
Shugoshin: From the Perspective of Clinical Disorders


Abstract
1. Introduction
2. Shugoshin Background
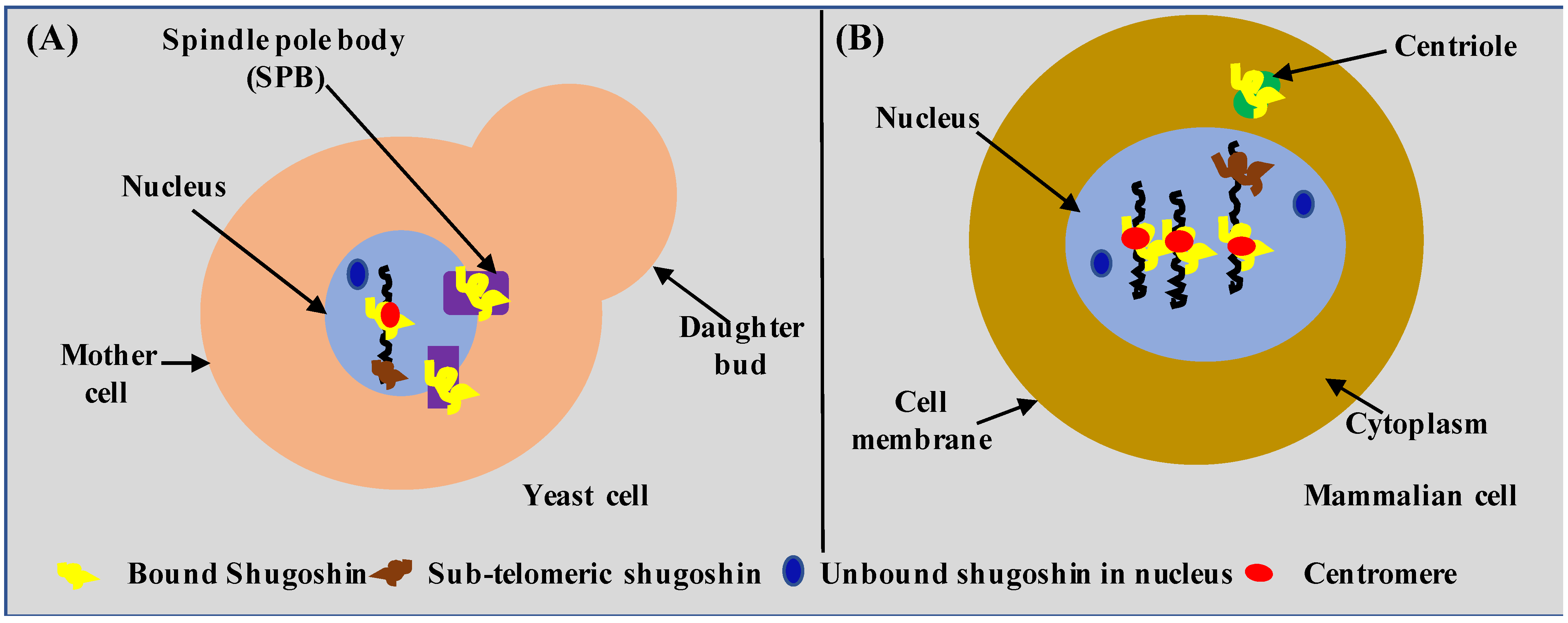
3. Cellular Localization
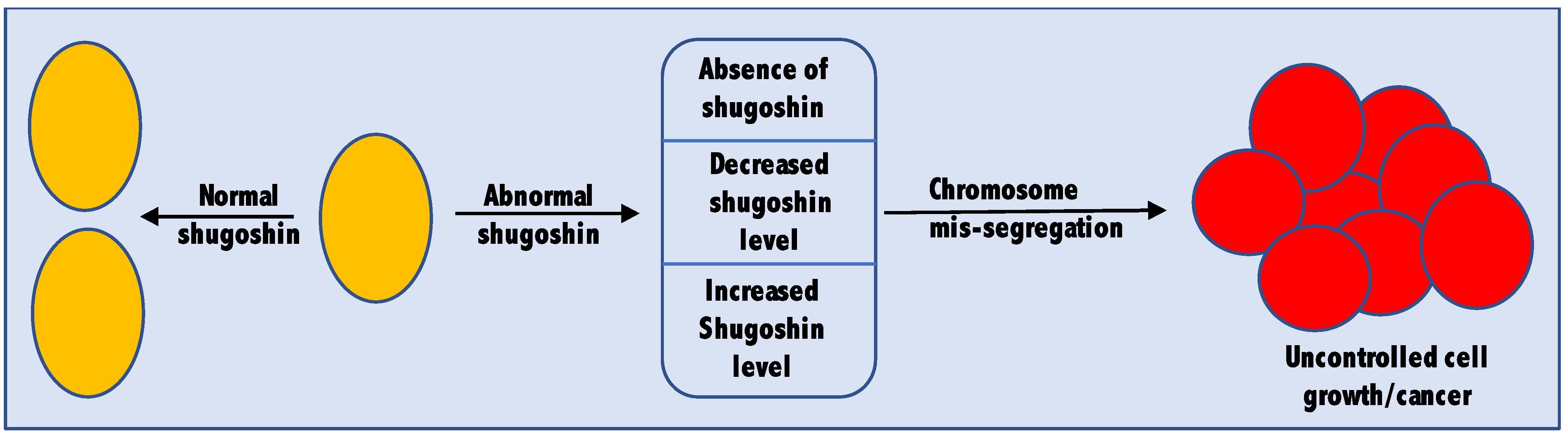
4. Shugoshin as a Tumor-Associated Gene
4.1. Shugoshin as a Tumor Suppressor Gene
4.2. Shugoshin as an Oncogene
5. Shugoshin in Other Clinical Disorders
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rajagopalan, H.; Lengauer, C. Aneuploidy and cancer. Nature 2004, 432, 338–341. [Google Scholar] [CrossRef]
- Hassold, T.; Hall, H.; Hunt, P. The origin of human aneuploidy: Where we have been, where we are going. Hum. Mol. Genet. 2007, 16, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Jallepalli, P.V.; Lengauer, C. Chromosome segregation and cancer: Cutting through the mystery. Nat. Rev. Cancer 2001, 1, 109–117. [Google Scholar] [CrossRef]
- Lengauer, C.; Kinzler, K.W.; Vogelstein, B. Genetic instability in colorectal cancers. Nature 1997, 386, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.K. The incidence, origin, and etiology of aneuploidy. Int. Rev. Cytol. 1996, 167, 263–296. [Google Scholar]
- Sen, S. Aneuploidy and cancer. Curr. Opin. Oncol. 2000, 12, 82–88. [Google Scholar] [CrossRef]
- Uhlmann, F. Secured cutting: Controlling separase at the metaphase to anaphase transition. EMBO Rep. 2001, 2, 487–492. [Google Scholar] [CrossRef]
- Kumar, R. Separase: Function Beyond Cohesion Cleavage and an Emerging Oncogene. J. Cell Biochem. 2017, 118, 1283–1299. [Google Scholar] [CrossRef] [PubMed]
- Mehta, G.D.; Kumar, R.; Srivastava, S.; Ghosh, S.K. Cohesin: Functions beyond sister chromatid cohesion. FEBS Lett. 2013, 587, 2299–2312. [Google Scholar] [CrossRef] [PubMed]
- Nasmyth, K.; Haering, C.H. Cohesin: Its roles and mechanisms. Annu. Rev. Genet. 2009, 43, 525–558. [Google Scholar] [CrossRef] [PubMed]
- Joglekar, A.P. A Cell Biological Perspective on Past, Present and Future Investigations of the Spindle Assembly Checkpoint. Biology 2016, 5, 44. [Google Scholar] [CrossRef]
- Kamenz, J.; Hauf, S. Time to Split up: Dynamics of Chromosome Separation. Trends Cell Biol. 2017, 27, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Yoshino, Y.; Chiba, N. Regulation of the centrosome cycle. Mol. Cell Oncol. 2015, 3, e1075643. [Google Scholar] [CrossRef] [PubMed]
- Simpson-Lavy, K.J.; Oren, Y.S.; Feine, O.; Sajman, J.; Listovsky, T.; Brandeis, M. Fifteen years of APC/cyclosome: A short and impressive biography. Biochem. Soc. Trans. 2010, 38, 78–82. [Google Scholar] [CrossRef][Green Version]
