
Evaluating Potential Indicators of Welfare for Zoo Birds during an Avian Influenza Enforced Housing Order



Abstract

1. Introduction
2. Materials and Methods
2.1. Study Population and Husbandry
2.2. Data Collection
2.3. Data Analyses
2.3.1. Behavioural Analysis
2.3.2. Plumage Condition Analysis
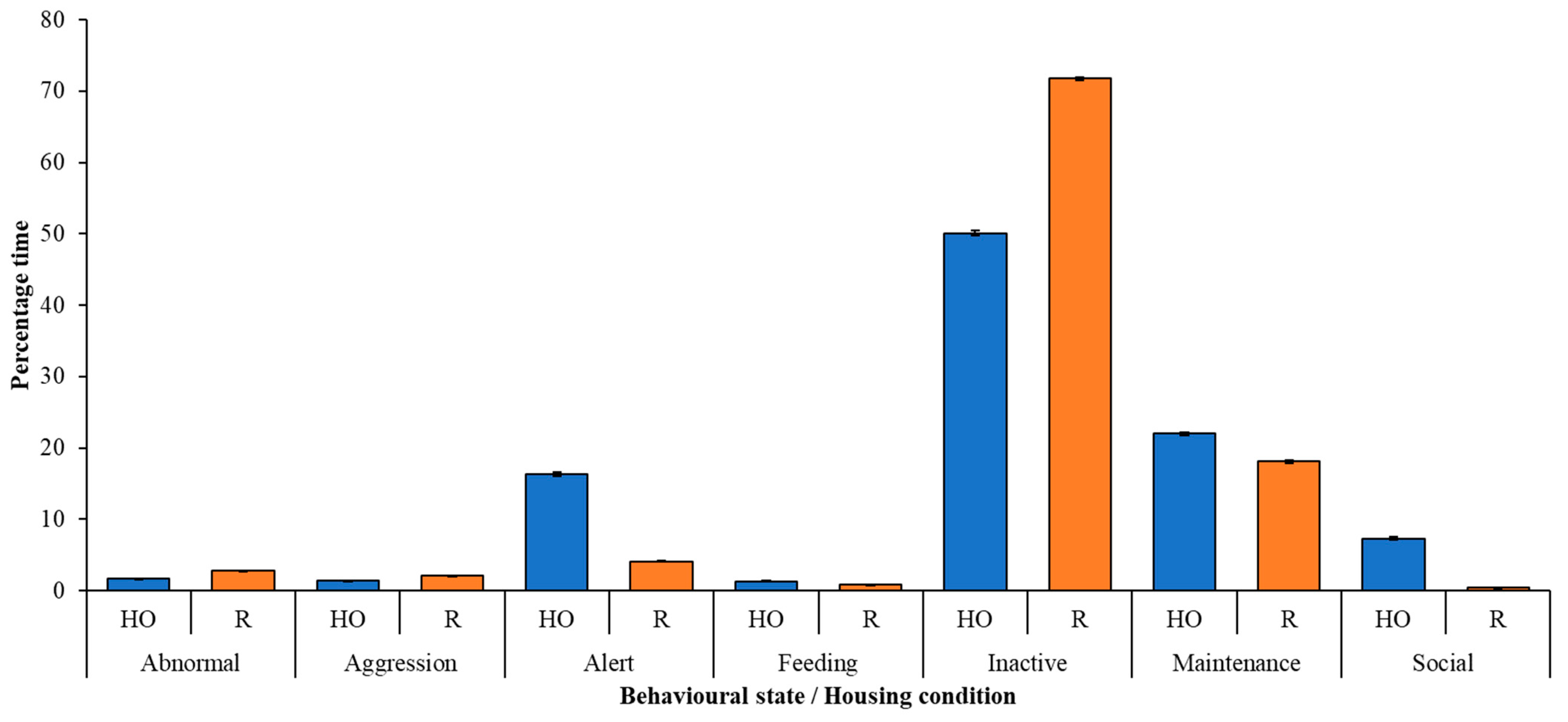
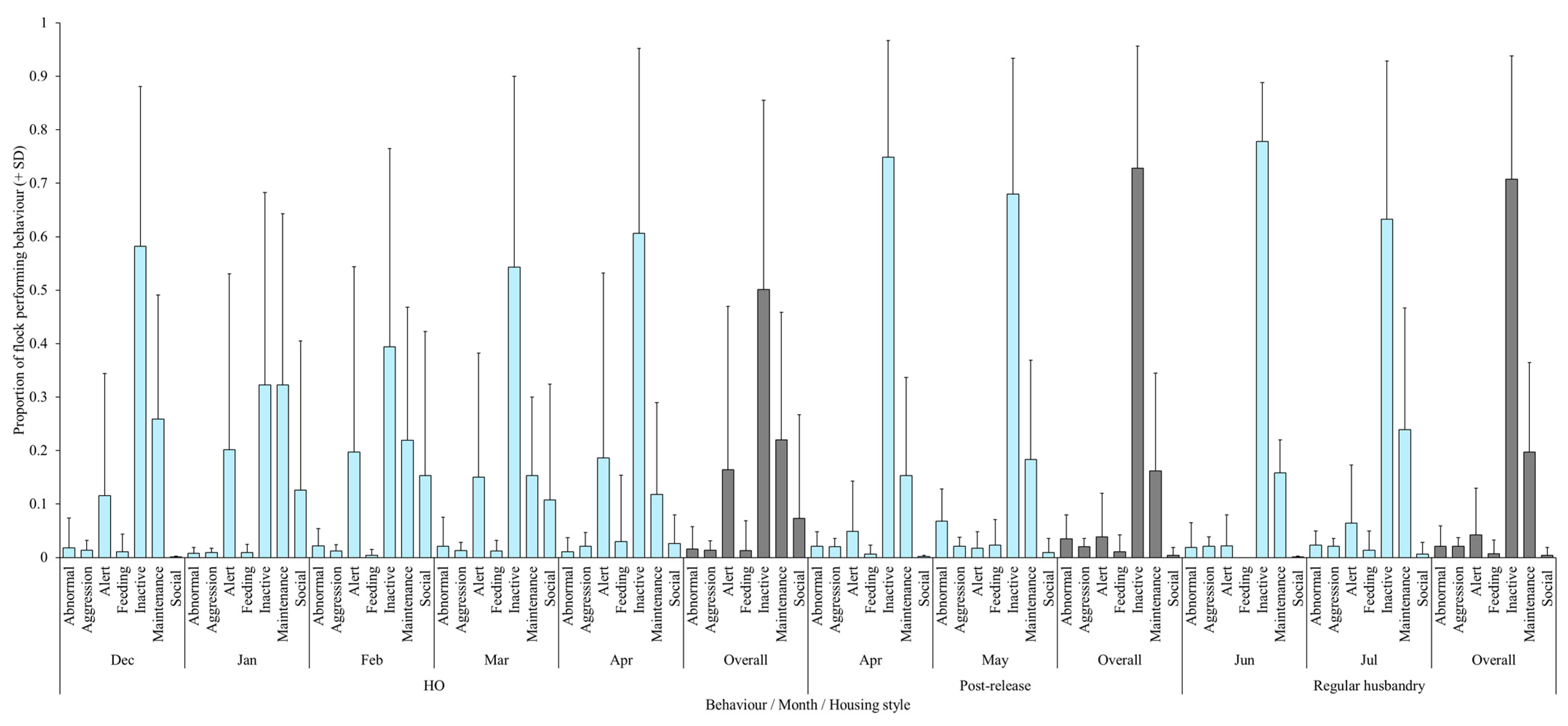
3. Results
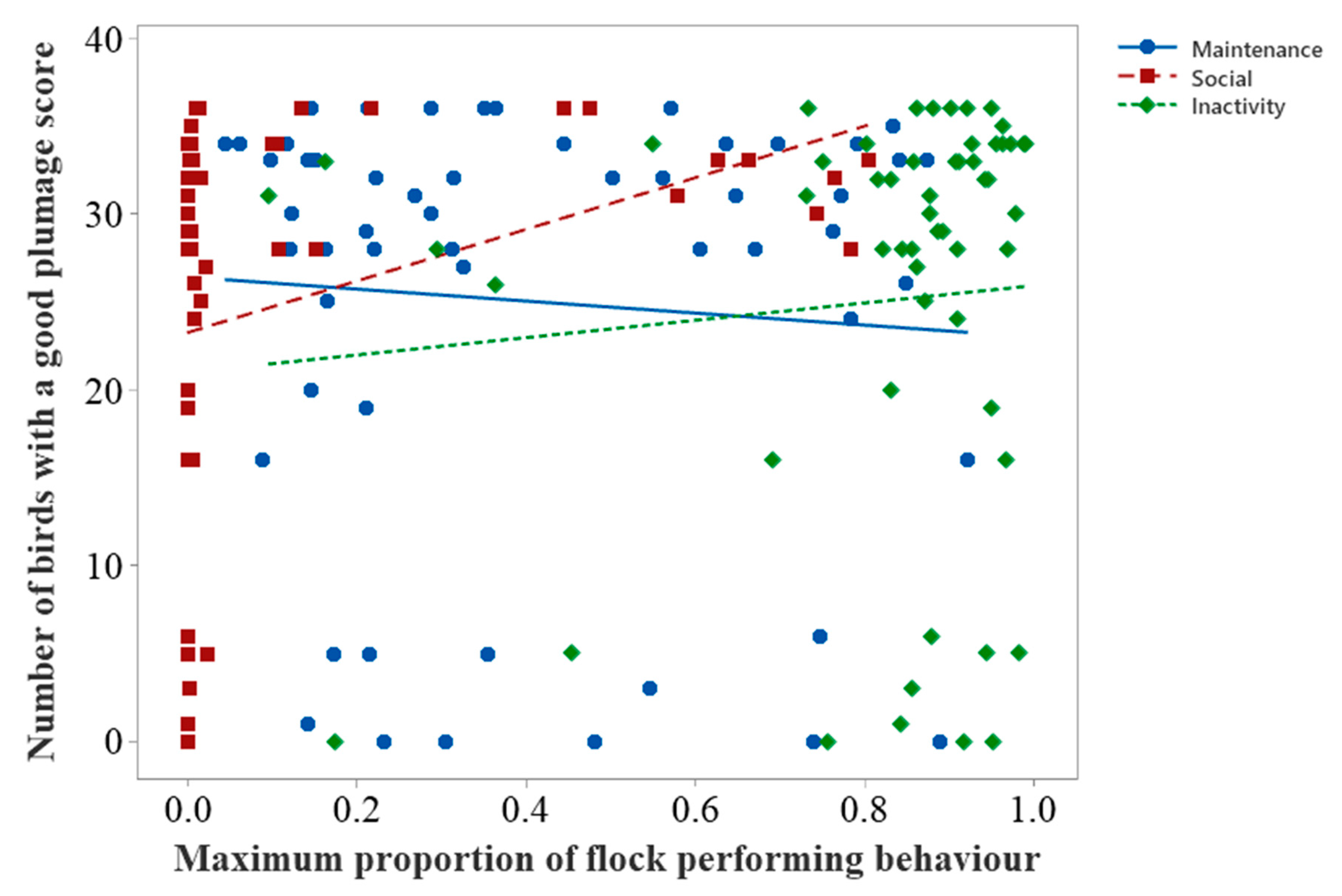
Correlation between Behaviour and Plumage Condition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jensen, T.H.; Kuiken, T. Update on Avian Influenza virus. In Fowler’s Zoo and Wild Animal Medicine: Current Therapy; Miller, R.E., Calle, P.P., Lamberski, N., Eds.; Elsevier: St Louis, MI, USA, 2023; Volume 10, pp. 139–144. [Google Scholar]
