
Morphological and Viscoelastic Properties of the Cicada Tymbal



Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Collection and Tymbal Preparation
2.2. Scanning Electron Microscopy (SEM)
2.3. Fluorescence Microscopy
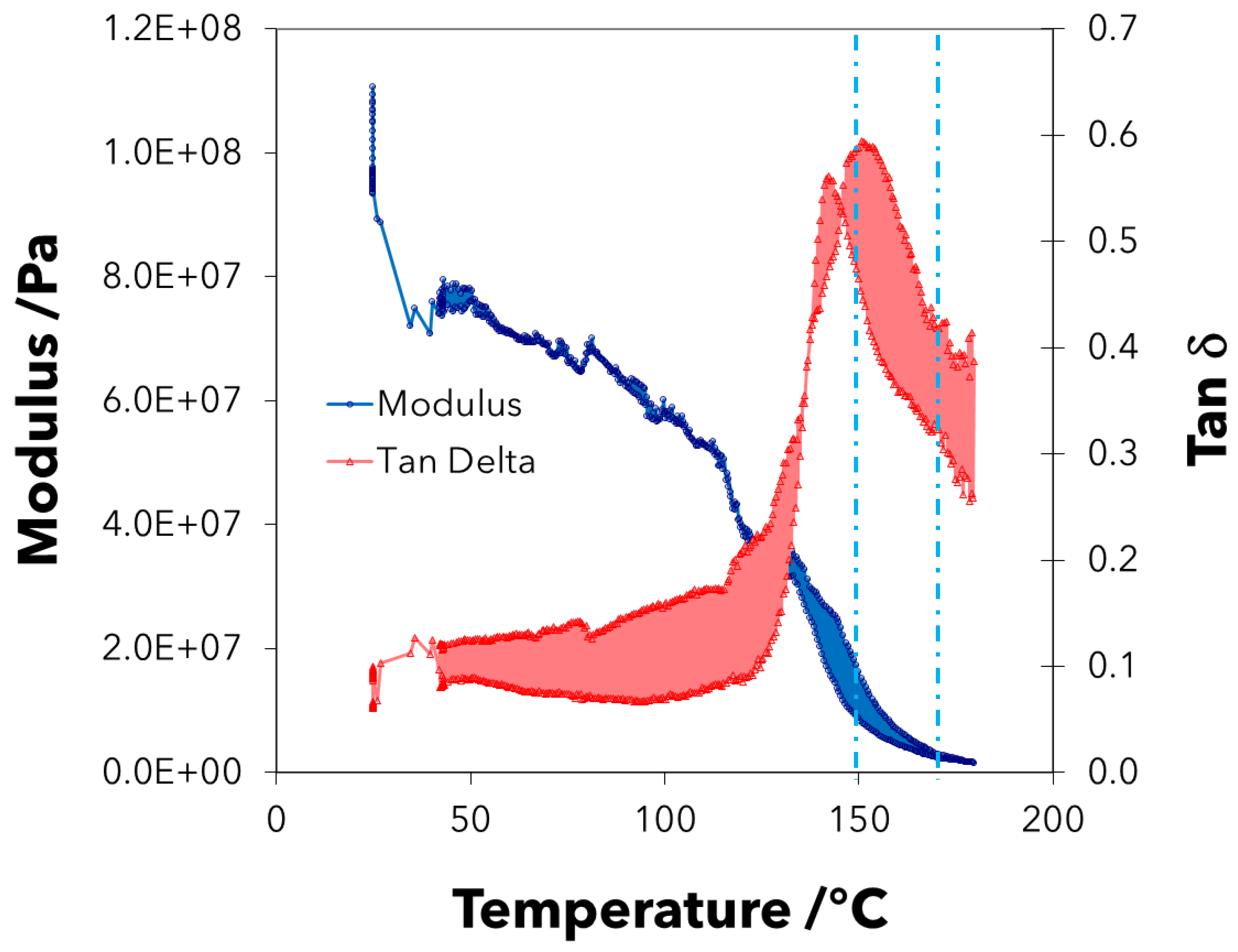
2.4. Dynamic Mechanical Thermal Analysis (DMTA)
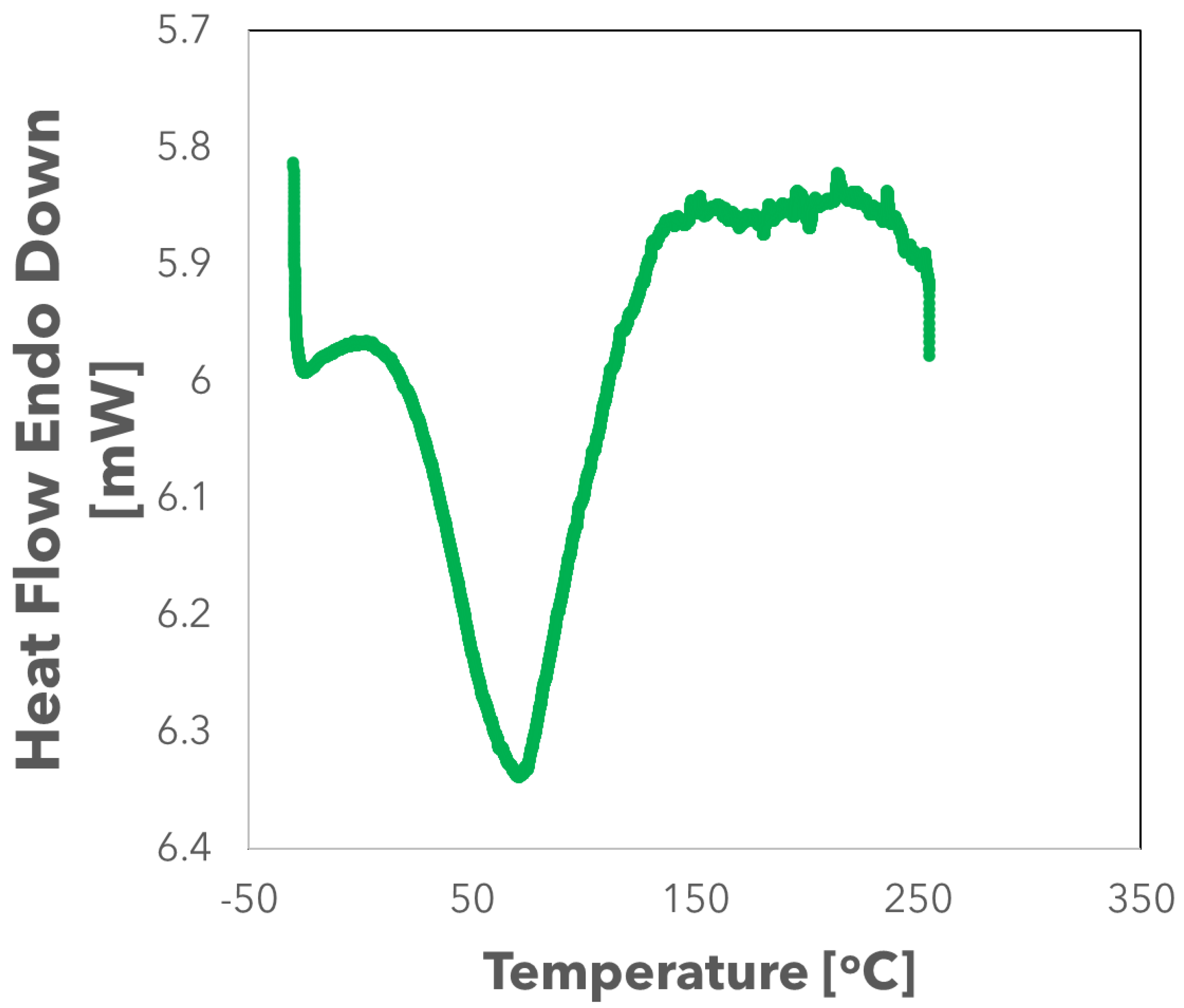
2.5. Differential Scanning Calorimetry (DSC)
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennet-Clark, H.C. How Cicadas Make Their Noise. Sci. Am. 1998, 278, 58–61. [Google Scholar] [CrossRef]
- Young, D. Do cicadas radiate sound through their ear-drums? J. Exp. Biol. 1990, 151, 41–56. [Google Scholar] [CrossRef]
- Young, D.; Bennet-Clark, H.C. The role of the tymbal in cicada sound production. J. Exp. Biol. 1995, 198, 1001–1019. [Google Scholar] [CrossRef] [PubMed]
- Bennet-Clark, H.C.; Young, D. A model of the mechanism of sound production in cicadas. J. Exp. Biol. 1992, 173, 123–153. [Google Scholar] [CrossRef]
- Schroeder, T.B.; Houghtaling, J.; Wilts, B.D.; Mayer, M. It’s not a bug, it’s a feature: Functional materials in insects. Adv. Mater. 2018, 30, 1705322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelsen, A.; Nocke, H. Biophysical aspects of sound communication in insects. In Advances in Insect Physiology; Treherne, J.E., Berridge, M.J., Wigglesworth, V.B., Eds.; Academic Press: London, UK, 1974; Volume 10, pp. 247–296. [Google Scholar]
- Bennet-Clark, H.C. Tymbal mechanics and the control of song frequency in the cicada Cyclochila australasiae. J. Exp. Biol. 1997, 200, 1681–1694. [Google Scholar] [CrossRef]
- Qin, G.; Hu, X.; Cebe, P.; Kaplan, D.L. Mechanism of resilin elasticity. Nat. Commun. 2012, 3, 1003. [Google Scholar] [CrossRef] [Green Version]