- Agarwal, M.; Mehta, G.; Ghosh, S.K. Role of Ctf3 and COMA subcomplexes in meiosis: Implication in maintaining Cse4 at the centromere and numeric spindle poles. Biochim. Biophys. Acta Mol. Cell Res. 2015, 1853, 671–684. [Google Scholar] [CrossRef]
- Agarwal, M.; Jin, H.; McClain, M.; Fan, J.; Koch, B.A.; Jaspersen, S.L.; Yu, H.G. The half-bridge component Kar1 promotes centrosome separation and duplication during budding yeast meiosis. Mol. Biol. Cell 2018, 29, 1798–1810. [Google Scholar] [CrossRef] [PubMed]
- Marston, A.L. Shugoshins: Tension-sensitive pericentromeric adaptors safeguarding chromosome segregation. Mol. Cell Biol. 2015, 35, 634–648. [Google Scholar] [CrossRef]
- Clift, D.; Marston, A.L. The role of shugoshin in meiotic chromosome segregation. Cytogenet. Genome Res. 2011, 133, 234–242. [Google Scholar] [CrossRef]
- Wassmann, K. Sister chromatid segregation in meiosis II: Deprotection through phosphorylation. Cell Cycle 2013, 12, 1352–1359. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, H. Functioning mechanisms of Shugoshin-1 in centromeric cohesion during mitosis. Essays Biochem. 2020, 64, 289–297. [Google Scholar]
- Xu, Z.; Cetin, B.; Anger, M.; Cho, U.S.; Helmhart, W.; Nasmyth, K.; Xu, W. Structure and function of the PP2A-shugoshin inter-action. Mol. Cell 2009, 35, 426–441. [Google Scholar] [CrossRef] [PubMed]
- Swaney, D.L.; Beltrao, P.; Starita, L.; Guo, A.; Rush, J.; Fields, S.; Krogan, N.J.; Villén, J. Global analysis of phosphorylation and ubiquitylation crosstalk in protein degradation. Nat. Methods 2013, 10, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.T.; Bickel, S.E.; Young, L.M.; Orr-Weaver, T.L. Maintenance of sister-chromatid cohesion at the centromere by the Drosophila MEI-S332 protein. Genes Dev. 1998, 12, 3843–3856. [Google Scholar] [CrossRef]
- Watanabe, Y. Shugoshin: Guardian spirit at the centromere. Curr. Opin. Cell Biol. 2005, 17, 590–595. [Google Scholar] [CrossRef]
- Watanabe, Y.; Kitajima, T.S. Shugoshin protects cohesin complexes at centromeres. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 515–521. [Google Scholar] [CrossRef]
- Watanabe, Y. Temporal and spatial regulation of targeting aurora B to the inner centromere. Cold Spring Harb. Symp. Quant. Biol. 2010, 75, 419–423. [Google Scholar] [CrossRef]
