Abstract
Myeloid/lymphoid neoplasms with tyrosine kinase gene fusions (MLN-TK) represent a distinct group of hematologic malignancies recognized in the latest WHO classification due to shared clinical, morphological, and molecular features, and their responsiveness to tyrosine kinase inhibitors (TKIs). Among these, fusions involving the SYK gene, such as ETV6::SYK and ITK::SYK, have emerged as rare but potentially targetable genetic events in both myeloid and lymphoid neoplasms. SYK, a non-receptor tyrosine kinase critical for hematopoietic signalling, can become constitutively activated through gene fusions, driving oncogenesis via the PI3K/AKT, MAPK, and JAK-STAT pathways. ETV6::SYK has been primarily associated with myeloid neoplasms, often presenting with eosinophilia, bone marrow dysplasia, and skin involvement. In vitro and in vivo models confirm its leukemogenic potential and identify SYK as a therapeutic target. Although SYK inhibitors like fostamatinib have shown transient efficacy, resistance mechanisms, possibly involving alternative pathway activation, remain a challenge. The ITK::SYK fusion, on the other hand, has been identified in peripheral T-cell lymphomas, particularly of the follicular helper T-cell subtype, with similar pathway activation and potential for targeted intervention. Additional rare SYK fusions, such as PML::SYK and CTLC::SYK, have been reported in myeloid neoplasms and juvenile xanthogranuloma, respectively, expanding the spectrum of SYK-driven diseases. Accumulating evidence supports the inclusion of SYK fusions in future classification systems and highlights the need for broader molecular screening and clinical evaluation of SYK-targeted therapies.
1. Introduction
The fourth revised edition of the World Health Organization (WHO) Classification of Tumours of Hematopoietic and Lymphoid Tissues defined a subgroup of entities as “Myeloid/lymphoid neoplasms with eosinophilia and rearrangement of PDGFRA, PDGFRB or FGFR1, or with PCM1-JAK2” [1]. The recognition of those entities as a separate group was justified by some shared morphological and clinical features, but even more importantly, by the already known sensitivity to available tyrosine kinase inhibitors [1]. This guiding principle of acknowledging pathological and genetic distinction along with clinical significance remained in the most recent fifth edition of the WHO classification [2]. The category was renamed to Myeloid/lymphoid neoplasms with eosinophilia and tyrosine kinase gene fusions (MLN-TK) and included entities with more recurrent genetic features such as PDGFRA rearrangement, PDGFRB rearrangement, FGFR1 rearrangement, JAK2 rearrangement, FLT3 rearrangement, ETV6::ABL1 fusion, ETV6::FGFR2; ETV6::LYN; ETV6::NTRK3; RANBP2::ALK; BCR::RET; and FGFR1OP::RET [2]. In concordance with this development of the WHO classification was also the International Consensus Classification (ICC), which used the same name for this category but included less genetic aberrations [3]. Provided the rapid advancements and wider usage of molecular genetic profiling in routine clinical practice, one can expect that in the subsequent revisions of those classifications more entities with recurrent rearrangements of tyrosine kinase genes will fall under the same category. One prominent example of such entities would be myeloid/lymphoid neoplasms with SYK gene rearrangements.
In this brief report we aimed to review the already reported cases with SYK gene rearrangements, including ETV6::SYK and ITK::SYK or other fusion partners of SYK, driving myeloid or lymphoid malignancies and to discuss the diagnostic and therapeutic implications of their wider recognition.
2. Spleen Tyrosine Kinase (SYK)
The spleen tyrosine kinase (SYK) is one of the non-receptor human tyrosine kinases which has considerable role in the hematopoietic tissues. It is a 72kDa protein, containing two SRC homology protein-binding domains (SH-2) and a kinase domain. The gene for the SYK is located on the long arm of chromosome nine (9: 90.8–90.9) and is described to interact with numerous key players, particularly along the immunoreceptors pathways. Upstream the SYK SH-2 domains would dock in the immunoreceptor tyrosine-based activation motifs (ITAMs) and downstream it would transduce the signal to numerous intermediaries including VAV, PI3K, PLC, SLP76, SLP65 involved in cell proliferation, cell differentiation, cytoskeletal changes, cell survival, and cytokine production [4,5,6]. While the role of SYK is well known in the hematopoietic tissue development and in the development of the adaptive immune response, there is also emerging evidence that it has significant levels of expression in non-hematopoietic tissues as well, such as the gastrointestinal tract and endocrine organs [7]. In the normal hematopoietic lineages, it is almost ubiquitously expressed, with the exception of CD4+ and CD8+ T cells, in which SYK expression is the lowest (Figure 1). As with other tyrosine kinases in a native state, SYK is autoinhibited, as in the absence of bonding to phosphorylated ITAMs of the SH-2 domains, two interdomains A and B (located between the two SH-2 and between the second SH-2 and the tyrosine kinase domain, respectively) would bind to the kinase domain and convert it to inactive state [4]. The role of constitutional hyperactivity of SYK has been well established, particularly in hematopoietic malignancies of mature B-cell origin [8,9], but there is growing body of evidence suggesting SYK may have a role in T-cell malignancies [10,11] and even in myeloid malignancies [12].
