
Increasing the High Throughput of a Luminescence-Based Serum Bactericidal Assay (L-SBA)




Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Reagents
2.2. Serum Samples
2.3. Luminescent-Based SBA (L-SBA) in 96- and 384-Well Plates
2.4. Calculations
3. Results
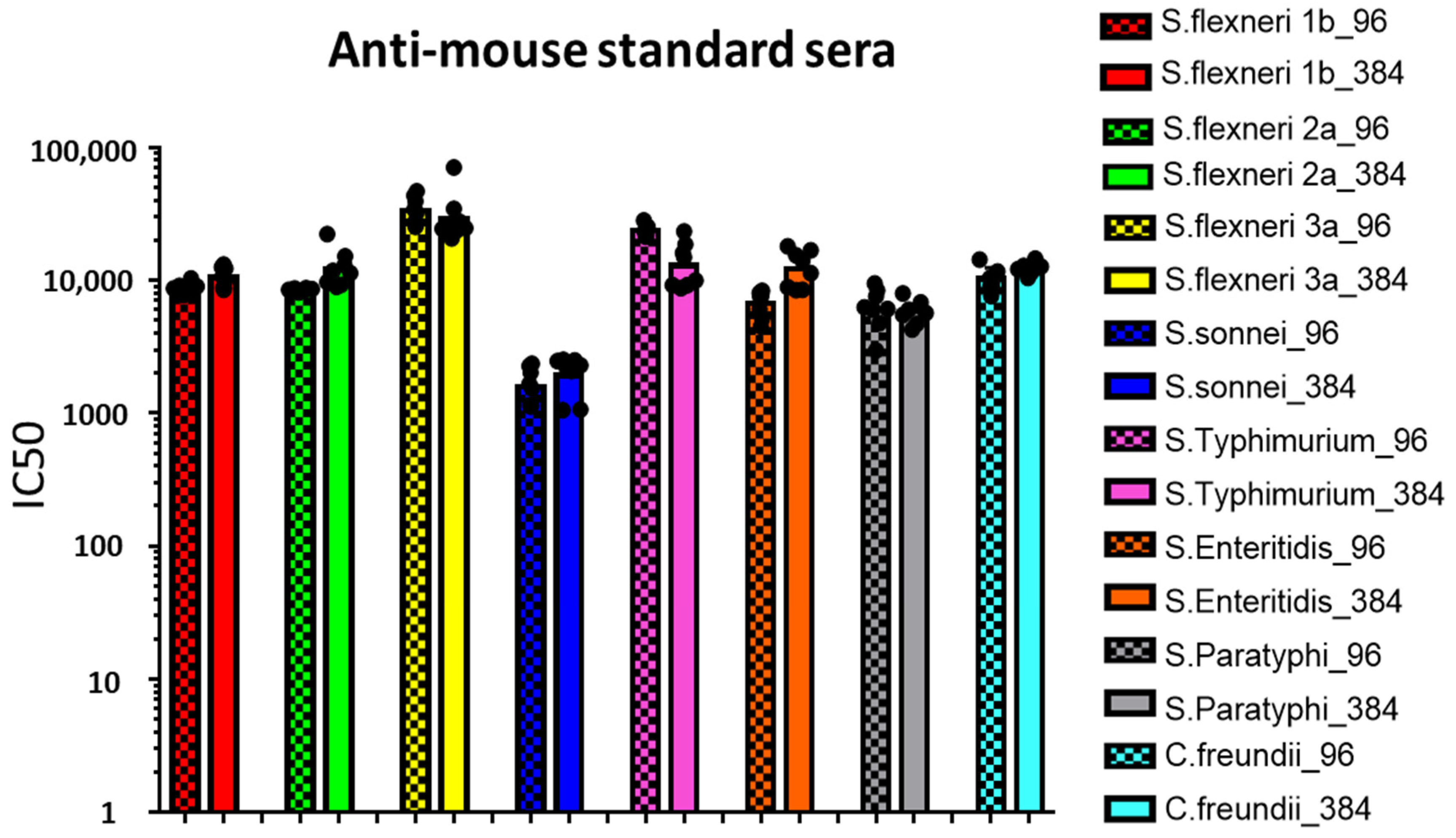
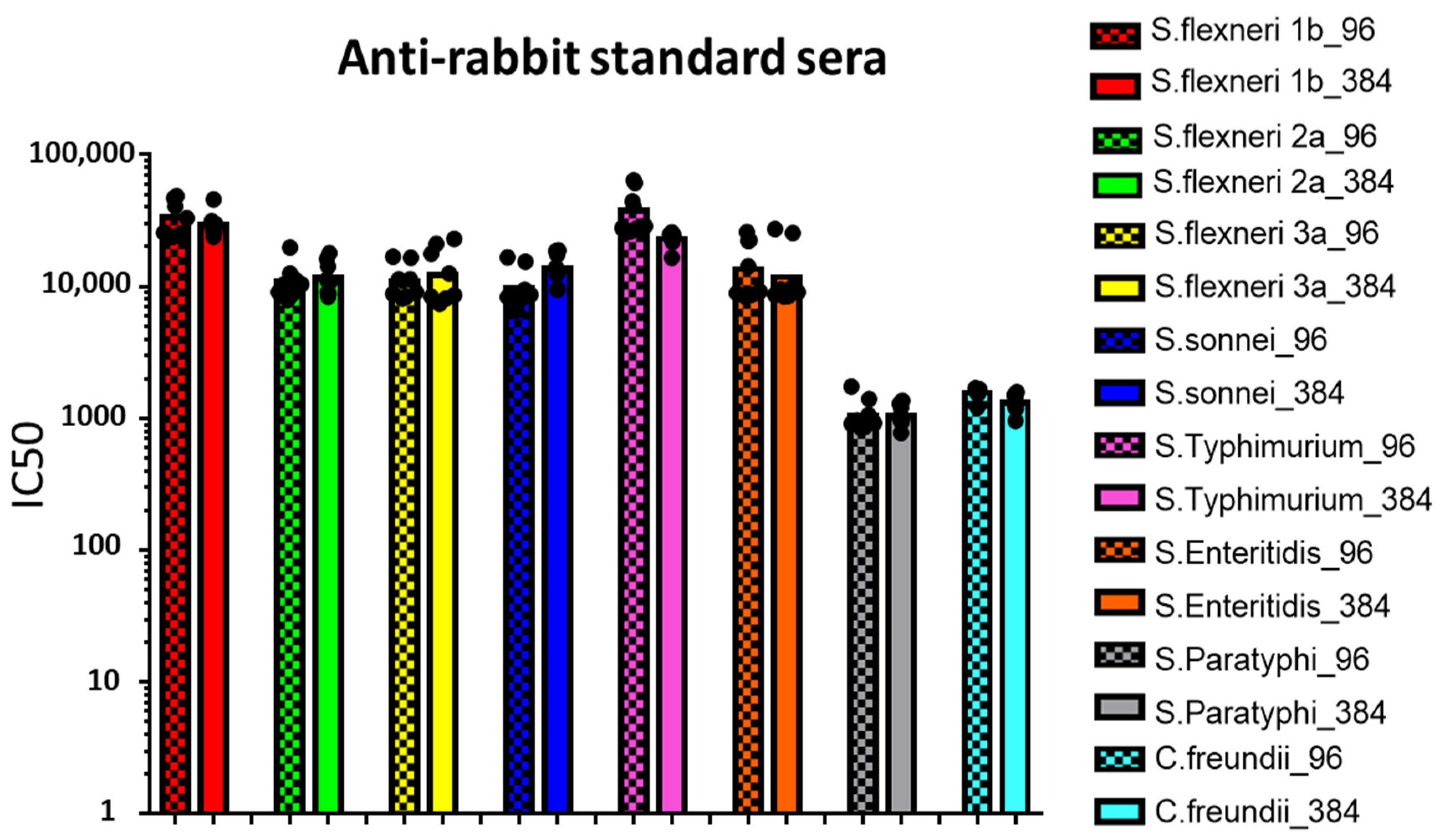
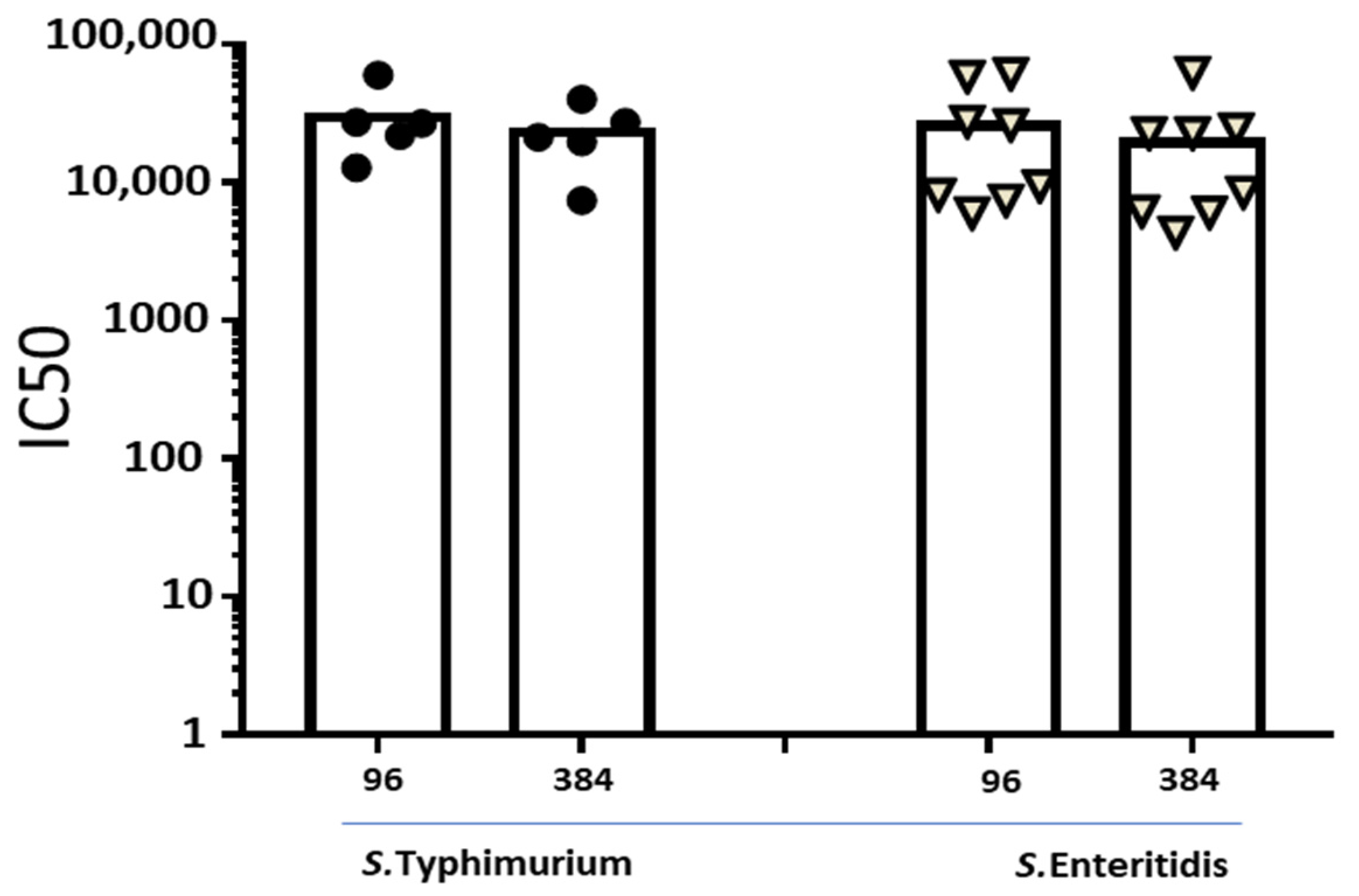
3.1. Moving from 96- to 384-Well Plates
3.2. Comparison between L-SBA in 96- and 384-Well Plates
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Son, M.S.; Taylor, R.K. Vibriocidal assays to determine the antibody titer of patient sera samples. Curr. Protoc. Microbiol. 2011, 23, 6A.3.1–6A.3.9. [Google Scholar] [CrossRef]
- Borrow, R.; Carlone, G.M.; Rosenstein, N.; Blake, M.; Feavers, I.; Martin, D.; Zollinger, W.; Robbins, J.; Aaberge, I.; Granoff, D.M.; et al. Neisseria meningitidis group B correlates of protection and assay standardization—International meeting report Emory University, Atlanta, Georgia, United States, 16–17 March 2005. Vaccine 2006, 24, 5093–5107. [Google Scholar] [CrossRef]
- Giersing, B.K.; Porter, C.K.; Kotloff, K.; Neels, P.; Cravioto, A.; MacLennan, C.A. How can controlled human infection models accelerate clinical development and policy pathways for vaccines against Shigella? Vaccine 2019, 37, 4778–4783. [Google Scholar] [CrossRef] [PubMed]
- Boyd, M.A.; Tennant, S.M.; Saague, V.A.; Simon, R.; Muhsen, K.; Ramachandran, G.; Cross, A.S.; Galen, J.E.; Pasetti, M.F.; Levine, M.M. Serum bactericidal assays to evaluate typhoidal and nontyphoidal Salmonella vaccines. Clin. Vaccine Immunol. 2014, 21, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.S.; Sahastrabuddhe, S.; Yun, C.H.; Han, S.H.; Yang, J.S. Serum bactericidal assay for the evaluation of typhoid vaccine using a semi-automated colony-counting method. Microb. Pathog. 2016, 97, 19–26. [Google Scholar] [CrossRef]
- Nahm, M.H.; Yu, J.; Weerts, H.P.; Wenzel, H.; Tamilselvi, C.S.; Chandrasekaran, L.; Pasetti, M.F.; Mani, S.; Kaminski, R.W. Development, Interlaboratory Evaluations, and Application of a Simple, High-Throughput Shigella Serum Bactericidal Assay. Msphere 2018, 3, e00146-18. [Google Scholar] [CrossRef]
- Kim, H.W.; Kim, K.H.; Kim, J.; Nahm, M.H. A high throughput serum bactericidal assay for antibodies to Haemophilus influenzae type b. BMC Infect. Dis. 2016, 16, 473. [Google Scholar] [CrossRef]