- Davis, B.K. A analysis of a meiotic mutant resulting in precocious sister-centromere separation in Drosophila melanogaster. Mol. Gen. Genet. 1971, 113, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Kerrebrock, A.W.; Miyazaki, W.Y.; Birnby, D.; Orr-Weaver, T.L. The Drosophila mei-S332 gene promotes sister-chromatid cohesion in meiosis following kinetochore differentiation. Genetics 1992, 130, 827–841. [Google Scholar] [CrossRef]
- Kerrebrock, A.; Moore, D.; Wu, J.; Orr-Weaver, T. Mei-S332, a Drosophila protein required for sister-chromatid cohesion, can localize to meiotic centromere regions. Cell 1995, 83, 247–256. [Google Scholar] [CrossRef]
- Kitajima, T.S.; Kawashima, S.A.; Watanabe, Y. The conserved kinetochore protein shugoshin protects centromeric cohesion during meiosis. Nature 2004, 427, 510–517. [Google Scholar] [CrossRef]
- Katis, V.L.; Galova, M.; Rabitsch, K.P.; Gregan, J.; Nasmyth, K. Maintenance of cohesin at centromeres after meiosis I in budding yeast requires a kinetochore-associated protein related to MEI-S332. Curr. Biol. 2004, 14, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Marston, A.L.; Tham, W.H.; Shah, H.; Amon, A. A genome-wide screen identifies genes required for centromeric cohesion. Science 2004, 303, 1367–1370. [Google Scholar] [CrossRef] [PubMed]
- Rabitsch, K.P.; Gregan, J.; Schleiffer, A.; Javerzat, J.P.; Eisenhaber, F.; Nasmyth, K. Two fission yeast homologs of Drosophila Mei-S332 are required for chromosome segregation during meiosis-I and II. Curr. Biol. 2004, 14, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Hamant, O.; Golubovskaya, I.; Meeley, R.; Fiume, E.; Timofejeva, L.; Schleiffer, A.; Nasmyth, K.; Cande, W.Z. A REC8-dependent plant Shugoshin is required for maintenance of centromeric cohesion during meiosis and has no mitotic functions. Curr. Biol. 2005, 15, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tang, D.; Wang, K.; Shen, Y.; Qin, B.; Miao, C.; Li, M.; Cheng, Z. OsSGO1 maintains synaptonemal complex stabilization in addition to protecting centromeric cohesion during rice meiosis. Plant J. 2011, 67, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Dai, W. Shugoshins function as a guardian for chromosomal stability in nuclear division. Cell Cycle 2012, 11, 2631–2642. [Google Scholar] [CrossRef]
- Llano, E.; Gómez, R.; Gutiérrez-Caballero, C.; Herrán, Y.; Sánchez-Martín, M.; Vázquez-Quiñones, L.E.; Hernández, T.; de Álava, E.; Cuadrado, A.; Barbero, J.; et al. Shugoshin-2 is essential for the completion of meiosis but not for mitotic cell division in mice. Genes Dev. 2008, 22, 2400–2413. [Google Scholar] [CrossRef]
