
Does the Urban Environment Act as a Filter on the Individual Quality of Birds?

 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Animal Sampling
2.2. Relative Telomere Length Analysis
2.3. Statistical Analysis
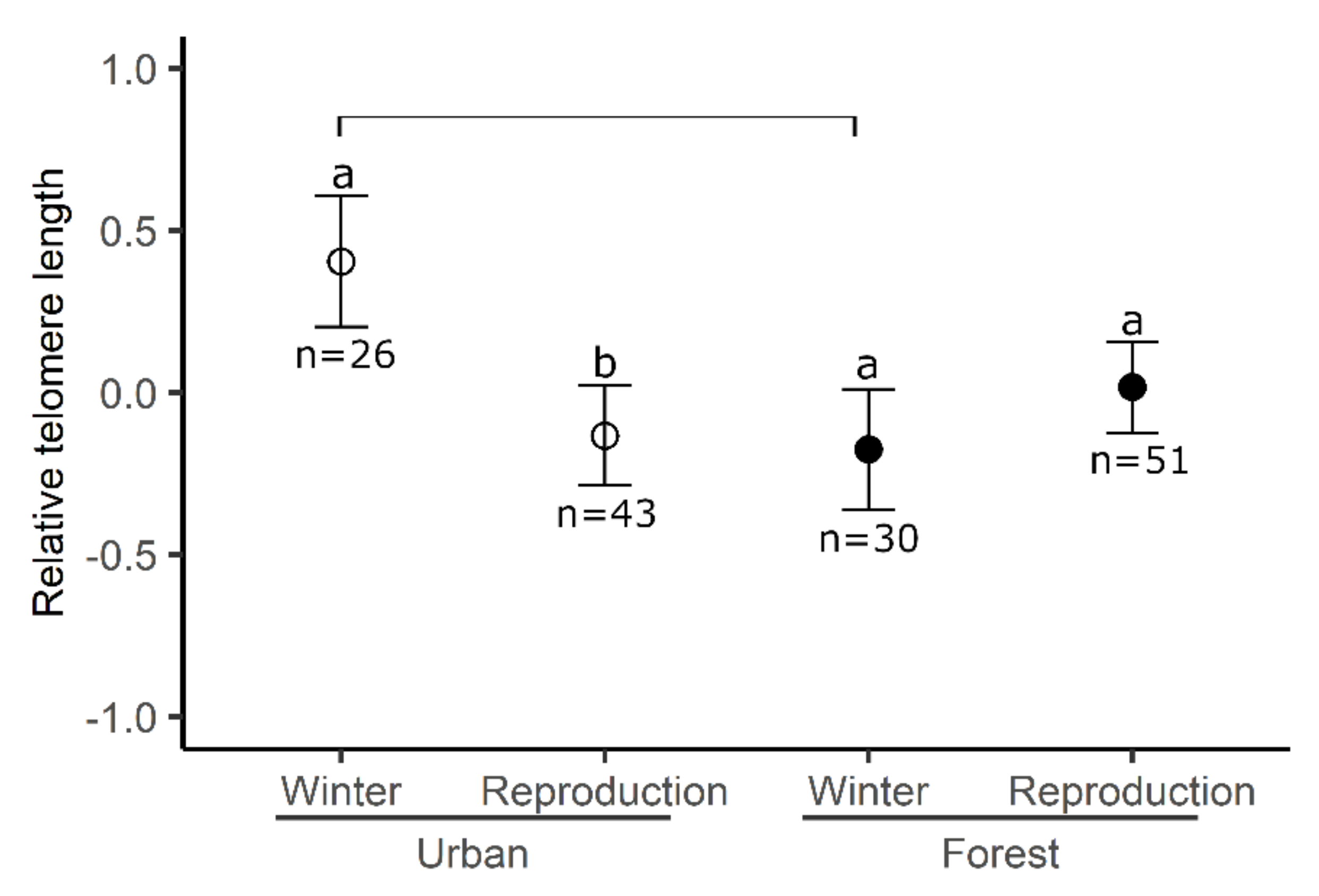
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Site | Groupe | Coordinates (Lat. Long.) | Type | Nb Nestboxes |
|---|---|---|---|---|
| Urban | Campus 1 | 48.5843 N; 7.7637 W | Urban square | 26 |
| Campus 2 | 48.5794 N; 7.7677W | Busy road | 11 | |
| Citadelle | 48.5759 N; 7.7748 W | Urban park | 40 | |
| Heyritz | 48.5727 N; 7.7442 W | Urban square | 15 | |
| Forest | La Wantzenau | 48.6480 N; 7.8337 W | Alluvial forest | 65 |
Appendix B
| Urban | Forest | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Winter | Reproduction | Winter | Reproduction | |||||||||
| F | M | Total | F | M | Total | F | M | Total | F | M | Total | |
| 2 years old | 4 | 7 | 11 | 16 | 5 | 21 | 8 | 6 | 14 | 20 | 9 | 29 |
| >2 years old | 5 | 12 | 17 | 9 | 18 | 27 | 4 | 12 | 16 | 12 | 14 | 26 |
| Total | 9 | 19 | 28 | 25 | 23 | 48 | 12 | 18 | 30 | 32 | 23 | 55 |
Appendix C
| Forward | Reverse | |
|---|---|---|
| Control gene P. major Zinc finger protein (GenBank:EF568148) | ZENK1: 5′-TACATGTGCCATGGTTTTGC-3′ | ZENK2:5′-AAGTGCTGCTCCCAAAGAAG-3′ |
| Telomeres | Tel1b: 5′-CGGTTTGTTTGGGTTTGGGTTTGGGTTTGGGTTTGGGTT-3′ | Tel2b: 5′-GGCTTGCCTTACCCTTACCCTTACCCTACCCTTACCCT-3′ |
Appendix D. Estimates of Generalized Linear Model Used for Morphological and Physiological Parameters
| Age | Body Size | Body Condition | Relative Telomere Length | |
|---|---|---|---|---|
| Estimate (SE) | Estimate (SE) | Estimate (SE) | Estimate (SE) | |
| Intercept | −9.17 (1.96) | −0.9 (0.2) | 0.07 (0.17) | 0.27 (0.25) |
| Site [Forest] | −0.65 (1.66) | 1.05 (0.21) | −0.1 (0.13) | −0.58 (0.27) |
| Sex [M] | 20.67 (2.67) | 1.11 (0.17) | −0.18 (0.14) | 0.11 (0.18) |
| Season [Reproduction] | −0.92 (1.45) | −0.29 (0.1) | 0.23 (0.11) | −0.54 (0.25) |
| Age [2 years old] | −0.43 (0.17) | −0.15 (0.14) | 0.16 (0.17) | |
| Body condition | 0.08 (0.11) | |||
| Site [Forest]:Season [Reproduction] | 0.29 (0.14) | 0.73 (0.34) |
| Body Condition | Relative Telomere Length | |
|---|---|---|
| Estimate (SE) | Estimate (SE) | |
| Intercept | −1.83 (2.62) | −3.83 (2.46) |
| Site [Forest] | −0.14 (0.63) | 0.25 (0.61) |
| Sex [M] | 0.27 (0.38) | −0.35 (0.34) |
| Fledging number | 0.08 (0.09) | −0.06 (0.07) |
| Hatching date | 0.02 (0.05) | 0.08 (0.05) |
| Age [2 years old.] | 0.56 (0.37) | −0.09 (0.35) |
| RTL Winter | −0.03 (0.23) |
References
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Lailvaux, S.P.; Husak, J.F. The life history of whole-organism performance. Q. Rev. Biol. 2014, 89, 285–318. [Google Scholar] [CrossRef] [PubMed]
- McNamara, J.M.; Houston, A.I. State-dependent life histories. Nature 1996, 380, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Sudyka, J. Does reproduction shorten telomeres? Towards integrating individual quality with life-history strategies in telomere biology. BioEssays 2019, 41, 1900095. [Google Scholar] [CrossRef]