- Necchi, F.; Saul, A.; Rondini, S. Development of a high-throughput method to evaluate serum bactericidal activity using bacterial ATP measurement as survival readout. PLoS ONE 2017, 12, e0172163. [Google Scholar] [CrossRef] [PubMed]
- Necchi, F.; Saul, A.; Rondini, S. Setup of luminescence-based serum bactericidal assay against Salmonella Paratyphi A. J. Immunol. Methods 2018, 461, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Rossi, O.; Molesti, E.; Saul, A.; Giannelli, C.; Micoli, F.; Necchi, F. Intra-Laboratory Evaluation of Luminescence Based High-Throughput Serum Bactericidal Assay (L-SBA) to Determine Bactericidal Activity of Human Sera against Shigella. High Throughput 2020, 9, 14. [Google Scholar] [CrossRef]
- Mancini, F.; Gasperini, G.; Rossi, O.; Aruta, M.G.; Raso, M.M.; Alfini, R.; Biagini, M.; Necchi, F.; Micoli, F. Dissecting the contribution of O-Antigen and proteins to the immunogenicity of Shigella sonnei generalized modules for membrane antigens (GMMA). Sci. Rep. 2021, 11, 906. [Google Scholar] [CrossRef]
- Palmieri, E.; Arato, V.; Oldrini, D.; Ricchetti, B.; Aruta, M.G.; Pansegrau, W.; Marchi, S.; Giusti, F.; Ferlenghi, I.; Rossi, O.; et al. Stability of Outer Membrane Vesicles-Based Vaccines, Identifying the Most Appropriate Methods to Detect Changes in Vaccine Potency. Vaccines 2021, 9, 229. [Google Scholar] [CrossRef] [PubMed]
- Richardson, N.I.; Ravenscroft, N.; Arato, V.; Oldrini, D.; Micoli, F.; Kuttel, M.M. Conformational and Immunogenicity Studies of the Shigella flexneri Serogroup 6 O-Antigen: The Effect of O-Acetylation. Vaccines 2021, 9, 432. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Rossi, O.; Conti, V.; Launay, O.; Scire, A.S.; Aruta, M.G.; Nakakana, U.N.; Marchetti, E.; Rappuoli, R.; Saul, A.; et al. Antibodies Elicited by the Shigella sonnei GMMA Vaccine in Adults Trigger Complement-Mediated Serum Bactericidal Activity: Results from a Phase 1 Dose Escalation Trial Followed by a Booster Extension. Front. Immunol. 2021, 12, 671325. [Google Scholar] [CrossRef]
- Juel, H.B.; Thomaides-Brears, H.B.; Darton, T.C.; Jones, C.; Jones, E.; Shrestha, S.; Sie, R.; Eustace, A.; Galal, U.; Kurupati, P.; et al. Salmonella Typhi Bactericidal Antibodies Reduce Disease Severity but Do Not Protect against Typhoid Fever in a Controlled Human Infection Model. Front. Immunol. 2017, 8, 1916. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.A.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdelalim, A.; Abdollahpour, I.; et al. Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980-2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1736–1788. [Google Scholar] [CrossRef]