- Redrobe, S.P. Avian influenza H5N1: A review of the current situation and relevance to zoos. Int. Zoo Yearb. 2007, 41, 96–109. [Google Scholar] [CrossRef]
- Swayne, D.E. Avian Influenza. Available online: https://www.msdvetmanual.com/poultry/avian-influenza/avian-influenza (accessed on 14 December 2023).
- Swayne, D.E.; Pantin-Jackwood, M. Pathogenicity of avian influenza viruses in poultry. Dev. Biol. 2006, 124, 61–67. [Google Scholar]
- Suarez, D.L. Evolution of avian influenza viruses. Vet. Microbiol. 2000, 74, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Swayne, D.E. Impact of vaccines and vaccination on global control of avian influenza. Avian Dis. 2012, 56, 818–828. [Google Scholar] [CrossRef]
- Global Consortium for H5N8 Related Influenza Viruses. Role for migratory wild birds in the global spread of avian influenza H5N8. Science 2016, 354, 213–217. [Google Scholar] [CrossRef]
- White, K. Poultry Sector Backs Month-Long Bird Flu Prevention Zone. Available online: https://www.thegrocer.co.uk/eggs-and-poultry/poultry-sector-backs-month-long-bird-flu-prevention-zone/545830.article (accessed on 14 December 2023).
- Defra/APHA. Bird Flu (Avian Influenza): How to Prevent It and Stop It Spreading. Available online: https://www.gov.uk/guidance/bird-flu-avian-influenza-how-to-prevent-it-and-stop-it-spreading (accessed on 14 December 2023).
- Defra/APHA. Bird Flu—Latest Situation: Chief Vet Lifts Prevention Zone. Available online: https://www.gov.uk/government/news/bird-flu-latest-situation-avian-influenza-prevention-zone-declared-across-great-britain (accessed on 14 December 2023).