- Wilson, A.J.; Nussey, D.H. What is individual quality? An evolutionary perspective. Trends Ecol. Evol. 2010, 25, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Blums, P.; Nichols, J.D.; Hines, J.E.; Lindberg, M.S.; Mednis, A. Individual quality, survival variation and patterns of phenotypic selection on body condition and timing of nesting in birds. Oecologia 2005, 143, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Vedder, O.; Bouwhuis, S. Heterogeneity in individual quality in birds: Overall patterns and insights from a study on common terns. Oikos 2018, 127, 719–727. [Google Scholar] [CrossRef]
- Peralta-Sánchez, J.M.; Colmenero, J.; Redondo-Sánchez, S.; Ontanilla, J.; Soler, M. Females are more determinant than males in reproductive performance in the house sparrow Passer domesticus. J. Avian Biol. 2020, 51, e02240. [Google Scholar] [CrossRef]
- Dukas, R. Animal expertise: Mechanisms, ecology and evolution. Anim. Behav. 2019, 147, 199–210. [Google Scholar] [CrossRef]
- Forslund, P.; Pärt, T. Age and reproduction in birds—Hypotheses and tests. Trends Ecol. Evol. 1995, 10, 374–378. [Google Scholar] [CrossRef]
- Pitera, A.M.; Branch, C.L.; Sonnenberg, B.R.; Benedict, L.M.; Kozlovsky, D.Y.; Pravosudov, V.V. Reproduction is affected by individual breeding experience but not pair longevity in a socially monogamous bird. Behav. Ecol. Sociobiol. 2021, 75, 1–11. [Google Scholar] [CrossRef]
- Espie, R.H.M.; Oliphant, L.W.; James, P.C.; Warkentin, I.G.; Lieske, D.J. Age-dependent breeding performance in Merlins (Falco columbarius). Ecology 2000, 81, 3404–3415. [Google Scholar] [CrossRef]
- Hamman, J.; Cooke, F. Age effects on clutch size and laying dates of individual female Lesser Snow Geese Anser caerulescens. Ibis 2008, 129, 527–532. [Google Scholar] [CrossRef]
- Wiktander, U.; Olsson, O.; Nilsson, S.G. Age and Reproduction in Lesser Spotted Woodpeckers (Dendrocopos minor). Auk 2001, 118, 624–635. [Google Scholar] [CrossRef]
- Hatch, M.I.; Westneat, D.F. Age-related patterns of reproductive success in house sparrows Passer domesticus. J. Avian Biol. 2007, 38, 603–611. [Google Scholar] [CrossRef]
- Riechert, J.; Chastel, O.; Becker, P.H. Why do experienced birds reproduce better? Possible endocrine mechanisms in a long-lived seabird, the common tern. Gen. Comp. Endocrinol. 2012, 178, 391–399. [Google Scholar] [CrossRef]
- Isaksson, C.; Andersson, S. Carotenoid diet and nestling provisioning in urban and rural Great Tits Parus major. J. Avian Biol. 2007, 38, 564–572. [Google Scholar] [CrossRef]
- Biard, C.; Brischoux, F.; Meillère, A.; Michaud, B.; Nivière, M.; Ruault, S.; Vaugoyeau, M.; Angelier, F. Growing in cities: An urban penalty for wild birds? A study of phenotypic differences between urban and rural Great Tit chicks (Parus major). Front. Ecol. Evol. 2017, 5, 1–14. [Google Scholar] [CrossRef]
