
| S. nicaeensis | 20 | 0.56 def | 2.99 def | 0.92 def | 0.77 de | 1.20 abcde | 0.78 hij | 0.84 defg |
| S. nocturna | 20 | 0.51 cde | 2.90 cde | 0.90 cd | 0.72 cd | 1.26 def | 0.76 fgh | 0.80 abcde |
| S. portensis | 20 | 0.69 g | 3.39 g | 0.99 fg | 0.88 g | 1.12 a | 0.75 efg | 0.90 h |
| S. scabriflora | 20 | 0.47 c | 2.73 c | 0.85 c | 0.71 c | 1.21 abcde | 0.79 ijk | 0.83 defg |
| S. sclerocarpa | 20 | 0.37 b | 2.45 b | 0.77 b | 0.61 b | 1.28 efg | 0.77 fghi | 0.79 abcd |
| S. stricta | 20 | 0.28 ab | 2.18 a | 0.67 a | 0.54 a | 1.26 cdef | 0.75 fg | 0.80 bcdef |
| S. tridentata | 20 | 0.29 ab | 2.23 a | 0.68 a | 0.54 a | 1.26 def | 0.74 def | 0.79 abcde |
| S. vivianii | 20 | 0.95 ij | 3.92 hi | 1.19 hi | 1.01 hi | 1.18 abcd | 0.77 ghi | 0.85 efgh |
| S. vulgaris | 20 | 1.97 l | 6.03 l | 1.73 k | 1.44 k | 1.20 abcde | 0.68 a | 0.84 defg |
| Subgenera | N | A | P | L | W | AR | C | R |
|---|---|---|---|---|---|---|---|---|
| S. subg. Behenantha | 100 | 1.04 b (0.52) | 4.06 b (1.12) | 1.22 b (0.30) | 1.03 b (0.24) | 1.18 a (0.05) | 0.77 b (0.08) | 0.85 b (0.04) |
| S. subg. Silene | 320 | 0.62 a (0.26) | 3.16 a (0.71) | 0.96 a (0.20) | 0.78 a (0.18) | 1.23 b (0.11) | 0.74 a (0.04) | 0.82 a (0.07) |
| Species | N | A | P | L | W | AR | C | R |
|---|---|---|---|---|---|---|---|---|
| S. conica | 18 | 0.56 h | 3.04 gh | 1.01 ef | 0.70 hi | 1.43 bcd | 0.76 fg | 0.70 i |
| S. coutinhoi | 16 | 0.56 h | 3.28 ij | 1.09 g | 0.65 fg | 1.69 ef | 0.65 cd | 0.59 ef |
| S. crassipes | 18 | 0.52 gh | 3.70 l | 1.25 h | 0.53 c | 2.41 i | 0.48 ab | 0.43 b |
| S. disticha | 15 | 0.33 abc | 2.43 bc | 0.68 a | 0.61 efg | 1.12 a | 0.70 e | 0.89 l |
| S. diversifolia | 19 | 0.43 defg | 2.80 def | 0.94 cde | 0.58 cdefg | 1.62 def | 0.69 e | 0.62 efgh |
| S. foetida | 19 | 0.76 i | 4.42 n | 1.19 h | 0.81 j | 1.47 cde | 0.49 b | 0.68 hi |
| S. gallica | 15 | 0.39 cdef | 2.68 de | 0.91 cd | 0.55 cdef | 1.66 def | 0.69 e | 0.60 efg |
| S. latifolia | 20 | 1.01 j | 4.04 m | 1.35 i | 0.95 k | 1.43 bcd | 0.77 fg | 0.70 i |
| S. littorea | 19 | 0.45 fg | 2.67 de | 0.94 cde | 0.60 defg | 1.56 def | 0.79 g | 0.64 fghi |
| S. mellifera | 15 | 0.71 i | 3.60 kl | 1.25 h | 0.72 i | 1.77 fg | 0.68 de | 0.58 de |