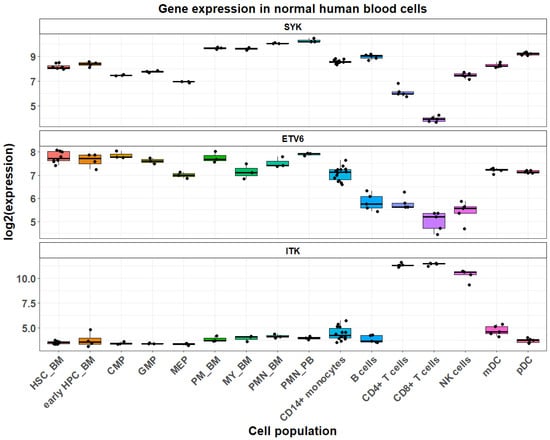
Figure 1.
Comparison of expression of the human SYK, ETV6, and ITK genes using HemaExplorer data from sorted blood cells available through the BloodSpot 3.0 server [13] (https://www.fobinf.com/) (last accessed 30 April 2025). Abbreviations: “HSC-BM”—bone marrow hematopoietic stem cells; “early HPC_BM”—early bone marrow hematopoietic progenitors; “CMP”—common myeloid progenitors; “GMP”—granulocyte monocyte progenitors; “MEP”—megakaryocyte-erythroid progenitors; “PM_BM”—bone marrow promyelocytes; “MY_BM”—bone marrow myelocytes; “PMN_BM”—bone marrow polymorphonuclear cells; “PMN_PB”—peripheral blood polymorphonuclear cells.
Translocations activating constitutively tyrosine kinases driving hematological malignancies are a well-known pathogenesis mechanism in leukemogenesis and lymphomagenesis. Several cases have been described so far, in which a fusion kinase, ETV6::SYK, demonstrates constitutive activation and evolution of clonal myeloid malignancies. Another fusion protein ITK::SYK seems to be associated with peripheral T-cell lymphoma.
3. ETV6::SYK in Myeloid Neoplasms
ETV6, previously known as TEL, is a transcription factor of the ETS (E-Twenty-Six) superfamily, that is encoded in the short arm of chromosome 12 (12p13.1). The gene spans along 250 kb and comprises eight exons and three functional domains—pointed N-terminal domain, also known as helix–loop–helix domain (PNT or HLH), internal or linker domain, and ETS domain [14,15]. ETV6 region is well known for clustering of numerous recombination events among which entities with recurrent genomic aberrations (Table 1), recognized by WHO 2022 classification, such as myeloid/lymphoid neoplasms with PDGFRA rearrangement, myeloid/lymphoid neoplasms with PDGFRB rearrangement, Myeloid/lymphoid neoplasm with ETV6::ABL1 fusion, and B-lymphoblastic leukemia/lymphoma with ETV6::RUNX1 fusion [2,16]. There appears to be an apparent clustering of the breakpoints for the specific partners, with intron 5 frequently cited as a hotspot for the most frequent translocation of the ETV6::RUNX1 fusion. Both cryptic signal V(D)J signal heptamers and also purine/pyrimidine tract repeats have been described as flanking the breakpoint cluster region of the ETV6 gene’s intron 5 [15,17]. The first described case of ETV6::SYK in myeloid malignancy to our knowledge also involves a breakpoint of ETV6 in intron 5. Apparently as with other tyrosine kinase fusion partners of ETV6, the fusion which eliminates the upstream binding sites and regulators of the tyrosine kinase, leaves a constitutionally activate tyrosine kinase, which was demonstrated with the first described case of ETV6::SYK [12]. This would drive SYK overexpression in the entire hematopoietic and progenitor cells pool and in the myeloid lineage but not in mature T cells, in which ETV6 has significantly lower expression (Figure 1).

