
Effects of Enclosure and Environmental Enrichment on the Behaviour of Ring-Tailed Lemurs (Lemur catta)


Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Subjects and Housing Conditions
2.3. Enrichment Items
2.4. Procedure
2.5. Observations
2.6. Analysis
3. Results
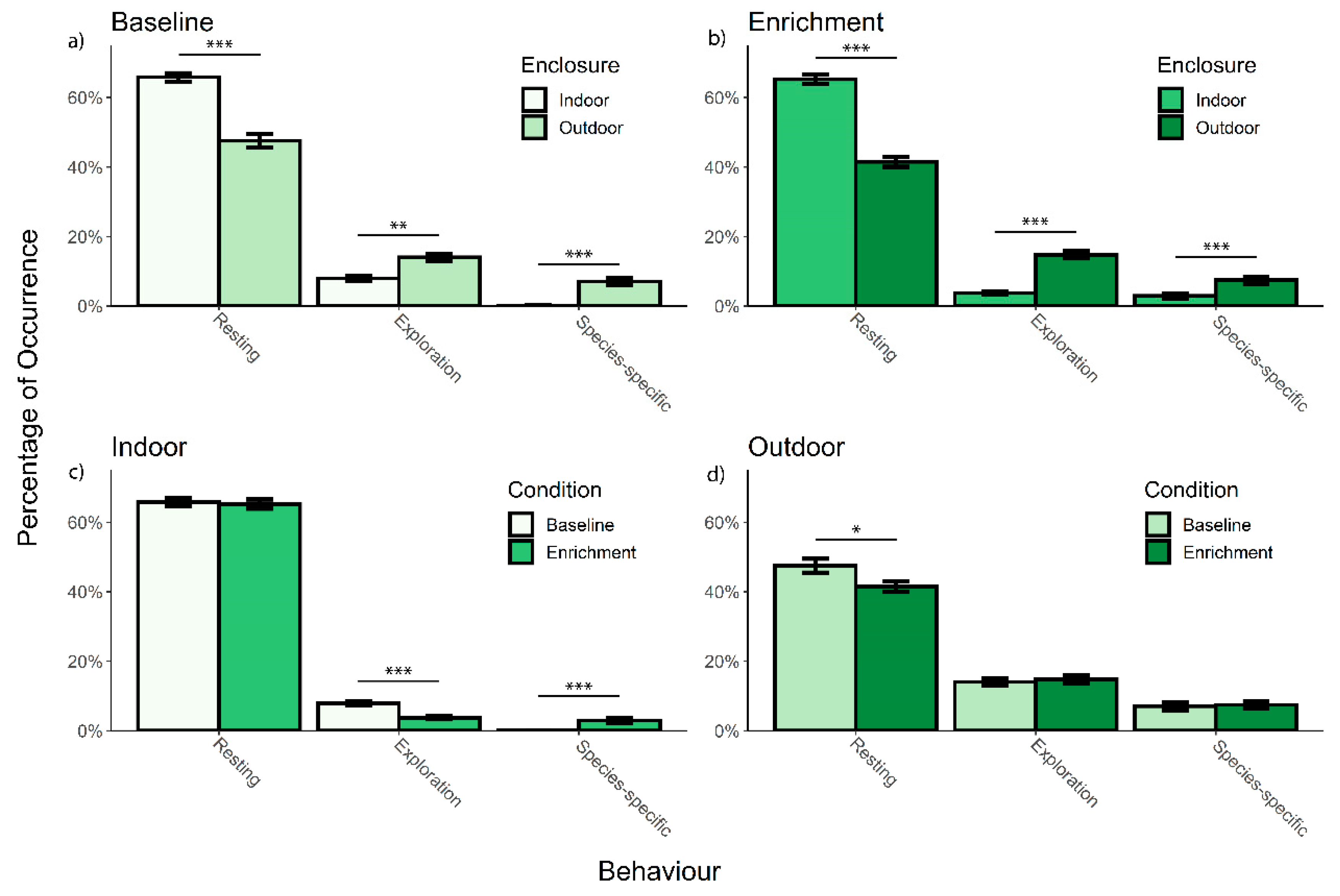
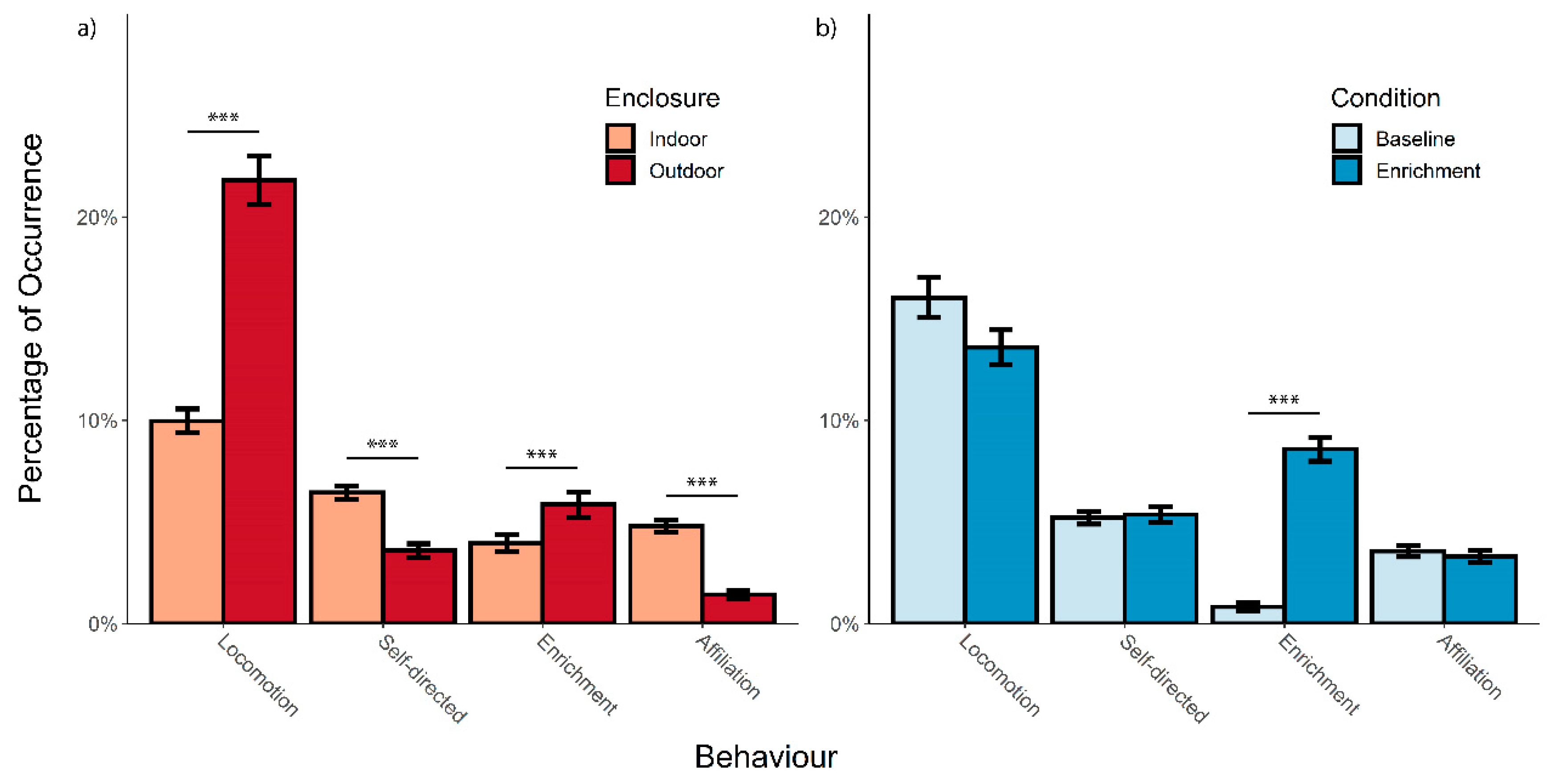
3.1. Overall Activity Budget
3.2. Effect of Enrichment and Enclosure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mellor, D.J. Updating Animal Welfare Thinking: Moving beyond the “Five Freedoms” towards “A Life Worth Living”. Animals 2016, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Buchanan-Smith, H.M. Environmental enrichment for primates in laboratories. Adv. Sci. Res. 2011, 5, 41–56. [Google Scholar] [CrossRef]
- de Azevedo, C.S.; Cipreste, C.F.; Young, R.J. Environmental enrichment: A GAP analysis. Appl. Anim. Behav. Sci. 2007, 102, 329–343. [Google Scholar] [CrossRef]
- Hoy, J.M.; Murray, P.J.; Tribe, A. Thirty years later: Enrichment practices for captive mammals. Zoo Biol. 2010, 29, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Maple, T.L.; Perdue, B.M. (Eds.) Environmental Enrichment. In Zoo Animal Welfare; Springer: Berlin/Heidelberg, Germany, 2013; pp. 95–117. ISBN 9783642359552. [Google Scholar]
- Binding, S.; Farmer, H.; Krusin, L.; Cronin, K. Status of animal welfare research in zoos and aquariums: Where are we, where to next? J. Zoo Aquar. Res. 2020, 8, 1–9. [Google Scholar]
- Mellen, J.; MacPhee, M.S. Philosophy of environmental enrichment: Past, present, and future. Zoo Biol. 2001, 20, 211–226. [Google Scholar] [CrossRef]