- Khalil, I.A.; Troeger, C.; Blacker, B.F.; Rao, P.C.; Brown, A.; Atherly, D.E.; Brewer, T.G.; Engmann, C.M.; Houpt, E.R.; Kang, G.; et al. Morbidity and mortality due to shigella and enterotoxigenic Escherichia coli diarrhoea: The Global Burden of Disease Study 1990-2016. Lancet Infect. Dis. 2018, 18, 1229–1240. [Google Scholar] [CrossRef]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- de Jong, H.K.; Parry, C.M.; van der Poll, T.; Wiersinga, W.J. Host-pathogen interaction in invasive Salmonellosis. PLoS Pathog. 2012, 8, e1002933. [Google Scholar] [CrossRef]
- Connor, B.A.; Schwartz, E. Typhoid and paratyphoid fever in travellers. Lancet Infect. Dis. 2005, 5, 623–628. [Google Scholar] [CrossRef]
- von Seidlein, L.; Kim, D.R.; Ali, M.; Lee, H.; Wang, X.; Thiem, V.D.; Canh, D.G.; Chaicumpa, W.; Agtini, M.D.; Hossain, A.; et al. A multicentre study of Shigella diarrhoea in six Asian countries: Disease burden, clinical manifestations, and microbiology. PLoS Med. 2006, 3, e353. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.M.; Kotloff, K.L.; Barry, E.M.; Pasetti, M.F.; Sztein, M.B. Clinical trials of Shigella vaccines: Two steps forward and one step back on a long, hard road. Nat. Rev. Microbiol. 2007, 5, 540–553. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, C.A.; Martin, L.B.; Micoli, F. Vaccines against invasive Salmonella disease: Current status and future directions. Hum. Vaccin Immunother. 2014, 10, 1478–1493. [Google Scholar] [CrossRef]
- Micoli, F.; Rondini, S.; Alfini, R.; Lanzilao, L.; Necchi, F.; Negrea, A.; Rossi, O.; Brandt, C.; Clare, S.; Mastroeni, P.; et al. Comparative immunogenicity and efficacy of equivalent outer membrane vesicle and glycoconjugate vaccines against nontyphoidal Salmonella. Proc. Natl. Acad. Sci. USA 2018, 115, 10428–10433. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, G.; Alfini, R.; Cescutti, P.; Caboni, M.; Lanzilao, L.; Necchi, F.; Saul, A.; MacLennan, C.A.; Rondini, S.; Micoli, F. Characterization of O-antigen delivered by Generalized Modules for Membrane Antigens (GMMA) vaccine candidates against nontyphoidal Salmonella. Vaccine 2017, 35, 419–426. [Google Scholar] [CrossRef]