- Vaugoyeau, M.; Adriaensen, F.; Artemyev, A.; Bańbura, J.; Barba, E.; Biard, C.; Blondel, J.; Bouslama, Z.; Bouvier, J.C.; Camprodon, J.; et al. Interspecific variation in the relationship between clutch size, laying date and intensity of urbanization in four species of hole-nesting birds. Ecol. Evol. 2016, 6, 5907–5920. [Google Scholar] [CrossRef]
- Gil, D.; Brumm, H. (Eds.) Avian Urban Ecology; Oxford University Press: Oxford, UK, 2013; ISBN 9780199661572. [Google Scholar]
- Jones, E.L.; Leather, S.R. Invertebrates in urban areas: A review. Eur. J. Entomol. 2012, 109, 463–478. [Google Scholar] [CrossRef]
- Fenoglio, M.S.; Rossetti, M.R.; Videla, M. Negative effects of urbanization on terrestrial arthropod communities: A meta-analysis. Glob. Ecol. Biogeogr. 2020, 29, 1412–1429. [Google Scholar] [CrossRef]
- MacLeod, R.; Barnett, P.; Clark, J.; Cresswell, W. Mass-dependent predation risk as a mechanism for house sparrow declines? Biol. Lett. 2006, 2, 43–46. [Google Scholar] [CrossRef]
- Marzluff, J.M. Worldwide urbanization and its effects on birds. In Avian Ecology and Conservation in an Urbanizing World; Springer: Boston, MA, USA, 2001; pp. 19–47. [Google Scholar]
- Sorace, A.; Gustin, M. Distribution of generalist and specialist predators along urban gradients. Landsc. Urban Plan. 2009, 90, 111–118. [Google Scholar] [CrossRef]
- Rodewald, A.D.; Kearns, L.J. Shifts in dominant nest predators along a rural-to-urban landscape gradient. Condor 2011, 113, 899–906. [Google Scholar] [CrossRef]
- Van Heezik, Y.; Smyth, A.; Adams, A.; Gordon, J. Do domestic cats impose an unsustainable harvest on urban bird populations? Biol. Conserv. 2010, 143, 121–130. [Google Scholar] [CrossRef]
- Chamberlain, D.E.; Cannon, A.R.; Toms, M.P.; Leech, D.I.; Hatchwell, B.J.; Gaston, K.J. Avian productivity in urban landscapes: A review and meta-analysis. Ibis 2009, 151, 1–18. [Google Scholar] [CrossRef]
- Sepp, T.; McGraw, K.J.; Kaasik, A.; Giraudeau, M. A review of urban impacts on avian life-history evolution: Does city living lead to slower pace of life? Glob. Chang. Biol. 2018, 24, 1452–1469. [Google Scholar] [CrossRef]
- Wawrzyniak, J.; Glądalski, M.; Kaliński, A.; Bańbura, M.; Markowski, M.; Skwarska, J.; Zieliński, P.; Bańbura, J. Differences in the breeding performance of Great Tits Parus major between a forest and an urban area: A long term study on first clutches. Eur. Zool. J. 2020, 87, 294–309. [Google Scholar] [CrossRef]
- Halfwerk, W.; Holleman, L.J.M.; Lessells, C.M.; Slabbekoorn, H. Negative impact of traffic noise on avian reproductive success. J. Appl. Ecol. 2011, 48, 210–219. [Google Scholar] [CrossRef]
- Bailly, J.; Scheifler, R.; Berthe, S.; Clément-Demange, V.A.; Leblond, M.; Pasteur, B.; Faivre, B. From eggs to fledging: Negative impact of urban habitat on reproduction in two tit species. J. Ornithol. 2016, 157, 377–392. [Google Scholar] [CrossRef]