- East, I.J.; Roche, S.E.; Wicks, R.M.; de Witte, K.; Garner, M.G. Options for managing animal welfare on intensive pig farms confined by movement restrictions during an outbreak of foot and mouth disease. Prev. Vet. Med. 2014, 117, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Lay, D.C., Jr.; Fulton, R.M.; Hester, P.Y.; Karcher, D.M.; Kjaer, J.B.; Mench, J.A.; Mullens, B.A.; Newberry, R.C.; Nicol, C.J.; O’Sullivan, N.P. Hen welfare in different housing systems. Poult. Sci. 2011, 90, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.R. Issues of choice and control in the behaviour of a pair of captive polar bears (Ursus maritimus). Behav. Process. 2006, 73, 117–120. [Google Scholar] [CrossRef]
- Mason, G.J.; Burn, C. Behavioural restriction. In Animal Welfare; Appleby, M., Mench, J.A., Olsson, A., Hughes, B.O., Eds.; CAB International: Wallingford, UK, 2011; pp. 98–119. [Google Scholar]
- Morgan, K.N.; Tromborg, C.T. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–302. [Google Scholar] [CrossRef]
- Broom, D.M. Indicators of poor welfare. Br. Vet. J. 1986, 142, 524–526. [Google Scholar] [CrossRef]
- Jones, N.; Sherwen, S.L.; Robbins, R.; McLelland, D.J.; Whittaker, A.L. Welfare assessment tools in zoos: From theory to practice. Vet. Sci. 2022, 9, 170. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.E.; Riley, L.M. Conducting behavioural research in the zoo: A guide to ten important methods, concepts and theories. J. Zool. Bot. Gard. 2021, 2, 421–444. [Google Scholar] [CrossRef]
- Riley, L.M.; Díez-León, M.; Rose, P.E. Behavioural biology and zoo animal welfare: For the future. In The Behavioural Biology of Zoo Animals; Rose, P.E., Ed.; CRC Press: Boca Raton, FL, USA, 2022; pp. 285–292. [Google Scholar]
- Dawkins, M.S. Using behaviour to assess animal welfare. Anim. Welf. 2004, 13, S3–S7. [Google Scholar] [CrossRef]
- Rose, P.E. Identifying essential elements of good giraffe welfare—Can we use knowledge of a species’ fundamental needs to develop welfare-focussed husbandry? J. Zool. Bot. Gard. 2023, 4, 549–566. [Google Scholar] [CrossRef]
- Dawkins, M.S. Behaviour as a tool in the assessment of animal welfare. Zoology 2003, 106, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Broom, D.M.; Kirkden, R.D. Welfare, stress, behaviour and pathophysiology. Vet. Pathophysiol. 2004, 2004, 337–369. [Google Scholar]
- Mason, G.J. Stereotypies and suffering. Behav. Process. 1991, 25, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Mellor, E.; Brilot, B.; Collins, S. Abnormal repetitive behaviours in captive birds: A Tinbergian review. Appl. Anim. Behav. Sci. 2018, 198, 109–120. [Google Scholar] [CrossRef]
- Rose, P.E.; Nash, S.M.; Riley, L.M. To pace or not to pace? A review of what abnormal repetitive behaviour tells us about zoo animal management. J. Vet. Behav. 2017, 20, 11–21. [Google Scholar] [CrossRef]
- Garner, J.P. Perseveration and stereotypy-systems-level insights from clinical psychology. In Stereotypic Animal Behaviour: Fundamentals and Applications to Welfare; Mason, G., Rushen, J., Eds.; Cabi: Wallingford, UK, 2006; pp. 121–152. [Google Scholar]
- Levitt, T. Poultry Farmers Call for Birds to Be Kept Inside to Combat Bird Flu. Available online: https://www.theguardian.com/world/2022/oct/12/poultry-farmers-nationwide-housing-order-combat-bird-flu-uk (accessed on 14 December 2023).
- Van Reeth, K. Avian and swine influenza viruses: Our current understanding of the zoonotic risk. Vet. Res. 2007, 38, 243–260. [Google Scholar] [CrossRef]
- Marwell Zoo. Avian Infuenza: 2023 Update. Available online: https://www.marwell.org.uk/zoo-news/avian-infuenza-2023-update/ (accessed on 14 December 2023).
- Church, E. Paignton Zoo Still Shut with Birds in Quarantine as It Makes New Announcement. Available online: https://www.devonlive.com/news/devon-news/paignton-zoo-still-shut-birds-7569627 (accessed on 14 December 2023).