- Alligood, C.; Leighty, K. Putting the “E” in SPIDER: Evolving Trends in the Evaluation of Environmental Enrichment Efficacy in Zoological Settings. Anim. Behav. Cogn. 2015, 2, 200–217. [Google Scholar] [CrossRef]
- Brando, S.; Buchanan-Smith, H.M. The 24/7 approach to promoting optimal welfare for captive wild animals. Behav. Processes 2018, 156, 83–95. [Google Scholar] [CrossRef]
- Ross, S.R.; Wagner, K.E.; Schapiro, S.J.; Hau, J. Ape behavior in two alternating environments: Comparing exhibit and short-term holding areas. Am. J. Primatol. 2010, 72, 951–959. [Google Scholar] [CrossRef]
- Kurtycz, L.M.; Wagner, K.E.; Ross, S.R. The Choice to Access Outdoor Areas Affects the Behavior of Great Apes. J. Appl. Anim. Welf. Sci. 2014, 17, 185–197. [Google Scholar] [CrossRef]
- Ross, S.R.; Calcutt, S.; Schapiro, S.J.; Hau, J. Space use selectivity by chimpanzees and gorillas in an indoor-outdoor enclosure. Am. J. Primatol. 2011, 73, 197–208. [Google Scholar] [CrossRef]
- Koyama, N.F.; Aureli, F. Social network changes during space restriction in zoo chimpanzees. Primates 2019, 60, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Videan, E.N.; Fritz, J.; Schwandt, M.L.; Smith, H.F.; Howell, S. Controllability in Environmental Enrichment for Captive Chimpanzees (Pan troglodytes). J. Appl. Anim. Welf. Sci. 2005, 8, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Sha, J.C.M.; Ismail, R.; Marlena, D.; Lee, J.L. Environmental complexity and feeding enrichment can mitigate effects of space constraints in captive callitrichids. Lab. Anim. 2016, 50, 137–144. [Google Scholar] [CrossRef]