- de Carvalho, C.E.; Zaaijer, S.; Smolikov, S.; Gu, Y.; Schumacher, J.M.; Colaiácovo, M.P. LAB-1 antagonizes the Aurora B kinase in C. elegans. Genes Dev. 2008, 22, 2869–2885. [Google Scholar] [CrossRef]
- Huh, W.K.; Falvo, J.V.; Gerke, L.C.; Carroll, A.S.; Howson, R.W.; Weissman, J.S.; O’Shea, E.K. Global analysis of protein localization in budding yeast. Nature 2003, 425, 686–691. [Google Scholar] [CrossRef]
- Kiburz, B.M.; Reynolds, D.B.; Megee, P.C.; Marston, A.L.; Lee, B.H.; Lee, T.I.; Levine, S.S.; Young, R.A.; Amon, A. The core centromere and Sgo1 establish a 50-kb cohesin-protected domain around centromeres during meiosis I. Genes Dev. 2005, 19, 3017–3030. [Google Scholar] [CrossRef][Green Version]
- Mishra, P.K.; Thapa, K.S.; Chen, P.; Wang, S.; Hazbun, T.R.; Basrai, M.A. Budding yeast CENP-ACse4 interacts with the N-terminus of Sgo1 and regulates its association with centromeric chromatin. Cell Cycle 2018, 17, 11–23. [Google Scholar] [CrossRef]
- Tashiro, S.; Handa, T.; Matsuda, A.; Ban, T.; Takigawa, T.; Miyasato, K.; Ishii, K.; Kugou, K.; Ohta, K.; Hiraoka, Y.; et al. Shugoshin forms a specialized chromatin domain at subtelomeres that regulates transcription and replication timing. Nat. Commun. 2016, 7, 10393. [Google Scholar] [CrossRef] [PubMed]
- Kanoh, J. Unexpected roles of a shugoshin protein at subtelomeres. Genes Genet. Syst. 2018, 92, 127–133. [Google Scholar] [CrossRef]
- McGuinness, B.E.; Hirota, T.; Kudo, N.R.; Peters, J.M.; Nasmyth, K. Shugoshin prevents dissociation of cohesin from centromeres during mitosis in vertebrate cells. PLoS Biol. 2005, 3, e86. [Google Scholar] [CrossRef]
- Karamysheva, Z.; Diaz-Martinez, L.A.; Crow, S.E.; Li, B.; Yu, H. Multiple anaphase-promoting complex/cyclosome degrons mediate the degradation of human Sgo1. J. Biol. Chem. 2009, 284, 1772–1780. [Google Scholar] [CrossRef]
- Mu, J.; Fan, L.; Liu, D.; Zhu, D. Overexpression of shugoshin1 predicts a poor prognosis for prostate cancer and promotes metastasis by affecting epithelial-mesenchymal transition. Onco Targets Ther. 2019, 12, 1111–1118. [Google Scholar] [CrossRef]
- Coon, S.W.; Savera, A.T.; Zarbo, R.J.; Benninger, M.S.; Chase, G.A.; Rybicki, B.A.; Van Dyke, D.L. Prognostic implications of loss of heterozygosity at 8p21 and 9p21 in head and neck squamous cell carcinoma. Int. J. Cancer 2004, 111, 206–212. [Google Scholar] [CrossRef]