- Gerke, C.; Colucci, A.M.; Giannelli, C.; Sanzone, S.; Vitali, C.G.; Sollai, L.; Rossi, O.; Martin, L.B.; Auerbach, J.; Di Cioccio, V.; et al. Production of a Shigella sonnei Vaccine Based on Generalized Modules for Membrane Antigens (GMMA), 1790GAHB. PLoS ONE 2015, 10, e0134478. [Google Scholar] [CrossRef] [PubMed]
- Rondini, S.; Micoli, F.; Lanzilao, L.; Pisoni, I.; Di Cioccio, V.; Saul, A.J.; Martin, L.B. Characterization of Citrobacter sp. line 328 as a source of Vi for a Vi-CRM(197) glycoconjugate vaccine against Salmonella Typhi. J. Infect. Dev. Ctries 2012, 6, 763–773. [Google Scholar] [CrossRef]
- MacLennan, C.A.; Gondwe, E.N.; Msefula, C.L.; Kingsley, R.A.; Thomson, N.R.; White, S.A.; Goodall, M.; Pickard, D.J.; Graham, S.M.; Dougan, G.; et al. The neglected role of antibody in protection against bacteremia caused by nontyphoidal strains of Salmonella in African children. J. Clin. Investig. 2008, 118, 1553–1562. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Msefula, C.L.; Thomson, N.R.; Kariuki, S.; Holt, K.E.; Gordon, M.A.; Harris, D.; Clarke, L.; Whitehead, S.; Sangal, V.; et al. Epidemic multiple drug resistant Salmonella Typhimurium causing invasive disease in sub-Saharan Africa have a distinct genotype. Genome Res. 2009, 19, 2279–2287. [Google Scholar] [CrossRef]
- Onsare, R.S.; Micoli, F.; Lanzilao, L.; Alfini, R.; Okoro, C.K.; Muigai, A.W.; Revathi, G.; Saul, A.; Kariuki, S.; MacLennan, C.A.; et al. Relationship between antibody susceptibility and lipopolysaccharide O-antigen characteristics of invasive and gastrointestinal nontyphoidal Salmonellae isolates from Kenya. PLoS Negl. Trop. Dis. 2015, 9, e0003573. [Google Scholar] [CrossRef]
- Dobinson, H.C.; Gibani, M.M.; Jones, C.; Thomaides-Brears, H.B.; Voysey, M.; Darton, T.C.; Waddington, C.S.; Campbell, D.; Milligan, I.; Zhou, L.; et al. Evaluation of the Clinical and Microbiological Response to Salmonella Paratyphi A Infection in the First Paratyphoid Human Challenge Model. Clin. Infect. Dis. 2017, 64, 1066–1073. [Google Scholar] [CrossRef]
- Micoli, F.; Bjarnarson, S.P.; Arcuri, M.; Aradottir Pind, A.A.; Magnusdottir, G.J.; Necchi, F.; Di Benedetto, R.; Carducci, M.; Schiavo, F.; Giannelli, C.; et al. Short Vi-polysaccharide abrogates T-independent immune response and hyporesponsiveness elicited by long Vi-CRM197 conjugate vaccine. Proc. Natl. Acad. Sci. USA 2020, 117, 24443–24449. [Google Scholar] [CrossRef]