- Peach, W.J.; Mallord, J.W.; Ockendon, N.; Orsman, C.J.; Haines, W.G. Invertebrate prey availability limits reproductive success but not breeding population size in suburban House Sparrows Passer domesticus. Ibis 2015, 157, 601–613. [Google Scholar] [CrossRef]
- Bailly, J.; Faivre, B.; Bernard, N.; Sage, M.; Crini, N.; Driget, V.; Garnier, S.; Rieffel, D.; Scheifler, R. Multi-element analysis of blood samples in a passerine species: Excesses and deficiencies of trace elements in an urbanization study. Front. Ecol. Evol. 2017, 5, 6. [Google Scholar] [CrossRef]
- Isaksson, C.; Johansson, A.; Andersson, S. Egg yolk carotenoids in relation to habitat and reproductive investment in the Great Tit Parus major. Physiol. Biochem. Zool. 2008, 81, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, C.; Powell, L.L.; McDevitt, H.; Helm, B.; Welch, A.J. Bitter fruits of hard labour: Diet metabarcoding and telemetry reveal that urban songbirds travel further for lower-quality food. Oecologia 2020, 193, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Hinsley, S.A.; Hill, R.A.; Bellamy, P.E.; Harrison, N.M.; Speakman, J.R.; Wilson, A.K.; Ferns, P.N. Effects of structural and functional habitat gaps on breeding woodland birds: Working harder for less. Landsc. Ecol. 2008, 23, 615–626. [Google Scholar] [CrossRef]
- Estes, W.A.; Mannan, R.W. Feeding behavior of cooper’s hawks at urban and rural nests in southeastern arizona. Condor 2003, 105, 107–116. [Google Scholar] [CrossRef]
- Blair, R.B. Land use and avian species diversity along an urban gradient. Ecol. Appl. 1996, 6, 506–519. [Google Scholar] [CrossRef]
- Clergeau, P.; Croci, S.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.L.; Dinetti, M. Avifauna homogenisation by urbanisation: Analysis at different European latitudes. Biol. Conserv. 2006, 127, 336–344. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Croci, S.; Butet, A.; Clergeau, P. Does urbanization filter birds on the basis of their biological traits? Condor 2008, 110, 223–240. [Google Scholar] [CrossRef]
- Kurucz, K.; Purger, J.J.; Batáry, P. Urbanization shapes bird communities and nest survival, but not their food quantity. Glob. Ecol. Conserv. 2021, 26, e01475. [Google Scholar] [CrossRef]
- Liker, A.; Papp, Z.; Bókony, V.; Lendvai, Á.Z. Lean birds in the city: Body size and condition of house sparrows along the urbanization gradient. J. Anim. Ecol. 2008, 77, 789–795. [Google Scholar] [CrossRef]
- Meillère, A.; Brischoux, F.; Parenteau, C.; Angelier, F. Influence of urbanization on body size, condition, and physiology in an urban exploiter: A multi-component approach. PLoS ONE 2015, 10, 1–19. [Google Scholar] [CrossRef]
- Evans, K.L.; Gaston, K.J.; Sharp, S.P.; McGowan, A.; Hatchwell, B.J. The effect of urbanisation on avian morphology and latitudinal gradients in body size. Oikos 2009, 118, 251–259. [Google Scholar] [CrossRef]
- Shochat, E. Credit or debit? Resource input changes population dynamics of city-slicker birds. Oikos 2004, 106, 622–626. [Google Scholar] [CrossRef]