Table 1.
Summary of the available clinical and biological characteristics of the published cases of ETV6::SYK-positive myeloid and lymphoid malignancies. Abbreviations: “CR”—complete remission; “allo-HSCT”—allogeneic hematopoietic stem cell transplant; “BSA”—body surface area; “BD”—bi in die (twice daily).
The cases described above share many features in common, primarily eosinophilia and dysplasia, particularly for the megakaryocytic line, and skin infiltrates with non-Langerhans histiocytosis type of cell population and mostly chronic course. In vitro studies are also supportive of the significance of the ETV6::SYK for the leukemogenesis demonstrating IL-3-independent proliferation for transformed cells; mouse models with the fusion protein rapidly developed myeloid dysplasia and marrow fibrosis, with the condition being fatal for mice [12,25,26]. To the best of our knowledge, the ETV6::SYK has been reported only for a single time in a non-myeloid malignancy, namely Philadelphia chromosome-negative B-ALL, which may suggest that the truncal gene fusion between ETV6 and SYK occurs before the commitment of progenitor cells to myeloid lineage [24].
A number of ex vivo and in vitro studies explored the signalling pathways affected by ETV6::SYK fusion kinase. Kanie et al. demonstrated that ETV6::SYK overexpression in BaF3 cells leads to IL-3 independence and overactivation of three key signalling pathways as follows: PI3-K/Akt; MAPK; and JAK-STAT pathways [10]. Notably overactivation of STAT5 was independent of upstream JAK2 signalling [10]. These observations were confirmed in vivo in xenotransplant mouse models [25,26]. In both models, ETV6::SYK induced severe myeloproliferative neoplasm, which was more aggressive than the one induced by ITK::SYK [26]. More recently, using primary human bone marrow cell harboring ETV6-SYK translocation, Lierman et al. clearly demonstrated overactivation [20] of PI3-K/Akt, MAPK, and JAK-STAT pathways, which were obviously dependent on SYK overexpression, as two SYK inhibitors (fostamatinib and entospletinib) were able to reverse the activation of the three pathways.
All these molecular signalling studies provided the mechanistic rationale for targeting the truncal ETV6::SYK oncogenic fusion with SYK inhibitors being currently marketed or under clinical development. To date only one patient has been reported being treated with fostamatinib [23]. Patient experienced clinically meaningful partial response, which, however, was relatively short-lived with a duration of approximately 18 months. Resequencing of the SYK gene did not reveal any additional point mutations. This observation raises the question of possible resistance to SYK inhibitors in such patients. Indeed, we surveyed the latest COSMIC database release in order to gain insight into whether fostamatinib and entospletinib binding sites are known to be frequently mutated in cancers. Previous crystallization studies showed that SYK amino acid residues that are important for its interaction with fostamatinib include L377, E449, A451, G454, P455, K458 and D512 [27,28]. Similarly, the important SYK residues for interaction with entospletinib included A451 and D512 [29].
As shown in Figure 2, mutations that may affect fostamatinib or entospletinib binding to SYK domain are very rarely reported in human cancers. Therefore, a more likely mechanism for acquired resistance to those inhibitors, when used to target ETV6::SYK, might be off-target mechanisms associated with alternative or downstream signalling pathways activation. This hypothesis was recently supported by the observation that resistance to SYK inhibition in acute myeloid leukemia (AML) was associated with re-activation of the RAS-MAPK-ERK pathway as the dominant mechanism of acquired resistance [30]. Notably, combination of entospletinib with a MEK inhibitor could overcome resistance to SYK inhibition [30]. Such an approach might also be feasible in patients with ETV6::SYK translocation either as up-front therapy or at the time of acquired resistance. In support of this we have previously demonstrated partial response to single agent therapy with a MEK inhibitor, cobimetinib, in a Bulgarian patient with ETV6::SYK-driven disease, who was not able to gain access to any SYK TKI [22].
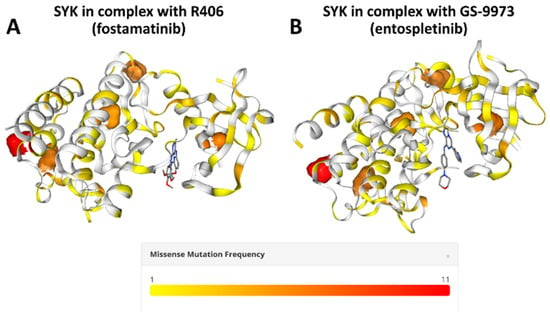
Figure 2.