- Shao, J.Y.; Wang, H.Y.; Huang, X.M.; Feng, Q.S.; Huang, P.; Feng, B.J.; Huang, L.X.; Yu, X.J.; Li, J.T.; Hu, L.F.; et al. Genome-wide allele type analysis of sporadic primary nasopharyngeal carcinoma from southern China. Int. J. Oncol. 2000, 17, 1267–1275. [Google Scholar]
- Altura, R.A.; Maris, J.M.; Li, H.; Boyett, J.M.; Brodeur, G.M.; Look, A.T. Novel regions of chromosomal loss in familial neuroblastoma by comparative genomic hybridization. Genes Chromosomes Cancer 1997, 19, 176–184. [Google Scholar] [CrossRef]
- Pallai, R.; Bhaskar, A.; Barnett-Bernodat, N.; Gallo-Ebert, C.; Nickels, J.T., Jr.; Rice, L.M. Cancerous inhibitor of protein phosphatase 2A promotes premature chromosome segregation and aneuploidy in prostate cancer cells through association with shugoshin. Tumor Biol. 2015, 36, 6067–6074. [Google Scholar] [CrossRef]
- Dahiya, R.; McCarville, J.; Hu, W.; Lee, C.; Chui, R.M.; Kaur, G.; Deng, G. Chromosome 3p24–26 and 3p22–12 loss in human prostatic adenocarcinoma. Int. J. Cancer 1997, 71, 20–25. [Google Scholar] [CrossRef]
- Beder, L.B.; Gunduz, M.; Ouchida, M.; Fukushima, K.; Gunduz, E.; Ito, S.; Sakai, A.; Nagai, N.; Nishizaki, K.; Shimizu, K. Genome-wide analyses on loss of heterozygosity in head and neck squamous cell carcinomas. Lab. Investig. 2003, 83, 99–105. [Google Scholar] [CrossRef]
- Kohno, T.; Morishita, K.; Takano, H.; Shapiro, D.N.; Yokota, J. Homozygous deletion at chromosome 2q33 in human small-cell lung carcinoma identified by arbitrarily primed PCR genomic fingerprinting. Oncogene 1994, 9, 103–108. [Google Scholar]
- Rader, J.S.; Kamarasova, T.; Huettner, P.C.; Li, L.; Li, Y.; Gerhard, D.S. Allelotyping of all chromosomal arms in invasive cervical cancer. Oncogene 1996, 13, 2737–2741. [Google Scholar]
- Takita, J.; Yang, H.W.; Chen, Y.Y.; Hanada, R.; Yamamoto, K.; Teitz, T.; Kidd, V.; Hayashi, Y. Allelic imbalance on chromosome 2q and alterations of the caspase 8 gene in neuroblastoma. Oncogene 2001, 20, 4424–4432. [Google Scholar] [CrossRef][Green Version]
- Iwaizumi, M.; Shinmura, K.; Mori, H.; Yamada, H.; Suzuki, M.; Kitayama, Y.; Igarashi, H.; Nakamura, T.; Suzuki, H.; Watanabe, Y.; et al. Human Sgo1 downregulation leads to chromosomal instability in colorectal cancer. Gut 2009, 58, 249–260. [Google Scholar] [CrossRef]