- Clubb, R.; Mason, G. Captivity effects on wide-ranging carnivores. Nature 2003, 425, 473–474. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.; Fuller, G.; Allard, S. Evaluation of the impact of behavioral opportunities on four zoo-housed aardvarks (Orycteropus afer). Animals 2020, 10, 1433. [Google Scholar] [CrossRef] [PubMed]
- Schwitzer, C.; Mittermeier, R.A.; Davies, N.; Johnson, S.; Ratsimbazafy, J.; Razafindramanana, J.; Louis, E.E., Jr.; Rajaobelina, S. Lemurs of Madagascar: A Strategy for Their Conservation 2013–2016; Bristol Conservation and Scienece Foundation: Bristol, UK, 2013. [Google Scholar]
- Maloney, M.A.; Meiers, S.T.; White, J.; Romano, M.A. Effects of three Food Enrichment Items on the behavior of black lemurs (Eulemur macaco macaco) and ringtail lemurs (Lemur catta) at the Henson Robinson Zoo, Springfield, Illinois. J. Appl. Anim. Welf. Sci. 2006, 9, 111–127. [Google Scholar] [CrossRef]
- Britt, A. Encouraging natural feeding behavior in captive-bred black and white ruffed Lemurs (Varecia variegata variegata). Zoo Biol. 1998, 17, 379–392. [Google Scholar] [CrossRef]
- Dishman, D.L.; Thomson, D.M.; Karnovsky, N.J. Does simple feeding enrichment raise activity levels of captive ring-tailed lemurs (Lemur catta)? Appl. Anim. Behav. Sci. 2009, 116, 88–95. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Timberlake, W. Selecting and Testing Environmental Enrichment in Lemurs. Front. Psychol. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Kappeler, P.M. Social status and scent-marking behaviour in Lemur catta. Anim. Behav. 1990, 40, 774–776. [Google Scholar] [CrossRef]