- Josephson, R.K.; Young, D. Body temperature and singing in the bladder cicada, Cystosoma saundersii. J. Exp. Biol. 1979, 80, 69–81. [Google Scholar] [CrossRef]
- Fonseca, P.J.; Popov, A.V. Sound radiation in a cicada: The role of different structures. J. Comp. Physiol. A 1994, 175, 349–361. [Google Scholar] [CrossRef]
- Fauziyah, S.; Alam, C.; Soesilohadi, R.C.H.; Retnoaji, B.; Alam, P. Morphological and mechanical characterisation of the hindwing nodus from the Libellulidae family of dragonfly (Indonesia). Arthropod Struct. Dev. 2014, 43, 415–422. [Google Scholar] [CrossRef]
- Fauziyah, S.; Soesilohadi, R.C.H.; Retnoaji, B.; Alam, P. Dragonfly wing venous cross-joints inspire the design of higher-performance bolted timber truss joints. Compos. Part B Eng. 2016, 87, 274–280. [Google Scholar] [CrossRef]
- Gorb, S.N. The jumping mechanism of cicada Cercopis vulnerata (Auchenorrhyncha, Cercopidae): Skeleton–muscle organisation, frictional surfaces, and inverse-kinematic model of leg movements. Arthropod Struct. Dev. 2004, 33, 201–220. [Google Scholar] [CrossRef]
- Chandran, R.; Williams, L.; Hung, A.; Nowlin, K.; LaJeunesse, D. SEM characterization of anatomical variation in chitin organization in insect and arthropod cuticles. Micron 2016, 82, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, E.C.; Kaplan, D.L. Preliminary characterization of resilin isolated from the cockroach, Periplaneta americana. In Materials Research Society Symposium Proceedings; Viney, C., Case, S.T., Waite, J.H., Eds.; Materials Research Society: Pittsburgh, PA, USA, 1993; pp. 3–7. [Google Scholar]
- Andersen, S.O. Characterization of a new type of cross-linkage in resilin, a rubber-like protein. Biochim. Biophys. Acta 1963, 69, 249–262. [Google Scholar] [CrossRef]