Three-dimensional structures of SYK in complex with known inhibitors with annotation of the most frequently mutated amino acid residues in human cancers according to the COSMIC database. (A) SYK in complex with R406 (fostamatinib). Structure was from Protein Database with accession number 3FQS [27]. (B) SYK in complex with GS-9973 (entospletinib). Structure was from Protein Database with accession number G-9973 [29]. Both structures show that SYK residues important for inhibition by either fostamatinib or entospletinib are very rarely mutated. Pictures were generated using COSMIC-3D [31] portal (https://cancer.sanger.ac.uk/cosmic3d/) (last accessed on 3 May 2025).
4. ITK::SYK in Lymphoid Neoplasms
The interleukin-2 inducible T-cell kinase (ITK) is a tyrosine kinase with considerable significance in the downstream conducting of the TCR signal. It is very significant in the signal transduction in T-cells and has a well-established role in allergy and hypersensitivity, autoimmunity, infection, and T-cell malignancies. Its upstream partners involve PIP3 and LAT/SLP76 complex, downstream the integral kinase activates the ERK/MAPK pathways and the NfkB pathway [32]. The gene for ITK is located on the long arm of chromosome 5, and the encoded kinase has one aminoterminal pleckstrin homology (PH) domain, a TEC homology (TH) domain, two Src homology (SH) domains, and a kinase domain for TCR downstream signalling. The fusion merges the PH and TEC domains of ITK to the kinase domain of SYK. This induces constitutively active state in the SYK and produces downstream signals, mimicking TCR signal which induces oncogenesis in T cells; this has been demonstrated in mice, and human patient cases have also been reported [11,33].
The first report to our knowledge described five cases of peripheral T-cell lymphoma (PTCL, unspecified), harbouring the ITK::SYK fusion t(5;9)(q33;q22) in 30 patients tested. The same team did not find ITK::SYK among nine cases of AITL and among seven cases of ALK-negative anaplastic large cell lymphoma [11]. A second report revealed four positive patients out of twenty-two tested, again demonstrating peripheral T-cell pattern with a follicular growth and immunophenotype suggestive of origin of follicular T-helper cells [34]. It is important to note that two of the patients are reported in both studies; therefore, the total number of unique ITK::SYK cases with nodal follicular helper cell lymphoma, follicular type, would amount to a total of seven. Despite the small number of published ITK::SYK-positive cases, this fusion has already been recognized by the WHO fifth edition as relevant to the pathogenesis and is recommended for testing (https://tumourclassification.iarc.who.int/welcome/) (last accessed 9 May 2025). It is not surprising that in those cases, the phenotype is of mature T-cell lineage, as ITK expression is the highest in mature CD4+ and CD8+ T cells and in NK cells, compared to other normal human blood cells (Figure 1) [35]. Interestingly, in vivo conditional expression of ITK::SYK in mouse CD4+ T cells leads to PTCL-like lymphoproliferative neoplasm and is associated with overactivation of AKT, ERK, and p38 MAPK pathways (the ERK/MAPK pathway being associated with the normal ITK pathway activation) [33]. Another ITK::SYK transgenic mouse model also suggested that while SYK tyrosine kinase activity of the fusion protein was indispensable for malignant T-cell transformation, membrane localization of the fusion protein might not essential to T cell transformation [36]. This latter observation contrasted to initial experiments with 3T3 fibroblasts [37]. Collectively, one can expect that ITK portion of the ITK-SYK fusion protein contributes to malignant T-cell phenotype albeit without directly affecting ITK-SYK activity. This suggests that patients with ITK::SYK-positive T-cell lymphomas may benefit from targeted therapies, as demonstrated in the individual cases ETV6::SYK-positive neoplasms discussed above. The common features of ETV6::SYK-driven myeloid malignancies and ITK::SYK-positive PTCLs are further supported by a single observation of a variant ITK::SYK fusion in a patient with myeloproliferative neoplasm [23].