- Kappeler, P.M. To whom it may concern: The transmission and function of chemical signals in Lemur catta. Behav. Ecol. Sociobiol. 1998, 42, 411–421. [Google Scholar] [CrossRef]
- Baker, B.; Taylor, S.; Montrose, V.T. The effects of olfactory stimulation on the behavior of captive ring-tailed lemurs (Lemur catta). Zoo Biol. 2018, 37, 16–22. [Google Scholar] [CrossRef]
- Vaglio, S.; Kaburu, S.S.K.; Young, C.; Pearce, R.; Smith, S.; Tompkins, B.; Elwell, E.; Fontani, S.; Marliani, G.; Accorsi, P.A. Effects of scent enrichment on behavioural and physiological indicators of stress in zoo primates—A preliminary study on red-ruffed lemurs and Lar gibbons. bioRxiv 2020. [Google Scholar] [CrossRef]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef]
- Wark, J.D.; Cronin, K.A.; Niemann, T.; Shender, M.A.; Horrigan, A.; Kao, A.; Ross, M.R. Monitoring the Behavior and Habitat Use of Animals to Enhance Welfare using the ZooMonitor App. Anim. Behav. Cogn. 2019, 6, 158–167. [Google Scholar] [CrossRef]
- Ross, M.R.; Niemann, T.; Wark, J.D.; Heintz, M.R.; Horrigan, A.; Cronin, K.A.; Shender, M.A.; Gillespie, K. Zoo Monitor. Available online: https://zoomonitor.org (accessed on 16 October 2018).
- R Core Team R: A Language and Environment for Statistical Computing. 2016. Available online: https://www.rstudio.com/ (accessed on 9 February 2021).
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. Available online: https://cran.r-project.org/package=DHARMa (accessed on 9 February 2021).