| S micropetala | 13 | 0.39 bcdef | 3.29 ij | 1.20 h | 0.41 ab | 3.04 j | 0.44 a | 0.34 a |
| S muscipula | 15 | 0.59 h | 3.46 jk | 1.24 h | 0.61 fg | 2.03 h | 0.62 c | 0.50 c |
| S. nicaeensis | 16 | 0.34 abcd | 2.64 cd | 0.96 de | 0.45 b | 2.19 hi | 0.61 c | 0.47 bc |
| S. nocturna | 17 | 0.55 h | 2.99 fgh | 0.92 cd | 0.76 ij | 1.21 ab | 0.77 fg | 0.83 jkl |
| S. portensis | 18 | 0.45 efg | 2.87 efg | 1.06 fg | 0.54 cde | 1.97 gh | 0.68 de | 0.51 cd |
| S. scabriflora | 19 | 0.39 cdef | 2.64 cd | 0.88 c | 0.56 cdef | 1.60 def | 0.70 e | 0.63 efgh |
| S. sclerocarpa | 18 | 0.44 efg | 2.71 de | 0.80 b | 0.70 hi | 1.15 a | 0.75 f | 0.87 kl |
| S. stricta | 17 | 0.30 ab | 2.23 ab | 0.69 a | 0.55 cdef | 1.26 abc | 0.75 fg | 0.80 j |
| S. tridentata | 17 | 0.28 a | 2.16 a | 0.66 a | 0.54 cd | 1.24 abc | 0.75 fg | 0.81 jk |
| S. vivianii | 15 | 0.36 abcde | 3.15 hi | 1.22 h | 0.37 a | 3.32 k | 0.45 ab | 0.30 a |
| S. vulgaris | 16 | 1.67 k | 5.73 o | 1.79 j | 1.19 l | 1.51 de | 0.64 c | 0.66 ghi |
| Subgen. | N | A | P | L | W | AR | C | R |
|---|---|---|---|---|---|---|---|---|
| Behenantha | 92 | 0.87 b (0.43) | 3.93 b (1.08) | 1.24 b (0.30) | 0.84 b (0.20) | 1.48 a (0.09) | 0.69 b (0.12) | 0.68 b (0.04) |
| Silene | 263 | 0.44 a (0.13) | 2.90 a (0.48) | 0.98 a (0.21) | 0.57 a (0.11) | 1.80 b (0.65) | 0.66 a (0.11) | 0.62 a (0.19) |
| Species | N | Mean J Index Cardioid (Standard Deviation) | Min | Max | J Index Value in Average Silhouette |
|---|---|---|---|---|---|
| S. conica | 20 | 92.5 k (1,18) | 90.0 | 94.4 | 94.2 |
| S. coutinhoi | 20 | 86.8 bcdef (1.81) | 83.3 | 89.5 | 88.3 |
| S. crassipes | 20 | 87.3 cdef (3.04) | 80.0 | 91.5 | 90.1 |
| S. disticha | 20 | 83.7 ab (4.26) | 70.8 | 88.1 | 88.0 |
| S. diversifolia | 20 | 89.7 efghijk (1.62) | 85.9 | 91.9 | 91.6 |
| S. foetida | 20 | 89.8 fghijk (1.10) | 87.8 | 91.3 | 92.6 |
| S. gallica | 20 | 84.7 bc (2.82) | 78.9 | 89.4 | 87.3 |
| S. latifolia | 20 | 89.7 efghijk (1.55) | 86.4 | 93.2 | 91.1 |
| S. littorea | 20 | 92.0 jk (2.07) | 86.7 | 94.7 | 93.3 |
| S. mellifera | 20 | 80.9 a (6.07) | 70.4 | 89.0 | 92.9 |
| S. micropetala | 20 | 88.7 defghi (2.89) | 80.3 | 91.7 | 91.7 |