5. CTLC::SYK in Juvenile Xanthogranuloma
A recent study identified recurrent CTLC::SYK fusion in 6 out of 16 studied children with juvenile xanthogranuloma (JXG) [38]. Interestingly, the malignant population of these dendritic cell neoplasms stained positive for phospho-SYK, along with cyclin D1 and phospho-S6, which is a marker of mammalian target of rapamycin (mTOR) activation [38]. Moreover, phospho-ERK expression was observed in both mononucleated and multinucleated cells in two cases, whereas rare phospho-ERK+ cells were seen in the other four cases. All cases were stained negative for phospho-Akt [38]. The tumours were surgically respectable or showed spontaneous regression. However, the signs of activation of MAPK and mTOR pathways suggested molecular similarities with ETV6::SYK-positive cases, as discussed above [38].
6. Other SYK Fusions Reported in Myeloid/Lymphoid Malignancies
To our knowledge, a single case has been presented so far with a PML::SYK t(9;15) fusion [39]. Interestingly, the case, involving a 56-year-old female, bears significant similarities in its clinical course to the ETV6::SYK cases described above. The woman was diagnosed with unclassified myeloproliferative disorder on the basis of a hypercellular bone marrow, with myeloid hyperplasia, megakaryocytic dysplasia, blasts of 6%, and upon relapse after allogeneic-HSCT, the patient manifested with AML with fibrosis and skin involvement [39]. Furthermore, PML::SYK was shown to lead to PI3-K/Akt, MAPK, and JAK-STAT pathways [39]. The study suggested alternative treatment approach as the patient achieved durable remission on continuous treatment with first-in-class epichaperome inhibitor PU-H71 (zelavespib) [39].
7. Discussions and Future Directions
The accumulating case reports and case series of myeloid neoplasms with translocations, involving SYK and the ITK::SYK-positive PTCLs, demonstrating not only the correlation of this lesion to particular diagnostic categories, but also demonstrating significant similarities in the clinical course, provide justification for further larger studies, including retrospective ones, to establish the true incidence of these aberrations. These could potentially confirm the distinct biology, clinical picture, course, and prognosis of these malignancies, and justify their definition as separate diagnostic entities. We expect that in subsequent editions of WHO and ICC, ETV6::SYK fusions will be specifically included in the list of aberrations defining the MLN-TK category. In our view ITK::SYK fusion in PTCLs would remain within the pathologically defined categories of mature T-cell lymphomas, even though they may benefit from targeted therapy approaches.
SYK-fusion proteins with preserved kinase domain are also actionable mutations, as there are readily available approved therapies targeting SYK (e.g., fostamatinib targets the ATP-binding site, which is within the kinase domain and is already authorized for chronic refractory ITP). Cerdulatinib is another TKI with SYK, JAK1, JAK3, and TYK2 specificity designated by FDA as an orphan drug for PTCL, which is still under clinical development [40]. Because of its dual targeting of SYK and JAK/STAT pathways it might be particularly active for PTCL cases with ITK::SYK fusion. Furthermore, considering that the ITK::SYK pathway is involving the PI3K in the upward part of the cascade, theoretical merit for PI3K inhibitors may also be considered. While targeted treatment has not been used in the majority of isolated cases reported above, with expanded active diagnostic search, single-patient clinical trials exploring the potential of SYK-inhibitors or programmes for their compassionate use in such rare cases may become a more attractive concept.
Author Contributions
V.S. proposed research, generated figures and drafted the manuscript. S.L. collected data, prepared the table, and drafted the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Velizar Shivarov is supported by the Bulgarian National Science Fund (BNSF) grants KP-06-H 41/2 and KP-06-H 83/3.
Institutional Review Board Statement
This work is considered exempt from IRB review as it is a review paper.
Informed Consent Statement
Not applicable as this study uses free publicly available data.