- Vincent, J.F.V.; Wegst, U.G.K. Design and mechanical properties of insect cuticle. Arthropod Struct. Dev. 2004, 33, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Weis-Fogh, T. Molecular Interpretation of the Elasticity of Resilin, a Rubber-like Protein. J. Mol. Biol. 1961, 3, 648–667. [Google Scholar] [CrossRef]
- Weis-Fogh, T. Thermodynamic Properties of Resilin, a Rubber-like Protein. J. Mol. Biol. 1961, 3, 520–531. [Google Scholar] [CrossRef]
- Su, R.S.; Kim, Y.; Liu, J.C. Resilin: Protein-based elastomeric biomaterials. Acta Biomater. 2014, 10, 1601–1611. [Google Scholar] [CrossRef]
- Gosline, M. Elastic properties of rubber-like proteins and highly extensive tissues. In Mechanical Properties of Biological Materials; Vincent, J.F.V., Currey, J.D., Eds.; Cambridge University Press: Cambridge, UK, 1980; pp. 331–357. [Google Scholar]
- King, R.J. Dynamic Mechanical Properties of Resilin. Ph.D. Dissertation, Virginia Tech, Blacksburg, VA, USA, 2010. [Google Scholar]
- Choudhury, U. Dynamic Mechanical Properties of Cockroach (Periplaneta Americana) Resilin. Ph.D. Dissertation, Virginia Tech, Blacksburg, VA, USA, 2012. [Google Scholar]
- Qin, G.; Lapidot, S.; Numata, K.; Hu, X.; Meirovitch, S.; Dekel, M.; Podoler, I.; Shoseyov, O.; Kaplan, D.L. Expression, Cross-Linking, and Characterization of Recombinant Chitin Binding Resilin. Biomacromolecules 2009, 10, 3227–3234. [Google Scholar] [CrossRef]
- Mushi, N.E. Chitin nanofibers, networks and composites—Preparation, structure and mechanical properties. Ph.D. Thesis, KTH, Royal Institute of Technology, Stockholm, Sweden, 2014. [Google Scholar]
- Kaya, M.; Mujtaba, M.; Ehrlich, H.; Salaberria, A.M.; Baran, T.; Amemiya, C.T.; Galli, R.; Akyuz, L.; Sargin, I.; Labidi, J. On chemistry of γ-chitin. Carbohydr. Polym. 2017, 176, 177–186. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. The insect cuticle. Biol. Rev. Camb. Philos. Soc. 1948, 23, 408–451. [Google Scholar] [CrossRef] [PubMed]