- Stallknecht, D.E. Ecology and epidemiology of avian influenza viruses in wild bird populations: Waterfowl, shorebirds, pelicans, cormorants, etc. Avian Dis. 2003, 47, 61–69. [Google Scholar]
- Shriner, S.A.; Root, J.J. A review of avian influenza A virus associations in synanthropic birds. Viruses 2020, 12, 1209. [Google Scholar] [CrossRef] [PubMed]
- Krauss, S.; Webster, R.G. Avian influenza virus surveillance and wild birds: Past and present. Avian Dis. 2010, 54, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.E.; Croft, D.P.; Lee, R. A review of captive flamingo (Phoenicopteridae) welfare: A synthesis of current knowledge and future directions. Int. Zoo Yearb. 2014, 48, 139–155. [Google Scholar] [CrossRef]
- Buschschlüter, V. Bird Flu Kills Hundreds of Flamingos in Argentina. Available online: https://www.bbc.com/news/world-latin-america-67509697 (accessed on 14 December 2023).
- Rose, P.E.; Brereton, J.E.; Gardner, L. Developing flamingo husbandry practices through workshop communication. J. Zoo Aquar. Res. 2016, 4, 115–121. [Google Scholar] [CrossRef]
- Buckles, E.L. Chapter 28—Phoenicopteriformes. In Pathology of Wildlife and Zoo Animals; Terio, K.A., McAloose, D., St Leger, J., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 687–696. [Google Scholar]
- McConnell, H.; Brereton, J.; Rice, T.; Rose, P. Do Birds of a Feather Always Flock Together? Assessing Differences in Group and Individual Zoo Enclosure Usage by Comparing Commonly Available Methods. J. Zool. Bot. Gard. 2022, 3, 71–88. [Google Scholar] [CrossRef]
- Langan, J.N.; Chinnadurai, S.K. Animal welfare and birds. In Fowler’s Zoo and Wild Animal Medicine: Current Therapy; Miller, R.E., Calle, P.P., Lamberski, N., Eds.; Elsevier: St. Louis, FL, USA, 2023; pp. 279–286. [Google Scholar]
- Amat, J.A.; Garrido, A.; Rendón-Martos, M.; Portavia, F.; Rendón, M.A. Plumage coloration in greater flamingos Phoenicopterus roseus is affected by interactions between foraging site, body condition and sex. Ardeola 2022, 69, 219–229. [Google Scholar] [CrossRef]
- Freeman, H.D.; Valuska, A.J.; Taylor, R.R.; Ferrie, G.M.; Grand, A.P.; Leighty, K.A. Plumage variation and social partner choice in the greater flamingo (Phoenicopterus roseus). Zoo Biol. 2016, 35, 409–414. [Google Scholar] [CrossRef]
- Amat, J.A.; Rendón, M.A. Flamingo coloration and its significance. In Flamingos, Behavior, Biology, and Relationship with Humans; Anderson, M.J., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2017; pp. 77–95. [Google Scholar]
- Rose, P.E.; Brereton, J.E.; Croft, D.P. Measuring welfare in captive flamingos: Activity patterns and exhibit usage in zoo-housed birds. Appl. Anim. Behav. Sci. 2018, 205, 115–125. [Google Scholar] [CrossRef]
- Rose, P.E.; Badman-King, A.; Hurn, S.; Rice, T. Visitor presence and a changing soundscape, alongside environmental parameters, can predict enclosure usage in captive flamingos. Zoo Biol. 2021, 40, 363–375. [Google Scholar] [CrossRef]
- Kidd, P.; Ford, S.; Rose, P.E. Exploring the effect of the COVID-19 zoo closure period on flamingo behaviour and enclosure use at two institutions. Birds 2022, 3, 117–137. [Google Scholar] [CrossRef]
- Wark, J.D.; Cronin, K.A.; Niemann, T.; Shender, M.A.; Horrigan, A.; Kao, A.; Ross, M.R. Monitoring the behavior and habitat use of animals to enhance welfare using the ZooMonitor app. Anim. Behav. Cogn. 2019, 6, 158–167. [Google Scholar] [CrossRef]
- Rose, P.E.; Roper, A.; Banks, S.; Giorgio, C.; Timms, M.; Vaughan, P.; Hatch, S.; Halpin, S.; Thomas, J.; O’Brien, M. Evaluation of the time-activity budgets of captive ducks (Anatidae) compared to wild counterparts. Appl. Anim. Behav. Sci. 2022, 251, 105626. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R. Available online: http://www.rstudio.com (accessed on 29 July 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Barton, K. Mu-Min-Model Interference. Available online: https://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf (accessed on 1 July 2021).
- Lenth, R.; Lenth, M.R. Package ‘lsmeans’. Am. Stat. 2018, 34, 216–221. [Google Scholar] [CrossRef]
- Minitab 21. Minitab Statistical Software; Minitab: State College, PA, USA, 2023; Available online: www.minitab.com (accessed on 29 July 2022).