Data Availability Statement
All sources of data used to generate the figures in the manuscript are referenced accordingly in the manuscript text.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef] [PubMed]
- Khoury, J.D.; Solary, E.; Abla, O.; Akkari, Y.; Alaggio, R.; Apperley, J.F.; Bejar, R.; Berti, E.; Busque, L.; Chan, J.K. The 5th edition of the World Health Organization classification of haematolymphoid tumours: Myeloid and histiocytic/dendritic neoplasms. Leukemia 2022, 36, 1703–1719. [Google Scholar] [CrossRef] [PubMed]
- Arber, D.A.; Orazi, A.; Hasserjian, R.P.; Borowitz, M.J.; Calvo, K.R.; Kvasnicka, H.-M.; Wang, S.A.; Bagg, A.; Barbui, T.; Branford, S.; et al. International Consensus Classification of Myeloid Neoplasms and Acute Leukemias: Integrating morphologic, clinical, and genomic data. Blood 2022, 140, 1200–1228. [Google Scholar] [CrossRef] [PubMed]
- Mócsai, A.; Ruland, J.; Tybulewicz, V.L. The SYK tyrosine kinase: A crucial player in diverse biological functions. Nat. Rev. Immunol. 2010, 10, 387–402. [Google Scholar] [CrossRef]
- Hobbs, H.T.; Shah, N.H.; Badroos, J.M.; Gee, C.L.; Marqusee, S.; Kuriyan, J. Differences in the dynamics of the tandem-SH2 modules of the Syk and ZAP-70 tyrosine kinases. Protein Sci. 2021, 30, 2373–2384. [Google Scholar] [CrossRef]
- Bradshaw, J.M. The Src, Syk, and Tec family kinases: Distinct types of molecular switches. Cell. Signal. 2010, 22, 1175–1184. [Google Scholar] [CrossRef]
- Yanagi, S.; Inatome, R.; Takano, T.; Yamamura, H. Syk expression and novel function in a wide variety of tissues. Biochem. Biophys. Res. Commun. 2001, 288, 495–498. [Google Scholar] [CrossRef]
- Buchner, M.; Fuchs, S.; Prinz, G.; Pfeifer, D.; Bartholomé, K.; Burger, M.; Chevalier, N.; Vallat, L.; Timmer, J.; Gribben, J.G. Spleen tyrosine kinase is overexpressed and represents a potential therapeutic target in chronic lymphocytic leukemia. Cancer Res. 2009, 69, 5424–5432. [Google Scholar] [CrossRef]
- Buchner, M.; Baer, C.; Prinz, G.; Dierks, C.; Burger, M.; Zenz, T.; Stilgenbauer, S.; Jumaa, H.; Veelken, H.; Zirlik, K. Spleen tyrosine kinase inhibition prevents chemokine-and integrin-mediated stromal protective effects in chronic lymphocytic leukemia. Blood 2010, 115, 4497–4506. [Google Scholar] [CrossRef]
- Feldman, A.; Sun, D.; Law, M.; Novak, A.; Attygalle, A.; Thorland, E.; Fink, S.; Vrana, J.; Caron, B.; Morice, W. Overexpression of Syk tyrosine kinase in peripheral T-cell lymphomas. Leukemia 2008, 22, 1139–1143. [Google Scholar] [CrossRef]
- Streubel, B.; Vinatzer, U.; Willheim, M.; Raderer, M.; Chott, A. Novel t (5; 9)(q33; q22) fuses ITK to SYK in unspecified peripheral T-cell lymphoma. Leukemia 2006, 20, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Kuno, Y.; Abe, A.; Emi, N.; Iida, M.; Yokozawa, T.; Towatari, M.; Tanimoto, M.; Saito, H. Constitutive kinase activation of the TEL-Syk fusion gene in myelodysplastic syndrome with t (9; 12)(q22; p12). Blood 2001, 97, 1050–1055. [Google Scholar] [CrossRef]
- Gíslason, M.H.; Demircan, G.S.; Prachar, M.; Furtwängler, B.; Schwaller, J.; Schoof, E.M.; Porse, B.T.; Rapin, N.; Bagger, F.O. BloodSpot 3.0: A database of gene and protein expression data in normal and malignant haematopoiesis. Nucleic Acids Res. 2024, 52, D1138–D1142. [Google Scholar]
- Bohlander, S.K. ETV6: A versatile player in leukemogenesis. Semin. Cancer Biol. 2005, 15, 162–174. [Google Scholar] [CrossRef] [PubMed]
- De Braekeleer, E.; Douet-Guilbert, N.; Morel, F.; Le Bris, M.-J.; Basinko, A.; De Braekeleer, M. ETV6 fusion genes in hematological malignancies: A review. Leuk. Res. 2012, 36, 945–961. [Google Scholar] [CrossRef]