- Rao, C.V.; Sanghera, S.; Zhang, Y.; Biddick, L.; Reddy, A.; Lightfoot, S.; Janakiram, N.B.; Mohammed, A.; Dai, W.; Yamada, H.Y. Systemic Chromosome Instability Resulted in Colonic Transcriptomic Changes in Metabolic, Proliferation, and Stem Cell Regulators in Sgo1-/+ Mice. Cancer Res. 2016, 76, 630–642. [Google Scholar] [CrossRef]
- Rao, C.V.; Sanghera, S.; Zhang, Y.; Biddick, L.; Reddy, A.; Lightfoot, S.; Dai, W.; Yamada, H.Y. Antagonizing pathways leading to differential dynamics in colon carcinogenesis in Shugoshin1 (Sgo1)-haploinsufficient chromosome instability model. Mol. Carcinog. 2016, 55, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.Y.; Zhang, Y.; Reddy, A.; Mohammed, A.; Lightfoot, S.; Dai, W.; Rao, C.V. Tumor-promoting/progressing role of additional chromosome instability in hepatic carcinogenesis in Sgo1 (Shugoshin 1) haploinsufficient mice. Carcinogenesis 2015, 36, 429–440. [Google Scholar] [CrossRef]
- Yamada, H.Y.; Yao, Y.; Wang, X.; Zhang, Y.; Huang, Y.; Dai, W.; Rao, C.V. Haploinsufficiency of SGO1 results in deregulated centrosome dynamics, enhanced chromosomal instability and colon tumorigenesis. Cell Cycle 2012, 11, 479–488. [Google Scholar] [CrossRef]
- Kahyo, T.; Iwaizumi, M.; Shinmura, K.; Matsuura, S.; Nakamura, T.; Watanabe, Y.; Yamada, H.; Sugimura, H. A novel tumor-derived SGOL1 variant causes abnormal mitosis and unstable chromatid cohesion. Oncogene 2011, 30, 4453–4463. [Google Scholar] [CrossRef]
- Yamada, H.Y.; Kumar, G.; Zhang, Y.; Rubin, E.; Lightfoot, S.; Dai, W.; Rao, C.V. Systemic chromosome instability in Shugosh-in-1 mice resulted in compromised glutathione pathway, activation of Wnt signaling and defects in immune system in the lung. Oncogenesis 2016, 5, e256. [Google Scholar] [CrossRef]
- Wang, L.H.; Yen, C.J.; Li, T.N.; Elowe, S.; Wang, W.C.; Wang, L.H. Sgo1 is a potential therapeutic target for hepatocellular carcinoma. Oncotarget 2015, 6, 2023–2033. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ikezoe, T.; Nishioka, C.; Yokoyama, A. A novel treatment strategy targeting shugoshin 1 in hematological malignancies. Leuk. Res. 2013, 37, 76–82. [Google Scholar] [CrossRef]
- Scanlan, M.J.; Gout, I.; Gordon, C.M.; Williamson, B.; Stocker Et Gure, A.O.; Jager, D.; Chen, Y.T.; Mackay, A.; O’Hare, M.J.; Old, L.J. Humoral immunity to human breast cancer: Antigen definition and quantitative analysis of mRNA expression. Cancer Immun. 2001, 1, 4. [Google Scholar] [PubMed]