- Arcuri, M.; Di Benedetto, R.; Cunningham, A.F.; Saul, A.; MacLennan, C.A.; Micoli, F. The influence of conjugation variables on the design and immunogenicity of a glycoconjugate vaccine against Salmonella Typhi. PLoS ONE 2017, 12, e0189100. [Google Scholar] [CrossRef]
- Micoli, F.; Rondini, S.; Gavini, M.; Lanzilao, L.; Medaglini, D.; Saul, A.; Martin, L.B. O:2-CRM(197) conjugates against Salmonella Paratyphi A. PLoS ONE 2012, 7, e47039. [Google Scholar] [CrossRef]
- Gasperini, G.; Raso, M.M.; Arato, V.; Aruta, M.G.; Cescutti, P.; Necchi, F.; Micoli, F. Effect of O-Antigen Chain Length Regulation on the Immunogenicity of Shigella and Salmonella Generalized Modules for Membrane Antigens (GMMA). Int. J. Mol. Sci. 2021, 22, 1309. [Google Scholar] [CrossRef] [PubMed]
- Campbell, H.; Borrow, R.; Salisbury, D.; Miller, E. Meningococcal C conjugate vaccine: The experience in England and Wales. Vaccine 2009, 27 (Suppl. 2), B20–B29. [Google Scholar] [CrossRef]
- Gill, C.J.; Baxter, R.; Anemona, A.; Ciavarro, G.; Dull, P. Persistence of immune responses after a single dose of Novartis meningococcal serogroup A, C, W-135 and Y CRM-197 conjugate vaccine (Menveo(R)) or Menactra(R) among healthy adolescents. Hum. Vaccines 2010, 6, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Mak, P.A.; Santos, G.F.; Masterman, K.A.; Janes, J.; Wacknov, B.; Vienken, K.; Giuliani, M.; Herman, A.E.; Cooke, M.; Mbow, M.L.; et al. Development of an automated, high-throughput bactericidal assay that measures cellular respiration as a survival readout for Neisseria meningitidis. Clin. Vaccine Immunol. 2011, 18, 1252–1260. [Google Scholar] [CrossRef] [PubMed][Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species and Serovar | Strain | Characteristics | Reference(s) |
|---|---|---|---|
| Shigella flexneri serotype 1b | 140 | Clinical isolate | Public Health England (PHE) |
| Shigella flexneri serotype 2a | 142 | Clinical isolate | Public Health England (PHE) |
| Shigella flexneri serotype 3a | 144 | Clinical isolate | Public Health England (PHE) |
| Shigella sonnei | 71 | S. sonnei 53G ΔvirG::cat | [26] |
| Citrobacter freundii | NVGH328 | Clinical isolate from Novartis Master Culture | [27] |
| Salmonella enterica serovar Typhimurium | D23580 | Clinical isolate from blood culture, Malawi | [28,29] |
| Salmonella enterica serovar Enteritidis | CMCC4314 | (corresponding to ATCC4931) obtained from the Novartis Master Culture Collection (NMCC) | [30] |