- Rasamimanana, H.; Andrianome, V.N.; Rambeloarivony, H.; Pasquet, P. Male and Female Ringtailed Lemurs’ Energetic Strategy Does Not Explain Female Dominance. In Ringtailed Lemur Biology: Lemur catta in Madagascar; Jolly, A., Sussman, R.W., Koyama, N., Rasamimanana, H., Eds.; Springer: New York, NY, USA, 2006; pp. 271–295. ISBN 9780387341262. [Google Scholar]
- Keith-lucas, T.; White, F.J.; Keith-lucas, L.; Vick, L.G. Changes in Behavior in Free-Ranging Lemur catta Following Release in a Natural Habitat. Am. J. Primatol. 1999, 47, 15–28. [Google Scholar] [CrossRef]
- Browning, H. The Natural Behavior Debate: Two Conceptions of Animal Welfare. J. Appl. Anim. Welf. Sci. 2020, 23, 325–337. [Google Scholar] [CrossRef]
- Fureix, C.; Kathleen, R. What can inactivity (in its various forms) reveal about affective states in non-human animals? A review. Appl. Anim. Behav. Sci. 2015, 171, 8–24. [Google Scholar] [CrossRef]
- Honess, P.E.; Marin, C.M. Enrichment and aggression in primates. Neurosci. Biobehav. Rev. 2006, 30, 413–436. [Google Scholar] [CrossRef] [PubMed]
- Swaisgood, R.R.; Ellis, S.; Forthman, D.L.; Shepherdson, D.J.; Atlanta, Z. Commentary: Improving Well-Being for Captive Giant Pandas: Theoretical and Practical Issues. Zoo Biol. 2003, 22, 347–354. [Google Scholar] [CrossRef]
- Pines, M.K.; Kaplan, G.; Rogers, L.J. A note on indoor and outdoor housing preferences of common marmosets (Callithrix jacchus). Appl. Anim. Behav. Sci. 2007, 108, 348–353. [Google Scholar] [CrossRef]
- Goodenough, A.E.; Mcdonald, K.; Moody, K.; Wheeler, C. Are “Visitor Effects” Overestimated? Behaviour in Captive Lemurs is Mainly Driven by Co-Variation with Time and Weather. J. Zoo Aquar. Res. 2019, 7, 59–66. [Google Scholar]
- Maestripieri, D.; Schino, G.; Aureli, F.; Troisi, A. A modest proposal: Displacement activities as an indicator of emotions in primates. Anim. Behav. 1992, 44, 967–979. [Google Scholar] [CrossRef]
- Bassett, L.; Buchanan-Smith, H.M. Effects of predictability on the welfare of captive animals. Appl. Anim. Behav. Sci. 2007, 102, 223–245. [Google Scholar] [CrossRef]
- Mason, G. Stereotypies: A critical review. Anim. Behav. 1991, 41, 1015–1037. [Google Scholar] [CrossRef]
- Neal, S.J.; Caine, N.G. Scratching under positive and negative arousal in common marmosets (Callithrix jacchus). Am. J. Primatol. 2016, 78, 216–226. [Google Scholar] [CrossRef]
- Costa, R.; Sousa, C.; Llorente, M. Assessment of environmental enrichment for different primate species under low budget: A case study. J. Appl. Anim. Welf. Sci. 2018, 1–15. [Google Scholar] [CrossRef]
- Brent, L.; Belik, M. The response of group-housed baboons to three enrichment toys. Lab. Anim. 1997, 31, 81–85. [Google Scholar] [CrossRef]
- De Waal, F.B.M. The Myth of a Simple Relation Between Space and Aggression in Captive Primates. Zoo Biol. 1989, 1, 141–148. [Google Scholar] [CrossRef]
- Crast, J.; Bloomsmith, M.A.; Jonesteller, T. Effects of Changing Housing Conditions on Mangabey Behavior (Cercocebus atys): Spatial Density, Housing Quality, and Novelty Effects. Am. J. Primatol. 2015, 77, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Perdue, B.M.; Clay, A.W.; Gaalema, D.E.; Maple, T.L.; Stoinski, T.S. Technology at the Zoo: The Influence of a Touchscreen Computer on Orangutans and Zoo Visitors. Zoo Biol. 2012, 31, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Buchanan-Smith, H.M.; Badihi, I. The psychology of control: Effects of control over supplementary light on welfare of marmosets. Appl. Anim. Behav. Sci. 2012, 137, 166–174. [Google Scholar] [CrossRef]
- Brown, J.L.; Carlstead, K.; Bray, J.D.; Dickey, D.; Farin, C.; Ange-van Heugten, K. Individual and environmental risk factors associated with fecal glucocorticoid metabolite concentrations in zoo-housed Asian and African elephants. PLoS ONE 2019, 14, e0217326. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Category | Behaviour | Description |