- Alaggio, R.; Amador, C.; Anagnostopoulos, I.; Attygalle, A.D.; Araujo, I.B.d.O.; Berti, E.; Bhagat, G.; Borges, A.M.; Boyer, D.; Calaminici, M. The 5th edition of the World Health Organization classification of haematolymphoid tumours: Lymphoid neoplasms. Leukemia 2022, 36, 1720–1748. [Google Scholar] [CrossRef]
- Thandla, S.P.; Ploski, J.E.; Raza-Egilmez, S.Z.; Chhalliyil, P.P.; Block, A.W.; De Jong, P.J.; Aplan, P.D. ETV6-AML1 translocation breakpoints cluster near a purine/pyrimidine repeat region in the ETV6 gene. Blood J. Am. Soc. Hematol. 1999, 93, 293–299. [Google Scholar]
- Kuno, Y.; Abe, A.; Emi, N.; Iida, M.; Yamamori, T.; Tanimoto, M.; Saito, H. An atypical myelodysplastic syndrome with t (9; 12)(q22; p12) and TEL gene rearrangement. Br. J. Haematol. 1999, 106, 570–571. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodi, M.; Swami, V.K.; Besa, E.C.; Punnett, H.H. A second case of myelodysplastic syndrome with t (9; 12)(q22; p12). Cancer Genet. Cytogenet. 2005, 157, 187–188. [Google Scholar] [CrossRef]
- Lierman, E.; Smits, S.; Debackere, K.; André, M.; Michaux, L.; Vandenberghe, P. t (9; 12)(q22; p13) ETV6:: SYK: A new recurrent cytogenetic aberration and tyrosine kinase gene fusion in myeloid or lymphoid neoplasms associated with eosinophilia. Br. J. Haematol. 2023, 200, 665–668. [Google Scholar] [CrossRef]
- Debois, D.; Marot, L.; Andre, M.; Dachelet, C. Thalidomide as an effective treatment for adult multiple xanthogranuloma. JAAD Case Rep. 2018, 4, 896–898. [Google Scholar] [CrossRef] [PubMed]
- Manuelyan, K.; Momcheva, I.; Angelova, S.; Nikolov, K.; Shivarov, V. Recurrent ETV6:: SYK rearrangement in myeloid malignancies confers partial susceptibility to MEK inhibition. Br. J. Haematol. 2024, 205, 382–386. [Google Scholar] [CrossRef]
- Risch, Z.; Kaffenberger, B.H.; Chung, C.G.; Samorodnitsky, E.; Hoskins, E.L.; Dao, T.; Smith, A.; Wall, S.A.; Brammer, J.; Reeser, J.W. Myeloid neoplasm with histiocytosis and spleen tyrosine kinase fusion responds to fostamatinib. Haematologica 2024, 109, 3816. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.-H.; Gao, L.; Jing, Y.; Xu, Y.-Y.; Ding, Y.; Wang, N.; Wang, W.; Li, M.-Y.; Han, X.-P.; Sun, J.-z. Detection of ETV6 gene rearrangements in adult acute lymphoblastic leukemia. Ann. Hematol. 2012, 91, 1235–1243. [Google Scholar] [CrossRef]
- Graham, M.T.; Abram, C.L.; Hu, Y.; Lowell, C.A. Expression of the TEL-Syk fusion protein in hematopoietic stem cells leads to rapidly fatal myelofibrosis in mice. PLoS ONE 2013, 8, e77542. [Google Scholar] [CrossRef] [PubMed]
- Sprissler, C.; Belenki, D.; Maurer, H.; Aumann, K.; Pfeifer, D.; Klein, C.; Müller, T.; Kissel, S.; Hülsdünker, J.; Alexandrovski, J. Depletion of STAT5 blocks TEL–SYK-induced APMF-type leukemia with myelofibrosis and myelodysplasia in mice. Blood Cancer J. 2014, 4, e240. [Google Scholar] [CrossRef]
- Villaseñor, A.G.; Kondru, R.; Ho, H.; Wang, S.; Papp, E.; Shaw, D.; Barnett, J.W.; Browner, M.F.; Kuglstatter, A. Structural insights for design of potent spleen tyrosine kinase inhibitors from crystallographic analysis of three inhibitor complexes. Chem. Biol. Drug Des. 2009, 73, 466–470. [Google Scholar] [CrossRef]
- Braselmann, S.; Taylor, V.; Zhao, H.; Wang, S.; Sylvain, C.; Baluom, M.; Qu, K.; Herlaar, E.; Lau, A.; Young, C. R406, an orally available spleen tyrosine kinase inhibitor blocks fc receptor signaling and reduces immune complex-mediated inflammation. J. Pharmacol. Exp. Ther. 2006, 319, 998–1008. [Google Scholar] [CrossRef]