| Salmonella Paratyphi A | NVGH308 | Invasive isolates, Nepal | [31] |
| L-SBA Titers (IC50) | L-SBA Titers (IC50) | |||||||
|---|---|---|---|---|---|---|---|---|
| 96-Wells-Plate SBA | 384-Wells-Plate SBA | 96-Wells-Plate SBA | 384-Wells-Plate SBA | |||||
| S. flexneri1b Strain | Mouse Standard Antiserum | Rep. 1 | 8126 | 8490 | Rabbit Standard Antiserum | Rep. 1 | 24,732 | 29,776 |
| Rep. 2 | 7738 | 12,255 | Rep. 2 | 25,699 | 31,448 | |||
| Rep. 3 | 8257 | 10,670 | Rep. 3 | 33,031 | 26,493 | |||
| Rep. 4 | 9092 | 8694 | Rep. 4 | 46,997 | 28,192 | |||
| Rep. 5 | 8697 | 9869 | Rep. 5 | 48,571 | 24,008 | |||
| Rep. 6 | 10,322 | 8784 | Rep. 6 | 40,635 | 25,847 | |||
| Rep. 7 | 8965 | 13,089 | Rep. 7 | 23,901 | 45,903 | |||
| Rep. 8 | 8533 | 12,554 | Rep. 8 | 28,454 | ||||
| GeoMean | 8687 | 10,406 | GeoMean | 33,418 | 29,456 | |||
| SE | 260 | 621 | SE | 3481 | 2258 | |||
| SE% | 3 | 6 | SE% | 10 | 8 | |||
| S. flexneri2a strain | Mouse standard antiserum | Rep. 1 | 8119 | 8905 | Rabbit Standard antiserum | Rep. 1 | 8770 | 8380 |
| Rep. 2 | 8473 | 9682 | Rep. 2 | 11,246 | 9054 | |||
| Rep. 3 | 8477 | 22,253 | Rep. 3 | 12,632 | 18,019 | |||
| Rep. 4 | 8450 | 15,172 | Rep. 4 | 19,843 | 14,242 | |||
| Rep. 5 | 8657 | 9233 | Rep. 5 | 10,420 | 8821 | |||
| Rep. 6 | 8741 | 11,761 | Rep. 6 | 10,428 | 16,238 | |||
| Rep. 7 | 8591 | 11,323 | Rep. 7 | 9104 | 10,757 | |||
| Rep. 8 | 8376 | 12,974 | Rep. 8 | 7926 | 11,323 | |||
| GeoMean | 8484 | 12,116 | GeoMean | 10,863 | 11,652 | |||
| SE | 63 | 1456 | SE | 1243 | 1206 | |||
| SE% | 1 | 12 | SE% | 11 | 10 | |||
| S. flexneri3a Strain | mouse Standard Antiserum | Rep. 1 | 46,819 | 24,862 | Rabbit standard antiserum | Rep. 1 | 8811 | 12,624 |
| Rep. 2 | 25,205 | 70,660 | Rep. 2 | 11,374 | 21,178 | |||
| Rep. 3 | 43,233 | 56,883 | Rep. 3 | 11,343 | 23,076 | |||
| Rep. 4 | 27,388 | 40,699 | Rep. 4 | 16,600 | 17,792 | |||
| Rep. 5 | 39,309 | 25,032 | Rep. 5 | 16,928 | 7450 | |||
| Rep. 6 | 34,009 | 24,001 | Rep. 6 | 9086 | 8619 | |||
| Rep. 7 | 32,125 | 24,675 | Rep. 7 | 8417 | 8149 | |||
| Rep. 8 | 26,146 | 23,684 | Rep. 8 | 8141 | 8387 | |||
| GeoMean | 33,454 | 33,063 | GeoMean | 10,897 | 12,171 | |||
| SE | 2691 | 6030 | SE | 1180 | 2110 | |||
| SE% | 8 | 18 | SE% | 11 | 17 | |||
| S. sonneiStrain | Mouse Standard Antiserum | Rep. 1 | 1658 | 2517 | Rabbit standard antiserum | Rep. 1 | 6630 | 17,512 |