- Jackman, S. pscl: Classes and methods for R developed in the Political Science Computational Laboratory, New South Wales, Australia. R package version 1.5.5.1; United States Studies Centre, University of Sydney: Sydney, Australia, 2020; Available online: https://github.com/atahk/pscl/ (accessed on 29 July 2022).
- Greening, L. Stereotypies and other abnormal behavior in welfare assessment. In Encyclopedia of Animal Behavior, 2nd ed.; Choe, J.C., Ed.; Academic Press: London, UK, 2019; pp. 141–146. [Google Scholar]
- Beauchamp, G.; McNeil, R. Vigilance in greater flamingos foraging at night. Ethology 2003, 109, 511–520. [Google Scholar] [CrossRef]
- Beauchamp, G. Nonrandom patterns of vigilance in flocks of the greater flamingo, Phoenicopterus ruber ruber. Anim. Behav. 2006, 71, 593–598. [Google Scholar] [CrossRef]
- Yosef, R. Individual distances among greater flamingos as indicators of tourism pressure. Waterbirds 2000, 23, 26–31. [Google Scholar] [CrossRef]
- Wemelsfelder, F. How animals communicate quality of life: The qualitative assessment of behaviour. Anim. Welf. 2007, 16, 25–31. [Google Scholar] [CrossRef]
- Muri, K.; Stubsjøen, S.M.; Vasdal, G.; Moe, R.O.; Granquist, E.G. Associations between qualitative behaviour assessments and measures of leg health, fear and mortality in Norwegian broiler chicken flocks. Appl. Anim. Behav. Sci. 2019, 211, 47–53. [Google Scholar] [CrossRef]
- Brereton, J.E.; Rose, P.E. Comparing the behaviour of wild and captive flamingos: An evaluation of published data on time-activity budgets. Flamingo 2019, 2, 34–49. [Google Scholar]
- Wyss, F.S.; Wolf, P.; Wenker, C.J.; Hoby, S.; Schumacher, V.; Béchet, A.; Robert, N.; Liesegang, A. Comparison of plasma vitamin A and E, copper and zinc levels in free-ranging and captive greater flamingos (P hoenicopterus roseus) and their relation to pododermatitis. J. Anim. Physiol. Anim. Nutr. 2014, 98, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Delfino, H.C.; Carlos, C.J. Intra-annual variation in activity budgets of a wild Chilean flamingo (Phoenicopterus chilensis) population in Southern Brazil. Austral Ecol. 2022, 47, 971–982. [Google Scholar] [CrossRef]
- Rose, P.E.; Lloyd, I.; Brereton, J.E.; Croft, D.P. Patterns of nocturnal activity in captive greater flamingos. Zoo Biol. 2018, 37, 290–299. [Google Scholar] [CrossRef]
- Rendón-Martos, M.; Vargas, J.M.; Rendón, M.A.; Garrido, A.; Ramírez, J.M. Nocturnal movements of breeding greater flamingos in southern Spain. Waterbirds 2000, 23, 9–19. [Google Scholar] [CrossRef]
- Tindle, R.W.; Tupiza, A.; Blomberg, S.P.; Tindle, L.E. The biology of an isolated population of the American flamingo Phoenicopterus ruber in the Galapagos Islands. Galapagos Res. J. Sci. Conserv. Galapagos Islands 2014, 68, 15–27. [Google Scholar]
- Rose, P.E.; Soole, L. What influences aggression and foraging activity in social birds? Measuring individual, group and environmental characteristics. Ethology 2020, 126, 900–913. [Google Scholar] [CrossRef]
- Stevens, E.F. Flamingo breeding: The role of group displays. Zoo Biol. 1991, 10, 53–63. [Google Scholar] [CrossRef]
- Rose, P.E.; Scales, J.S.; Brereton, J.E. Why the “visitor effect” is complicated. Unraveling individual animal, visitor number, and climatic influences on behavior, space use and interactions with keepers- a case study on captive hornbills. Front. Vet. Sci. 2020, 7, 236. [Google Scholar] [CrossRef]
- Goodenough, A.E.; McDonald, K.; Moody, K.; Wheeler, C. Are “visitor effects” overestimated? Behaviour in captive lemurs is mainly driven by co-variation with time and weather. J. Zoo Aquar. Res. 2019, 7, 59–66. [Google Scholar] [CrossRef]