- Møller, A.P. Successful city dwellers: A comparative study of the ecological characteristics of urban birds in the Western Palearctic. Oecologia 2009, 159, 849–858. [Google Scholar] [CrossRef]
- Caizergues, A.E.; Charmantier, A.; Lambrechts, M.M.; Perret, S.; Demeyrier, V.; Lucas, A.; Grégoire, A. An avian urban morphotype: How the city environment shapes Great Tit morphology at different life stages. Urban Ecosyst. 2021, 24, 929–941. [Google Scholar] [CrossRef]
- Meillère, A.; Brischoux, F.; Henry, P.Y.; Michaud, B.; Garcin, R.; Angelier, F. Growing in a city: Consequences on body size and plumage quality in an urban dweller, the house sparrow (Passer domesticus). Landsc. Urban Plan. 2017, 160, 127–138. [Google Scholar] [CrossRef]
- Milenkaya, O.; Catlin, D.H.; Legge, S.; Walters, J.R. Body condition indices predict reproductive success but not survival in a sedentary, tropical bird. PLoS ONE 2015, 10, e0136582. [Google Scholar] [CrossRef]
- Lamb, J.S.; O’Reilly, K.M.; Jodice, P.G.R. Physical condition and stress levels during early development reflect feeding rates and predict pre-And post-fledging survival in a nearshore seabird. Conserv. Physiol. 2016, 4, cow060. [Google Scholar] [CrossRef]
- Hõrak, P.; Tegelmann, L.; Ots, I.; Møller, A.P. Immune function and survival of Great Tit nestlings in relation to growth conditions. Oecologia 1999, 121, 316–322. [Google Scholar] [CrossRef]
- Blackburn, E.H. Structure and function of telomeres. Nature 1991, 350, 569–573. [Google Scholar] [CrossRef]
- Blackburn, E.H. Telomeres and telomerase: Their mechanisms of action and the effects of altering their functions. FEBS Lett. 2005, 579, 859–862. [Google Scholar] [CrossRef]
- Tricola, G.M.; Simons, M.J.P.; Atema, E.; Boughton, R.K.; Brown, J.L.; Dearborn, D.C.; Divoky, G.; Eimes, J.A.; Huntington, C.E.; Kitaysky, A.S.; et al. The rate of telomere loss is related to maximum lifespan in birds. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20160445. [Google Scholar] [CrossRef] [PubMed]
- Haussmann, M.F.; Mauck, R.A. Telomeres and longevity: Testing an evolutionary hypothesis. Mol. Biol. Evol. 2008, 25, 220–228. [Google Scholar] [CrossRef]
- Monaghan, P.; Metcalfe, N.B.; Torres, R. Oxidative stress as a mediator of life history trade-offs: Mechanisms, measurements and interpretation. Ecol. Lett. 2009, 12, 75–92. [Google Scholar] [CrossRef]
- Stauffer, J.; Panda, B.; Eeva, T.; Rainio, M.J.; Ilmonen, P. Telomere damage and redox status alterations in free-living passerines exposed to metals. Sci. Total Environ. 2017, 575, 841–848. [Google Scholar] [CrossRef]
- Blévin, P.; Angelier, F.; Tartu, S.; Bustamante, P.; Herzke, D.; Moe, B.; Bech, C.; Gabrielsen, G.W.; Bustnes, J.O.; Chastel, O. Perfluorinated substances and telomeres in an Arctic seabird: Cross-sectional and longitudinal approaches. Environ. Pollut. 2017, 230, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Meillère, A.; Brischoux, F.; Ribout, C.; Angelier, F. Traffic noise exposure affects telomere length in nestling house sparrows. Biol. Lett. 2015, 11, 20150559. [Google Scholar] [CrossRef] [PubMed]