- Currie, K.S.; Kropf, J.E.; Lee, T.; Blomgren, P.; Xu, J.; Zhao, Z.; Gallion, S.; Whitney, J.A.; Maclin, D.; Lansdon, E.B. Discovery of GS-9973, a selective and orally efficacious inhibitor of spleen tyrosine kinase. J. Med. Chem. 2014, 57, 3856–3873. [Google Scholar] [CrossRef]
- Cremer, A.; Ellegast, J.M.; Alexe, G.; Frank, E.S.; Ross, L.; Chu, S.H.; Pikman, Y.; Robichaud, A.; Goodale, A.; Häupl, B. Resistance mechanisms to SYK inhibition in acute myeloid leukemia. Cancer Discov. 2020, 10, 214–231. [Google Scholar] [CrossRef]
- Jubb, H.C.; Saini, H.K.; Verdonk, M.L.; Forbes, S.A. COSMIC-3D provides structural perspectives on cancer genetics for drug discovery. Nat. Genet. 2018, 50, 1200–1202. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Kannan, A.K.; August, A. Structure and function of Tec family kinase Itk. Biomol. Concepts 2011, 2, 223–232. [Google Scholar] [CrossRef][Green Version]
- Pechloff, K.; Holch, J.; Ferch, U.; Schweneker, M.; Brunner, K.; Kremer, M.; Sparwasser, T.; Quintanilla-Martinez, L.; Zimber-Strobl, U.; Streubel, B. The fusion kinase ITK-SYK mimics a T cell receptor signal and drives oncogenesis in conditional mouse models of peripheral T cell lymphoma. J. Exp. Med. 2010, 207, 1031–1044. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Moreau, A.; Dupuis, J.; Streubel, B.; Petit, B.; Le Gouill, S.; Martin-Garcia, N.; Copie-Bergman, C.; Gaillard, F.; Qubaja, M. Peripheral T-cell lymphomas with a follicular growth pattern are derived from follicular helper T cells (TFH) and may show overlapping features with angioimmunoblastic T-cell lymphomas. Am. J. Surg. Pathol. 2009, 33, 682–690. [Google Scholar] [CrossRef]
- Berg, L.J.; Finkelstein, L.D.; Lucas, J.A.; Schwartzberg, P.L. Tec family kinases in T lymphocyte development and function. Annu. Rev. Immunol. 2005, 23, 549–600. [Google Scholar] [CrossRef]
- Dierks, C.; Adrian, F.; Fisch, P.; Ma, H.; Maurer, H.; Herchenbach, D.; Forster, C.U.; Sprissler, C.; Liu, G.; Rottmann, S. The ITK-SYK fusion oncogene induces a T-cell lymphoproliferative disease in mice mimicking human disease. Cancer Res. 2010, 70, 6193–6204. [Google Scholar] [CrossRef] [PubMed]
- Rigby, S.; Huang, Y.; Streubel, B.; Chott, A.; Du, M.-Q.; Turner, S.D.; Bacon, C.M. The lymphoma-associated fusion tyrosine kinase ITK-SYK requires pleckstrin homology domain-mediated membrane localization for activation and cellular transformation. J. Biol. Chem. 2009, 284, 26871–26881. [Google Scholar] [CrossRef]
- Kemps, P.G.; Baelde, H.J.; Vorderman, R.H.; Stelloo, E.; Swennenhuis, J.F.; Szuhai, K.; Lamers, M.H.; Kenkhuis, B.; Al-Hussaini, M.; Briaire-de Bruijn, I.H. Recurrent CLTC:: SYK fusions and CSF1R mutations in juvenile xanthogranuloma of soft tissue. Blood 2024, 144, 2439–2455. [Google Scholar] [CrossRef]
- Sugita, M.; Wilkes, D.C.; Bareja, R.; Eng, K.W.; Nataraj, S.; Jimenez-Flores, R.A.; Yan, L.; De Leon, J.P.; Croyle, J.A.; Kaner, J. Targeting the epichaperome as an effective precision medicine approach in a novel PML-SYK fusion acute myeloid leukemia. NPJ Precis. Oncol. 2021, 5, 44. [Google Scholar] [CrossRef]
- Horwitz, S.M.; Feldman, T.A.; Ye, J.C.; Khodadoust, M.S.; Munoz, J.; Hamlin, P.A.; Kim, Y.H.; Wilcox, R.A.; Patel, M.R.; Coffey, G. Results from an open-label phase 2a study of cerdulatinib, a dual spleen tyrosine kinase/janus kinase inhibitor, in relapsed/refractory peripheral T-cell lymphoma. Leuk. Lymphoma 2025, 66, 1100–1110. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).