| S. muscipula | 20 | 86.6 bcde (3.18) | 79.8 | 90.4 | 90.2 |
| S. nicaeensis | 20 | 87.4 cdefg (3.16) | 81.4 | 92.7 | 89.6 |
| S. nocturna | 20 | 86.3 bcd (4.14) | 76.1 | 91.5 | 94.9 |
| S. portensis | 20 | 90.8 hijk (0.85) | 88.8 | 92.1 | 90.3 |
| S. scabriflora | 20 | 89.1 defghij (2.10) | 84.1 | 93.3 | 90.6 |
| S. sclerocarpa | 20 | 87.9 defgh (1.98) | 84.3 | 92.2 | 89.8 |
| S. stricta | 20 | 87.5 cdefg (2.88) | 82.6 | 91.3 | 90.1 |
| S. tridentata | 20 | 88.5 defgh (2.27) | 82.8 | 91.6 | 89.1 |
| S. vivianii | 20 | 90.5 ghijk (1.36) | 86.4 | 92.3 | 90.4 |
| S. vulgaris | 20 | 88.6 defghi (2.44) | 82.1 | 91.9 | 92.2 |
| Models/Species | Model 2 | Model 3 | Model 4 | Model 5 | Model 6 | Model 7 | Model 8 |
|---|---|---|---|---|---|---|---|
| S. conica | 91.0 hi 94.3 | 86.0 bcdef 88.0 | 89.2 efgh 92.1 | 88.6 defg 91.2 | 84.1 cdefg 87.3 | 87.7 bcdef 90.4 | 88.6 cdef 90.1 |
| S. coutinhoi | 84.7 bcde 87.4 | 84.5 abcd 90.0 | 83.2 b 87.4 | 86.3 cde 91.4 | 83.4 bcdef 87.4 | 87.2 bcdef 92.6 | 86.3 cd 91.2 |
| S. crassipes | 84.2 abc 90.9 | 82.8 ab 88.0 | 83.9 bc 87.7 | 84.1 abc 91.9 | 80.3 ab 84.9 | 85.8 bc 92.0 | 86.3 cd 90.6 |
| S. disticha | 82.8 ab 87.8 | 83.8 abc 91.3 | 79.2 a 92.2 | 82.3 a 91.2 | 84.1 cdefg 90.1 | 81.3 a 88.8 | 81.2 a 88.3 |
| S. diversifolia | 89.9 ghi 91.4 | 84.8 abcd 88.9 | 86.2 bcde 87.3 | 91.0 g 93.3 | 83.4 bcdef 84.4 | 90.4 f 93.6 | 87.3 cde 91.4 |
| S. foetida | 89.8 ghi 94.3 | 84.4 abcd 87.7 | 89.7 gh 93.2 | 87.8 defg 90.6 | 81.1 abcd 85.6 | 85.3 bc 88.3 | 88.1 cdef 89.8 |
| S. gallica | 84.3 abcd 86.6 | 88.4 efg 93.4 | 87.2 defg 88.9 | 88.1 defg 92.0 | 86.5 fg 92.0 | 84.9 b 89.5 | 85.5 bc 88.5 |
| S. latifolia | 91.1 hi 91.3 | 85.6 abcdef 86.7 | 90.8 h 92.5 | 87.8 defg 88.2 | 82.3 abcde 86.4 | 85.8 bc 85.3 | 87.1 cde 86.9 |
| S. littorea | 92.4 i 95.1 | 84.2 abcd 85.7 | 89.2 efgh 90.4 | 88.5 defg 89.9 | 80.5 abc 81.0 | 89.4 ef 91.1 | 91.4 f 91.3 |
| S. mellifera | 81.6 a 85.2 | 82.6 a 91.4 | 83.5 b 89.4 | 82.8 ab 89.5 | 82.7 abcde 92.0 | 81.4 a 87.3 | 82.3 ab 85.3 |
| S. micropetala | 88.9 fgh 92.0 | 85.2 abcde 89.6 | 87.7 defgh 90.3 | 88.7 defg 92.4 | 81.8 abcde 87.8 | 88.5 cdef 91.7 | 87.9 cdef 88.6 |