- Grunst, A.S.; Grunst, M.L.; Bervoets, L.; Pinxten, R.; Eens, M. Proximity to roads, but not exposure to metal pollution, is associated with accelerated developmental telomere shortening in nestling Great Tits. Environ. Pollut. 2020, 256, 113373. [Google Scholar] [CrossRef]
- Young, R.C.; Welcker, J.; Barger, C.P.; Hatch, S.A.; Merkling, T.; Kitaiskaia, E.V.; Haussmann, M.F.; Kitaysky, A.S. Effects of developmental conditions on growth, stress and telomeres in black-legged kittiwake chicks. Mol. Ecol. 2017, 26, 3572–3584. [Google Scholar] [CrossRef]
- Boonekamp, J.J.; Bauch, C.; Mulder, E.; Verhulst, S. Does oxidative stress shorten telomeres? Biol. Lett. 2017, 13, 20170164. [Google Scholar] [CrossRef] [PubMed]
- Chatelain, M.; Drobniak, S.M.; Szulkin, M. The association between stressors and telomeres in non-human vertebrates: A meta-analysis. Ecol. Lett. 2020, 23, 381–398. [Google Scholar] [CrossRef] [PubMed]
- Epel, E.S.; Blackburn, E.H.; Lin, J.; Dhabhar, F.S.; Adler, N.E.; Morrow, J.D.; Cawthon, R.M. Accelerated telomere shortening in response to life stress. Proc. Natl. Acad. Sci. USA 2004, 101, 17312–17315. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Velando, A. Antioxidants safeguard telomeres in bold chicks. Biol. Lett. 2015, 11, 20150211. [Google Scholar] [CrossRef] [PubMed]
- Haussmann, M.F.; Winkler, D.W.; Vleck, C.M. Longer telomeres associated with higher survival in birds. Biol. Lett. 2005, 1, 212–214. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Álamo, J.D.; Pineda-Pampliega, J.; Thomson, R.L.; Aguirre, J.I.; Díez-Fernández, A.; Faivre, B.; Figuerola, J.; Verhulst, S. Urban blackbirds have shorter telomeres. Biol. Lett. 2018, 14, 20180083. [Google Scholar] [CrossRef] [PubMed]
- Salmón, P.; Nilsson, J.F.; Watson, H.; Bensch, S.; Isaksson, C. Selective disappearance of Great Tits with short telomeres in urban areas. Proc. R. Soc. B Biol. Sci. 2017, 284, 20171349. [Google Scholar] [CrossRef]
- Criscuolo, F.; Bize, P.; Nasir, L.; Metcalfe, N.B.; Foote, C.G.; Griffiths, K.; Gault, E.A.; Monaghan, P. Real-time quantitative PCR assay for measurement of avian telomeres. J. Avian Biol. 2009, 40, 342–347. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing, Ed.; R Development Core Team: Vienna, Austria, 2021; ISBN 3-900051-07-0. [Google Scholar]
- Verhulst, S. Improving comparability between qPCR-based telomere studies. Mol. Ecol. Resour. 2020, 20, 11–13. [Google Scholar] [CrossRef]
- Labocha, M.K.; Hayes, J.P. Morphometric indices of body condition in birds: A review. J. Ornithol. 2012, 153, 1–22. [Google Scholar] [CrossRef]
- Hayes, J.P.; Shonkwiler, J.S. Morphological indicators of body condition: Useful or wishful thinking? Body Compos. Anal. Anim. 2001, 10, 8–38. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Chastel, O.; Weimerskirch, H.; Jouventin, P. Influence of body condition on reproductive decision and reproductive success in the Blue Petrel. Auk 1995, 112, 964–972. [Google Scholar] [CrossRef]