- Mooney, A.; Teare, J.A.; Staerk, J.; Smeele, S.Q.; Rose, P.E.; Edell, R.H.; King, C.E.; Conrad, L.; Buckley, Y.M. Flock size and structure influence reproductive success in four species of flamingo in 540 captive populations worldwide. Zoo Biol. 2023, 42, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.D.; Parker, M.O.; Proops, L.; McBride, S.D. Risk factors for stereotypic behaviour in captive ungulates. Proc. R. Soc. B 2022, 289, 20221311. [Google Scholar] [CrossRef] [PubMed]
- Loader, A.; Rose, P.E. Age-related change in the association choices of two species of juvenile flamingos. Animals 2023, 13, 2623. [Google Scholar] [CrossRef] [PubMed]
- Mellor, D.J. Positive animal welfare states and encouraging environment-focused and animal-to-animal interactive behaviours. N. Z. Vet. J. 2015, 63, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Claydon, M.; Brereton, J.E.; Rose, P.E. Never be mute about bird welfare: Swanning around with environmental enrichment. Zoo Biol. 2023, 43, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Mooney, A.; McCall, K.; Bastow, S.; Rose, P.E. Changes in environment and management practices improve foot health in zoo-housed flamingos. Animals 2023, 13, 2483. [Google Scholar] [CrossRef]
- Rose, P.E.; Chapman, J.; Brereton, J.E.; Riley, L.M. What’s black and white and pink all over? Lesser flamingo nocturnal behaviour captured by remote cameras. J. Zool. Bot. Gard. 2022, 3, 624–640. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State Behaviour | Description |
|---|---|
| Inactive | Rest/Sleep: motionless with head tucked under wing, standing, or sitting, with eye(s) open or closed; standing: motionless, not vigilant, head is held in front of body; sitting: flamingo is sitting motionless, not vigilant. Eyes are open, legs are tucked under body and neck is not erect. |
| Maintenance | Cleaning and oiling feathers with bill or using water to wash feathers by scooping water over body with wings and/or bill. |
| Feeding and Foraging | Consumption of food from feed trough or natural filtering (pumping water through bill) in water bodies (e.g., the flamingos’ lake). |
| Social | Social: long-duration positive social association, defined as one bird following another around the enclosure. Courtship: long duration head-flagging (movement from side-to-side) or marching displays, or extended wing saluting (spreading of wings out to the bird’s sides). Nesting: manipulating substrate with beak, dragging items from one location to another to form a nest/mound. |
| Aggression | Spreading the scapular feathers to look more threatening, either whilst sitting or standing; hooking and/or jousting: extension of the neck and pointing of the bill at a nearby bird with the head swayed side to side and engaging in direct contact; fighting; birds push and shove one-another, using wings and beaks, either sitting or standing. |
| Alert | A flamingo stands, with head held high and neck erect, scanning immediate surroundings. Alert behaviour could be caused by disturbance, e.g., keeper access to the enclosure, passing vehicles or other extraneous stimuli. |
| Abnormal | Pacing (i.e., repetitive, invariant locomotion along the same path within the enclosure), flight attempt (i.e., consistent attempts at take-off to evade a potential or perceived threat, see Rose et al. [48]) and redirected foraging (i.e., mock foraging behaviour and the presence of filtering actions in the absence of a water source). |
| Behaviour | Predictor | F Value | Degrees of Freedom | r2 Value | p Value |
|---|---|---|---|---|---|
| Abnormal | Season | 1.496 | 2, 58.31 | 0.16 | 0.232 |
| Time slot | 0.991 | 5, 205.4 | 0.424 | ||
| Housing condition | 3.229 | 1, 55.82 | 0.078 | ||
| Good plumage | 0.619 | 1, 50.3 | 0.435 | ||
| Weather | 0.840 | 3, 125.02 | 0.475 | ||
| Aggression | Season | 2.042 | 2, 45.10 | 0.20 | 0.142 |
| Time slot | 1.611 | 5, 196.02 | 0.159 | ||
| Housing condition | 0.192 | 1, 43.04 | 0.663 | ||