- Yang, Q.; Yoshimura, G.; Nakamura, M.; Nakamura, Y.; Shan, L.; Suzuma, T.; Tamaki, T.; Umemura, T.; Mori, I.; Kakudo, K. Allelic loss of chromosome 3p24 correlates with tumor progression rather than with retinoic acid receptor beta2 expression in breast carcinoma. Breast Cancer Res. Treat. 2011, 70, 39–45. [Google Scholar] [CrossRef]
- Matsuura, S.; Kahyo, T.; Shinmura, K.; Iwaizumi, M.; Yamada, H.; Funai, K.; Kobayashi, J.; Tanahashi, M.; Niwa, H.; Ogawa, H.; et al. SGOL1 variant B induces abnormal mitosis and resistance to taxane in non-small cell lung cancers. Sci. Rep. 2013, 3, 3012. [Google Scholar] [CrossRef]
- Wong, W.K.; Kelly, T.; Li, J.; Ma, H.T.; Poon, R.Y. SGO1C is a non-functional isoform of Shugoshin and can disrupt sister chromatid cohesion by interacting with PP2A-B56. Cell Cycle 2015, 14, 3965–3977. [Google Scholar] [CrossRef]
- Chen, Q.; Wan, X.; Chen, Y.; Liu, C.; Gu, M.; Wang, Z. SGO1 induces proliferation and metastasis of prostate cancer through AKT-mediated signaling pathway. Am. J. Cancer Res. 2019, 9, 2693–2705. [Google Scholar] [PubMed]
- Ribeiro-Varandas, E.; Viegas, W.; Sofia Pereira, H.; Delgado, M. Bisphenol A at concentrations found in human serum induces aneugenic effects in endothelial cells. Mutat. Res. 2013, 751, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Ruiz, M.; García-Martínez, A.; Corral-Juan, M.; Pérez-Álvarez, Á.I.; Plasencia, A.M.; Villamar, M.; Moreno-Pelayo, M.A.; Matilla-Dueñas, A.; Menéndez-González, M.; Del Castillo, I. Perrault syndrome with neurological features in a compound heterozygote for two TWNK mutations: Overlap of TWNK-related recessive disorders. J. Transl. Med. 2019, 17, 290. [Google Scholar] [CrossRef]
- Faridi, R.; Rehman, A.U.; Morell, R.J.; Friedman, P.L.; Demain, L.; Zahra, S.; Khan, A.A.; Tohlob, D.; Assir, M.Z.; Beaman Khan, S.N.; et al. Mutations of SGO2 and CLDN14 collectively cause coincidental Perrault syndrome. Clin. Genet. 2017, 91, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.V.; Farooqui, M.; Zhang, Y.; Asch, A.S.; Yamada, H.Y. Spontaneous development of Alzheimer’s disease-associated brain pathology in a Shugoshin-1 mouse cohesinopathy model. Aging Cell 2018, 17, e12797. [Google Scholar] [CrossRef]
- Rao, C.V.; Farooqui, M.; Madhavaram, A.; Zhang, Y.; Asch, A.S.; Yamada, H.Y. GSK3-ARC/Arg3.1 and GSK3-Wnt signaling axes trigger amyloid-β accumulation and neuroinflammation in middle-aged Shugoshin 1 mice. Aging Cell 2020, 19, e13221. [Google Scholar] [CrossRef]