| Rep. 2 | 1150 | 2290 | Rep. 2 | 8097 | 9570 | |||
| Rep. 3 | 2357 | 2081 | Rep. 3 | 8673 | 12,372 | |||
| Rep. 4 | 1529 | 2460 | Rep. 4 | 9433 | 18,587 | |||
| Rep. 5 | 1139 | 2489 | Rep. 5 | 16,669 | 18,851 | |||
| Rep. 6 | 1121 | 1068 | Rep. 6 | 15,471 | 9528 | |||
| Rep. 7 | 2014 | 2160 | Rep. 7 | 8789 | 12,956 | |||
| Rep. 8 | 2258 | 1054 | Rep. 8 | 8342 | 14,005 | |||
| GeoMean | 1585 | 1912 | GeoMean | 9770 | 13,721 | |||
| SE | 168 | 201 | SE | 1219 | 1250 | |||
| SE% | 11 | 11 | SE% | 12 | 9 | |||
| S.Typhimurium strain | Mouse standard antiserum | Rep. 1 | 28,133 | 16,038 | Anti-rabbit standard serum | Rep. 1 | 61,185 | 25,018 |
| Rep. 2 | 21,515 | 23,282 | Rep. 2 | 40,788 | 24,404 | |||
| Rep. 3 | 21,259 | 9986 | Rep. 3 | 27,883 | 25,895 | |||
| Rep. 4 | 22,669 | 14,926 | Rep. 4 | 28,813 | 23,882 | |||
| Rep. 5 | 25,324 | 18,654 | Rep. 5 | 44,251 | 23,305 | |||
| Rep. 6 | 23,509 | 9197 | Rep. 6 | 27,725 | 21,824 | |||
| Rep. 7 | 23,182 | 9208 | Rep. 7 | 26,312 | 16,567 | |||
| Rep. 8 | 8749 | Rep. 8 | 63,920 | 23,675 | ||||
| GeoMean | 23,557 | 12,905 | GeoMean | 37,765 | 22,888 | |||
| SE | 783 | 1777 | SE | 5077 | 956 | |||
| SE% | 3 | 14 | SE% | 13 | 4 | |||
| S.Enteritidis strain | Mouse standard antiserum | Rep. 1 | 7984 | 18,043 | Rabbit standard antiserum | Rep. 1 | 14,317 | 8527 |
| Rep. 2 | 8327 | 11,270 | Rep. 2 | 8448 | 9505 | |||
| Rep. 3 | 6037 | 16,781 | Rep. 3 | 8931 | 8681 | |||
| Rep. 4 | 7377 | 13,950 | Rep. 4 | 22,548 | 27,381 | |||
| Rep. 5 | 5105 | 15,416 | Rep. 5 | 25,966 | 25,482 | |||
| Rep. 6 | 4482 | 8436 | Rep. 6 | 22,255 | 9927 | |||
| Rep. 7 | 7463 | 8867 | Rep. 7 | 9179 | 9091 | |||
| Rep. 8 | 7867 | 8457 | Rep. 8 | 8701 | 8442 | |||
| GeoMean | 6683 | 12,116 | GeoMean | 13,532 | 11,793 | |||
| SE | 475 | 1293 | SE | 2448 | 2675 | |||
| SE% | 7 | 11 | SE% | 18 | 23 | |||
| S.Paratyphi strain | Mouse standard antiserum | Rep. 1 | 6012 | 6008 | Rabbit standard antiserum | Rep. 1 | 1400 | 941 |
| Rep. 2 | 2962 | 4244 | Rep. 2 | 851 | 1296 | |||
| Rep. 3 | 6090 | 6353 | Rep. 3 | 865 | 974 | |||
| Rep. 4 | 7579 | 6832 | Rep. 4 | 968 | 998 | |||
| Rep. 5 | 9471 | 5474 | Rep. 5 | 1748 | 1361 | |||
| Rep. 6 | 8381 | 5657 | Rep. 6 | 1065 | 775 | |||
| Rep. 7 | 4740 | 4694 | Rep. 7 | 924 | 1201 | |||
| Rep. 8 | 6259 | 7945 | Rep. 8 | 912 | 1026 | |||
| GeoMean | 6107 | 5798 | GeoMean | 1058 | 1056 | |||
| SE | 680 | 389 | SE | 105 | 65 | |||