| Good plumage | 0.401 | 1, 38.50 | 0.530 | ||
| Weather | 1.252 | 3, 105.89 | 0.294 | ||
| Alert | Season | 0.006 | 2, 234 | 0.27 | 0.994 |
| Time slot | 12.42 | 5, 234 | <0.001 * | ||
| Housing condition | 6.299 | 1, 234 | 0.013 * | ||
| Good plumage | 0.160 | 1, 234 | 0.690 | ||
| Weather | 0.703 | 3, 234 | 0.551 | ||
| Inactive | Season | 4.178 | 2, 51.94 | 0.43 | 0.021 * |
| Time slot | 12.76 | 5, 199.66 | <0.001 * | ||
| Housing condition | 3.110 | 1, 50.01 | 0.084 | ||
| Good plumage | 0.237 | 1, 45.07 | 0.629 | ||
| Weather | 2.882 | 3, 125.61 | 0.039 * | ||
| Maintenance | Season | 9.066 | 2, 51.44 | 0.28 | <0.001 * |
| Time slot | 4.735 | 5, 200.76 | <0.001 * | ||
| Housing condition | 0.352 | 1, 49.22 | 0.555 | ||
| Good plumage | 0.360 | 1, 44.19 | 0.552 | ||
| Weather | 4.706 | 3, 116.75 | 0.004 * | ||
| Social | Season | 0.096 | 2, 50.95 | 0.27 | 0.908 |
| Time slot | 1.784 | 5, 197.31 | 0.118 | ||
| Housing condition | 1.695 | 1, 49.34 | 0.199 | ||
| Good plumage | 5.035 | 1, 44.66 | 0.030 * | ||
| Weather | 0.370 | 3, 133.23 | 0.775 |
| Behaviour | Comparison | Estimate | SE | Degrees of Freedom | T Ratio | p Value |
|---|---|---|---|---|---|---|
| Alert | Time A-Time B | 0.258 | 0.047 | 200 | 5.516 | <0.001 |
| Time A-Time C | 0.323 | 0.047 | 203 | 6.887 | <0.001 | |
| Time A-Time D | 0.305 | 0.049 | 210 | 6.163 | <0.001 | |
| Time A-Time E | 0.275 | 0.047 | 211 | 5.818 | <0.001 | |
| Time A-Time F | 0.242 | 0.051 | 212 | 4.713 | <0.001 | |
| HO-R | 0.120 | 0.048 | 46.4 | 2.494 | 0.016 | |
| Inactive | Spring-Winter | 0.181 | 0.065 | 54.0 | 2.800 | 0.019 |
| Time A-Time C | −0.306 | 0.055 | 198 | −5.567 | <0.001 | |
| Time A-Time D | −0.365 | 0.058 | 203 | −6.282 | <0.001 | |
| Time A-Time E | −0.276 | 0.056 | 206 | −4.949 | <0.001 | |
| Time B-Time C | −0.236 | 0.055 | 198 | −4.275 | <0.001 | |
| Time B-Time D | −0.296 | 0.058 | 200 | −5.107 | <0.001 | |
| Time B-Time E | −0.206 | 0.056 | 204 | −3.720 | 0.004 | |
| Time D-Time F | 0.236 | 0.062 | 206 | 3.815 | 0.003 | |
| Rain-Sun | −0.135 | 0.050 | 108 | −2.682 | 0.042 | |
| Maintenance | Spring-Winter | −0.182 | 0.046 | 53.2 | −3.979 | 0.001 |
| Time A-Time B | −0.167 | 0.041 | 196 | −4.097 | 0.001 | |
| Time B-Time C | 0.136 | 0.041 | 199 | 3.293 | 0.015 | |
| Time B-Time D | 0.168 | 0.043 | 202 | 3.895 | 0.002 | |
| Time B-Time E | 0.136 | 0.041 | 205 | 3.291 | 0.015 | |
| Rain-Sun | 0.134 | 0.036 | 102 | 3.696 | 0.002 |
| Variable | Estimate | SE | Z Value | p Value |
|---|---|---|---|---|
| Spring | 0.276 | 0.078 | 3.549 | <0.001 * |
| Summer | 0.164 | 0.130 | 1.257 | 0.209 |
| Regular access | −0.106 | 0.087 | −1.207 | 0.227 |
| Average temperature | 0.021 | 0.008 | 2.804 | 0.005 * |
| Maintenance | 0.152 | 0.112 | 1.356 | 0.175 |
| Social | 0.779 | 0.119 | 6.567 | <0.001 * |
| Inactivity | 0.357 | 0.140 | 2.552 | 0.011 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collard, T.; Rose, P. Evaluating Potential Indicators of Welfare for Zoo Birds during an Avian Influenza Enforced Housing Order. J. Zool. Bot. Gard. 2024, 5, 416-433. https://doi.org/10.3390/jzbg5030028
Collard T, Rose P. Evaluating Potential Indicators of Welfare for Zoo Birds during an Avian Influenza Enforced Housing Order. Journal of Zoological and Botanical Gardens. 2024; 5(3):416-433. https://doi.org/10.3390/jzbg5030028
Chicago/Turabian StyleCollard, Thomas, and Paul Rose. 2024. "Evaluating Potential Indicators of Welfare for Zoo Birds during an Avian Influenza Enforced Housing Order" Journal of Zoological and Botanical Gardens 5, no. 3: 416-433. https://doi.org/10.3390/jzbg5030028
APA StyleCollard, T., & Rose, P. (2024). Evaluating Potential Indicators of Welfare for Zoo Birds during an Avian Influenza Enforced Housing Order. Journal of Zoological and Botanical Gardens, 5(3), 416-433. https://doi.org/10.3390/jzbg5030028