- Chetaille, P.; Preuss, C.; Burkhard, S.; Côté, J.-M.; Houde, C.; Castilloux, J.; Piché, J.; Gosset, N.; Leclerc, S.; Wünnemann, F.; et al. Mutations in SGOL1 cause a novel cohesinopathy affecting heart and gut rhythm. Nat. Genet. 2014, 46, 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Dai, W. Shugoshin, a guardian for sister chromatid segregation. Exp. Cell Res. 2005, 310, 1–9. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | No. of Genes | Kingdom | References |
|---|---|---|---|
| Saccharomyces cerevisiae | 1 | Fungi | * |
| Schizosaccharomyces pombe | 2 | Fungi | * |
| Mus musculus | 2 | Animalia | Uniprot |
| Arabidopsis thialana | 2 | Plantae | Uniprot |
| §Homo sapiens | 2 | Animalia | Uniprot |
| Drosophila melanogaster | 1 | Animalia | Uniprot |
| Caenorhabditis elegence | 1 | Animalia | Uniprot |
| Oryza sativa | 1 | Plantae | Uniprot |
| Xenopus laevis | 1 | Animalia | Uniprot |
| Neurospora crassa | 1 | Fungi | Uniprot |
| Danio rerio | 1 | Animalia | Uniprot |
| Zea mays | 1 | Plantae | Uniprot |
| Rattus norvegicus | 1 | Animalia | Uniprot |
| Candida glabrata | 1 | Fungi | * |
| Kluyveromyces lactis | 1 | Fungi | * |
| Aphis gossypii | 1 | Fungi | * |
| Pristionchus pacificus | 1 | Animalia | Uniprot |
| Oryzias latipes | 1 | Animalia | Uniprot |
| Candida albicans | 1 | Fungi | * |
| Shugoshin | Isoform | Number of Amino Acid Residues | Mol. Mass (in kDa) | Identifier |
|---|---|---|---|---|
| * SGO1 | Isoform 1 | 561 | 64.19 | Q5FBB7-1 |
| Isoform 2 | 309 | 35.344 | Q5FBB7-2 | |
| Isoform 3 | 292 | 33.501 | Q5FBB7-3 | |
| Isoform 4 | 275 | 31.276 | Q5FBB7-4 | |
| Isoform 5 | 258 | 29.433 | Q5FBB7-5 | |
| Isoform 6 | 527 | 60.122 | Q5FBB7-6 | |
| Isoform 7 | 215 | 24.646 | Q5FBB7-7 | |
| ! SGO2 | Isoform 1 | 1265 | 144.739 | Q562F6-1 |
| Isoform 2 | 1261 | 144.181 | Q562F6-2 | |
| Isoform 3 | 247 | 28.23 | Q562F6-3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, R.; Agarwal, M. Shugoshin: From the Perspective of Clinical Disorders. BioChem 2021, 1, 51-59. https://doi.org/10.3390/biochem1020006
Kumar R, Agarwal M. Shugoshin: From the Perspective of Clinical Disorders. BioChem. 2021; 1(2):51-59. https://doi.org/10.3390/biochem1020006
Chicago/Turabian StyleKumar, Ravinder, and Meenakshi Agarwal. 2021. "Shugoshin: From the Perspective of Clinical Disorders" BioChem 1, no. 2: 51-59. https://doi.org/10.3390/biochem1020006
APA StyleKumar, R., & Agarwal, M. (2021). Shugoshin: From the Perspective of Clinical Disorders. BioChem, 1(2), 51-59. https://doi.org/10.3390/biochem1020006