| SE% | 11 | 7 | SE% | 10 | 6 | |||
| C. freundiistrain | Mouse standard antiserum | Rep. 1 | 11,600 | 12,664 | Rabbit standard antiserum | Rep. 1 | 1216 | 959 |
| Rep. 2 | 9214 | 10,450 | Rep. 2 | 1579 | 1317 | |||
| Rep. 3 | 14,273 | 11,422 | Rep. 3 | 1693 | 1503 | |||
| Rep. 4 | 9719 | 12,792 | Rep. 4 | 1665 | 1490 | |||
| Rep. 5 | 10,345 | 14,563 | Rep. 5 | 1634 | 1588 | |||
| Rep. 6 | 8914 | 12,121 | Rep. 6 | 1629 | 1222 | |||
| Rep. 7 | 7585 | 11,946 | Rep. 7 | 1539 | 1300 | |||
| Rep. 8 | 12,104 | Rep. 8 | 1195 | |||||
| GeoMean | 10,052 | 12,209 | GeoMean | 1557 | 1307 | |||
| SE | 710 | 393 | SE | 53 | 67 | |||
| SE% | 7 | 3 | SE% | 3 | 5 | |||
| Traditional CFU-Based SBA | 96-Wells-Plate L-SBA | 384-Wells-Plate L-SBA | |
|---|---|---|---|
| Final Volume Reaction | 100 µL/well | 100 µL/well | 50 µL/well |
| Assay Time | 1.5 working day | 6 h | 6 h |
| Plate Reading | 2–3 h/SBA plate | 2 min/SBA plate | 5 min/SBA plate |
| Reproducibility | Lower operator independence (for manual CFU counting) than 96- and 384-wells L-SBA | High operator independence | Higher operator independence than 96-well L-SBA |
| Throughput | Plates/day: 2 | Plates/day: 8 | Plates/day: 4 (equivalent to sixteen 96-wells plates) |
| 1 operator/1.5 day: 22 individual sera in single | 1 operator/day: 88 individual sera in single | 1 operator/day: 188 individual sera in single | |
| Reagent Costs | 4 EUR/serum | 12 EUR/serum | 8 EUR/serum |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aruta, M.G.; Carducci, M.; Micoli, F.; Necchi, F.; Rossi, O. Increasing the High Throughput of a Luminescence-Based Serum Bactericidal Assay (L-SBA). BioTech 2021, 10, 19. https://doi.org/10.3390/biotech10030019
Aruta MG, Carducci M, Micoli F, Necchi F, Rossi O. Increasing the High Throughput of a Luminescence-Based Serum Bactericidal Assay (L-SBA). BioTech. 2021; 10(3):19. https://doi.org/10.3390/biotech10030019
Chicago/Turabian StyleAruta, Maria Grazia, Martina Carducci, Francesca Micoli, Francesca Necchi, and Omar Rossi. 2021. "Increasing the High Throughput of a Luminescence-Based Serum Bactericidal Assay (L-SBA)" BioTech 10, no. 3: 19. https://doi.org/10.3390/biotech10030019
APA StyleAruta, M. G., Carducci, M., Micoli, F., Necchi, F., & Rossi, O. (2021). Increasing the High Throughput of a Luminescence-Based Serum Bactericidal Assay (L-SBA). BioTech, 10(3), 19. https://doi.org/10.3390/biotech10030019