- Appel, E.; Heepe, L.; Lin, C.P.; Gorb, S.N. Ultrastructure of dragonfly wing veins: Composite structure of fibrous material supplemented by resilin. J. Anat. 2015, 227, 561–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrow, M.; Shaw, S.R.; Sutton, G.P. Resilin and chitinous cuticle form a composite structure for energy storage in jumping by froghopper insects. BMC Biol. 2008, 6, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirks, J.H.; Parle, E.; Taylor, D. Fatigue of insect cuticle. J. Exp. Biol. 2013, 216, 1924–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Frequency | Displacement Imposed | Average Storage Modulus |
|---|---|---|
| [Hz] | [mm] | GPa |
| 1 | 0.01 | 0.132 |
| 1 | 0.02 | 0.126 |
| 1 | 0.05 | 0.114 |
| 10 | 0.01 | 0.146 |
| 10 | 0.02 | 0.138 |
| 10 | 0.05 | 0.101 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fahrunnida; Sayekti, P.R.; Robert, C.; Retnoaji, B.; Alam, P. Morphological and Viscoelastic Properties of the Cicada Tymbal. Macromol 2022, 2, 315-323. https://doi.org/10.3390/macromol2030020
Fahrunnida, Sayekti PR, Robert C, Retnoaji B, Alam P. Morphological and Viscoelastic Properties of the Cicada Tymbal. Macromol. 2022; 2(3):315-323. https://doi.org/10.3390/macromol2030020
Chicago/Turabian StyleFahrunnida, Puspa Restu Sayekti, Colin Robert, Bambang Retnoaji, and Parvez Alam. 2022. "Morphological and Viscoelastic Properties of the Cicada Tymbal" Macromol 2, no. 3: 315-323. https://doi.org/10.3390/macromol2030020
APA StyleFahrunnida, Sayekti, P. R., Robert, C., Retnoaji, B., & Alam, P. (2022). Morphological and Viscoelastic Properties of the Cicada Tymbal. Macromol, 2(3), 315-323. https://doi.org/10.3390/macromol2030020