- Isaksson, C.; Andersson, M.N.; Nord, A.; Von Post, M.; Wang, H.L. Species-dependent effects of the urban environment on fatty acid composition and oxidative stress in birds. Front. Ecol. Evol. 2017, 5, 1–13. [Google Scholar] [CrossRef]
- Corsini, M.; Schöll, E.M.; Di Lecce, I.; Chatelain, M.; Dubiec, A.; Szulkin, M. Growing in the city: Urban evolutionary ecology of avian growth rates. Evol. Appl. 2020, 14, 69–84. [Google Scholar] [CrossRef]
- Caizergues, A.E.; Grégoire, A.; Charmantier, A. Urban versus forest ecotypes are not explained by divergent reproductive selection. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180261. [Google Scholar] [CrossRef]
- McDonnell, M.J.; Hahs, A.K. Adaptation and Adaptedness of Organisms to Urban Environments. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 261–280. [Google Scholar] [CrossRef]
- Hedenstrom, A.; Møller, A.P. Morphological adaptations to song flight in passerine birds: A comparative study. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1992, 247, 183–187. [Google Scholar] [CrossRef]
- Møller, A.P. Flight distance of urban birds, predation, and selection for urban life. Behav. Ecol. Sociobiol. 2008, 63, 63–75. [Google Scholar] [CrossRef]
- Gosler, A.G.; Greenwood, J.J.D.; Perrins, C. Predation risk and the cost of being fat. Nature 1995, 377, 621–623. [Google Scholar] [CrossRef]
- Kiat, Y.; Sapir, N. Life-history trade-offs result in evolutionary optimization of feather quality. Biol. J. Linn. Soc. 2018, 125, 613–624. [Google Scholar] [CrossRef]
- Pap, P.L.; Barta, Z.; Tökölyi, J.; Vágási, I.C. Increase of feather quality during moult: A possible implication of feather deformities in the evolution of partial moult in the Great Tit Parus major. J. Avian Biol. 2007, 38, 471–478. [Google Scholar] [CrossRef]
- Giraudeau, M.; Angelier, F.; Sepp, T. Do telomeres influence pace-of-life-strategies in response to environmental conditions over a lifetime and between generations? BioEssays 2019, 41, 1800162. [Google Scholar] [CrossRef]
- Heidinger, B.J.; Blount, J.D.; Boner, W.; Griffiths, K.; Metcalfe, N.B.; Monaghan, P. Telomere length in early life predicts lifespan. Proc. Natl. Acad. Sci. USA 2012, 109, 1743–1748. [Google Scholar] [CrossRef]
- Bauch, C.; Becker, P.H.; Verhulst, S. Telomere length reflects phenotypic quality and costs of reproduction in a long-lived seabird. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122540. [Google Scholar] [CrossRef]
- Angelier, F.; Weimerskirch, H.; Barbraud, C.; Chastel, O. Is telomere length a molecular marker of individual quality? Insights from a long-lived bird. Funct. Ecol. 2019, 33, 1076–1087. [Google Scholar] [CrossRef]
- Krijgsveld, K.L.; Visser, G.H.; Daan, S. Foraging behavior and physiological changes in precocial quail chicks in response to low temperatures. Physiol. Behav. 2003, 79, 311–319. [Google Scholar] [CrossRef]