| S. muscipula | 87.3 defg 90.4 | 83.6 abc 86.7 | 85.5 bcd 89.3 | 85.5 bcd 89.1 | 80.0 ab 83.9 | 85.8 bcd 88.7 | 87.4 cde 93.3 |
| S. nicaeensis | 87.9 fg 89.8 | 86.2 cdef 88.1 | 86.9 cdefg 89.9 | 88.3 defg 90.9 | 83.2 abcdef 85.5 | 88.4 cdef 89.3 | 90.1 ef 91.0 |
| S. nocturna | 86.3 cdef 90.3 | 86.8 cdefg 92.2 | 87.1 cdefg 90.4 | 88.3 defg 94.3 | 85.3 efg 89.7 | 87.6 bcdef 91.9 | 87.4 cde 90.3 |
| S. portensis | 90.2 ghi 90.4 | 84.3 abcd 84.5 | 86.4 bcdef 88.4 | 87.6 def 89.1 | 79.7 a 80.9 | 90.0 ef 90.8 | 90.4 ef 92.7 |
| S. scabriflora | 88.6 fgh 89.8 | 87.2 defg 88.3 | 88.1 defgh 89.0 | 89.1 efg 91.2 | 84.0 cdefg 84.5 | 89.8 ef 91.4 | 91.3 f 94.1 |
| S. sclerocarpa | 87.6 efg 88.8 | 88.6 fg 92.1 | 89.6 gh 92.2 | 89.3 efg 93.2 | 86.9 fg 90.2 | 88.3 bcdef 91.2 | 87.6 cde 88.9 |
| S. stricta | 87.3 defg 87.9 | 86.6 cdefg 91.5 | 88.3 defgh 90.9 | 87.8 defg 92.8 | 84.4 defg 89.8 | 86.9 bcde 90.2 | 85.7 c 88.5 |
| S. tridentata | 88.3 fgh 89.6 | 89.7 g 92.9 | 89.5 fgh 90.8 | 90.8 fg 93.2 | 87.5 g 90.2 | 89.5 ef 90.5 | 88.9 cdef 88.3 |
| S. viviani | 90.1 ghi 90.1 | 86.0 bcdef 85.1 | 87.7 defgh 87.4 | 89.4 efg 86.3 | 82.4 abcde 83.4 | 89.2 def 89.7 | 89.8 def 91.9 |
| S. vulgaris | 89.3 ghi 92.6 | 84.9 abcd 87.8 | 88.8 efgh 93.0 | 86.6 cde 90.4 | 82.8 abcde 86.0 | 85.1 bc 87.4 | 87.6 cde 89.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juan, A.; Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Janoušek, B.; Cervantes, E. New Techniques for Seed Shape Description in Silene Species. Taxonomy 2022, 2, 1-19. https://doi.org/10.3390/taxonomy2010001
Juan A, Martín-Gómez JJ, Rodríguez-Lorenzo JL, Janoušek B, Cervantes E. New Techniques for Seed Shape Description in Silene Species. Taxonomy. 2022; 2(1):1-19. https://doi.org/10.3390/taxonomy2010001
Chicago/Turabian StyleJuan, Ana, José Javier Martín-Gómez, José Luis Rodríguez-Lorenzo, Bohuslav Janoušek, and Emilio Cervantes. 2022. "New Techniques for Seed Shape Description in Silene Species" Taxonomy 2, no. 1: 1-19. https://doi.org/10.3390/taxonomy2010001
APA StyleJuan, A., Martín-Gómez, J. J., Rodríguez-Lorenzo, J. L., Janoušek, B., & Cervantes, E. (2022). New Techniques for Seed Shape Description in Silene Species. Taxonomy, 2(1), 1-19. https://doi.org/10.3390/taxonomy2010001