|---|---|---|
| Abnormal | Stereotypic walking | Walking the same route repeatedly. |
| Affiliation | Grooming | Licking or combing (using teeth) another’s fur, sometimes using hands to part the fur. |
| Mutual grooming | Two individuals simultaneously groom each other. | |
| Playing | Could include non-aggressive chasing, jumping, rolling around. | |
| Sexual behaviour | Mounting, mating, etc. | |
| Agonistic | Chasing | Moving quickly towards and following another individual, causing it to flee. |
| Fighting | Any succession of hitting, biting and/or grabbing, where two or more individuals each display at least one of the behaviours. | |
| Enrichment | Manipulating enrichment item | Any form of behaviour targeted towards the enrichment items. This also includes structural enrichment items. |
| Exploration | Foraging | Actively looking for food using hands and mouth, by exploring surface or vegetation. |
| Gnawing | Chewing on an item that is not food and is not eaten. | |
| Licking | Moving tongue up and down an item that is not their or another’s body part. | |
| Sniffing | Placing the nose close to or on an object and smelling it. | |
| Locomotion | Climbing | Moving up or down from a vertical surface, using its four limbs. |
| Hanging | Hanging from an item in the environment, not touching a horizontal surface for support. | |
| Jumping | Moving from the ground or an elevated surface to the next, during which all four limbs are in the air at one point. | |
| Running | Moving quickly from one area to another by using four limbs. | |
| Standing on four legs | Using both feet and hands for support. Not any other body part is touching the ground/supporting surface (except for the tail). | |
| Standing upright | Standing on feet only, with the body upright. | |
| Walking | Moving from one area to another by using four limbs. | |
| Resting | Resting | Sitting/laying down with eyes open or closed. |
| Sitting | Placed on the base of the tail. | |
| Species-specific | Scent marking | Marking an item of the environment with a scent gland. Lifting tail and rubbing genital area/scent glands against an object. |
| Sunbathing | Sitting upright in an area with sunlight, with the belly-side directed towards the sun and arms open. | |
| Tail rubbing | Rubbing tail with the glands on forearms. | |
| Self-directed | Self-scratching | Quickly moving fingernails or toenails up and down on one of its body parts. |
| Self-grooming | Licking or combing (using teeth) own fur, sometimes using hands to part the fur. | |
| Other | Behaviours not listed above. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laméris, D.W.; Verspeek, J.; Depoortere, A.; Plessers, L.; Salas, M. Effects of Enclosure and Environmental Enrichment on the Behaviour of Ring-Tailed Lemurs (Lemur catta). J. Zool. Bot. Gard. 2021, 2, 164-173. https://doi.org/10.3390/jzbg2020012
Laméris DW, Verspeek J, Depoortere A, Plessers L, Salas M. Effects of Enclosure and Environmental Enrichment on the Behaviour of Ring-Tailed Lemurs (Lemur catta). Journal of Zoological and Botanical Gardens. 2021; 2(2):164-173. https://doi.org/10.3390/jzbg2020012
Chicago/Turabian StyleLaméris, Daan W., Jonas Verspeek, Arno Depoortere, Lise Plessers, and Marina Salas. 2021. "Effects of Enclosure and Environmental Enrichment on the Behaviour of Ring-Tailed Lemurs (Lemur catta)" Journal of Zoological and Botanical Gardens 2, no. 2: 164-173. https://doi.org/10.3390/jzbg2020012
APA StyleLaméris, D. W., Verspeek, J., Depoortere, A., Plessers, L., & Salas, M. (2021). Effects of Enclosure and Environmental Enrichment on the Behaviour of Ring-Tailed Lemurs (Lemur catta). Journal of Zoological and Botanical Gardens, 2(2), 164-173. https://doi.org/10.3390/jzbg2020012