- Avery, M.I.; Krebs, J.R. Temperature and foraging success of Great Tits Parus major hunting for spiders. Ibis 1984, 126, 33–38. [Google Scholar] [CrossRef]
- Atwell, J.W.; Cardoso, G.C.; Whittaker, D.J.; Campbell-Nelson, S.; Robertson, K.W.; Ketterson, E.D. Boldness behavior and stress physiology in a novel urban environment suggest rapid correlated evolutionary adaptation. Behav. Ecol. 2012, 23, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Herborn, K.A.; Coffey, J.; Larcombe, S.D.; Alexander, L.; Arnold, K.E. Oxidative profile varies with personality in European greenfinches. J. Exp. Biol. 2011, 214, 1732–1739. [Google Scholar] [CrossRef] [PubMed][Green Version]


| Age (n = 161) | Body Size Index (n = 161) | Body Condition (n = 161) | RTL (n = 151) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | Chi² | df | p | F | df | p | F | df | p | F | df | p |
| Site | 0.156 | 1, 154 | 0.693 | 45.66 | 1, 153 | <0.001 | 0.6002 | 1, 149 | 0.440 | 0.548 | 1, 142 | 0.461 |
| Sex | 61.35 | 1, 154 | <0.001 | 39.97 | 1, 153 | <0.001 | 1.6297 | 1, 149 | 0.203 | 0.395 | 1, 142 | 0.531 |
| Season | 0.400 | 1, 154 | 0.527 | 4.46 | 1, 153 | 0.042 | 3.9457 | 1, 149 | 0.051 | 0.623 | 1, 142 | 0.432 |
| Age | 5.82 | 1, 153 | 0.017 | 1.1800 | 1, 149 | 0.280 | 0.812 | 1, 142 | 0.370 | |||
| Body condition | 0.577 | 1, 142 | 0.450 | |||||||||
| Site × Age | ||||||||||||
| Site × Season | 4.18 | 1, 153 | 0.048 | 4.525 | 1, 142 | 0.037 | ||||||
| Site × Sex | ||||||||||||
| Body Condition (n = 27) | RTL (n = 23) | |||||
|---|---|---|---|---|---|---|
| Variable | F | df | p | F | df | p |
| Site | 0.048 | 1, 21 | 0.829 | 0.161 | 1, 16 | 0.694 |
| Sex | 0.523 | 1, 21 | 0.478 | 1.105 | 1, 16 | 0.310 |
| Age | 2.287 | 1, 21 | 0.146 | 0.069 | 1, 16 | 0.796 |
| Fledging number | 0.850 | 1, 21 | 0.367 | 0.689 | 1, 16 | 0.419 |
| Hatching date | 0.111 | 1, 21 | 0.742 | 2.778 | 1, 16 | 0.115 |
| RTL Winter | 0.527 | 1, 16 | 0.479 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saulnier, A.; Bleu, J.; Lemonnier, G.; Uhlrich, P.; Zahn, S.; Massemin, S. Does the Urban Environment Act as a Filter on the Individual Quality of Birds? Birds 2022, 3, 84-98. https://doi.org/10.3390/birds3010007
Saulnier A, Bleu J, Lemonnier G, Uhlrich P, Zahn S, Massemin S. Does the Urban Environment Act as a Filter on the Individual Quality of Birds? Birds. 2022; 3(1):84-98. https://doi.org/10.3390/birds3010007
Chicago/Turabian StyleSaulnier, Agnès, Josefa Bleu, Gildas Lemonnier, Pierre Uhlrich, Sandrine Zahn, and Sylvie Massemin. 2022. "Does the Urban Environment Act as a Filter on the Individual Quality of Birds?" Birds 3, no. 1: 84-98. https://doi.org/10.3390/birds3010007
APA StyleSaulnier, A., Bleu, J., Lemonnier, G., Uhlrich, P., Zahn, S., & Massemin, S. (2022). Does the Urban Environment Act as a Filter on the Individual Quality of Birds? Birds, 3(1), 84-98. https://doi.org/10.3390/birds3010007