
Unlocking the Puzzle: Investigating the Role of Interleukin 17 Genetic Polymorphisms, Circulating Lymphocytes, and Serum Levels in Venezuelan Women with Recurrent Pregnancy Loss

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Samples
2.2. Antibodies and Reagents
2.3. Genetic Polymorphism Analysis
2.4. Analysis of Intracellular IL-17 in Different Cell Subpopulations
2.5. Determination of Il-17A from Serum Samples and Supernatants
2.6. Statistical Analysis
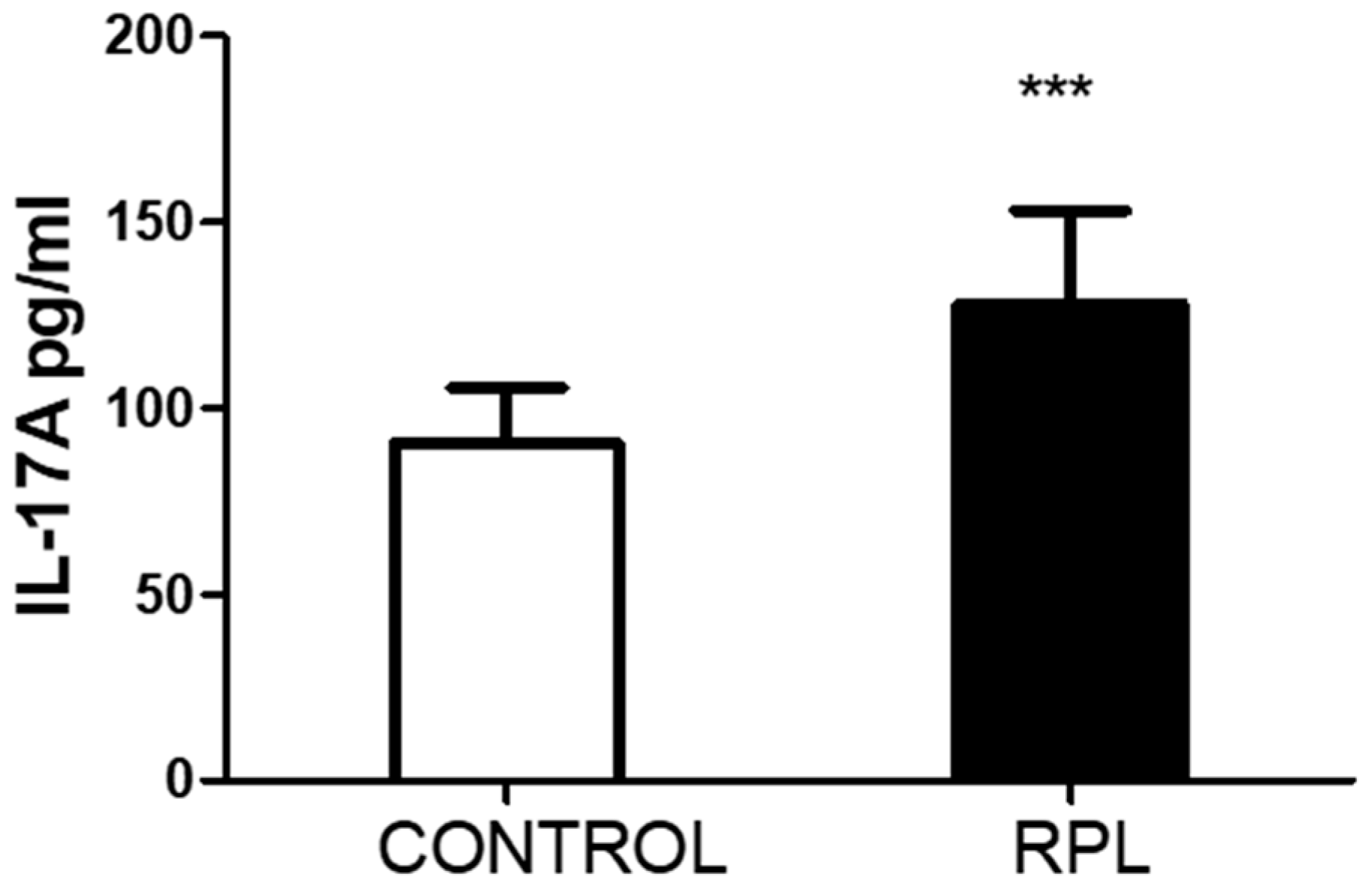
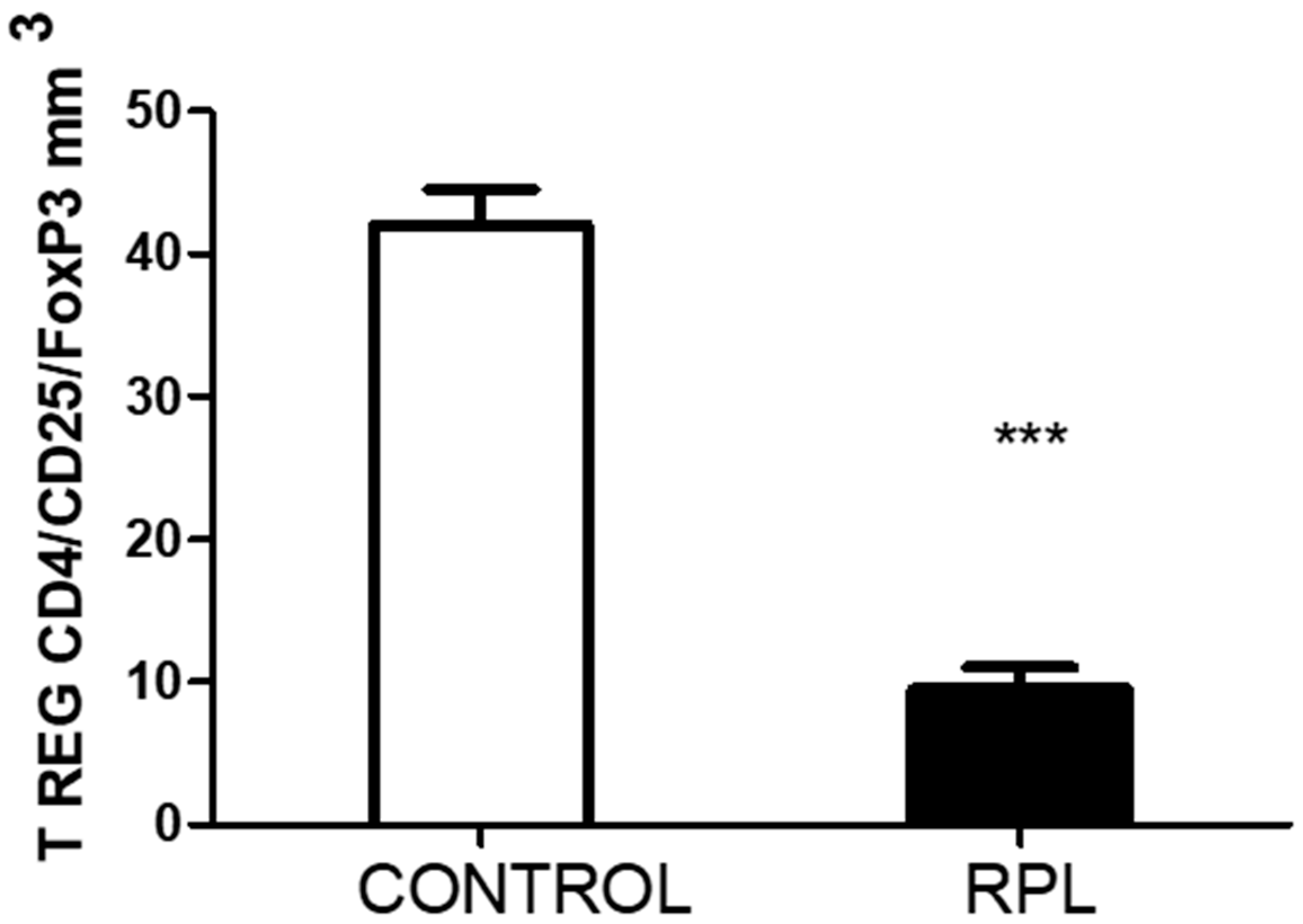
3. Results
4. Discussion
5. Conclusions
- There is no association between the two polymorphisms analyzed, IL-17A rs2275913 and IL-17F rs763780, with RPL. The values recorded from plasma of RPL patients were independent of the genetic polymorphisms.
- The peripheral lymphocytes of RPL patients were activated based on the percentage of HLA-DR expression. In addition, the number of T regulatory cells decreased.
- The number of IL-17-positive cells, CD8 and CD56, was significantly lower in RPL patients than in controls. However, IL-17 positiveness in the CD3CD4 subpopulation was higher in RPL patients than in controls.
- In stimulated cells, the response of all different cell populations and subpopulations was lower in RPL patients than in the controls. The effect of PMA/ionomycin stimulation on whole blood may be responsible for this effect.
- NK cells of RPL patients responded significantly more strongly upon PMA ionomycin stimulation than those of controls.
6. Limitations of the Study
- Other circulating cytokines were not measured in the present study. An analysis of inflammatory and anti-inflammatory cytokines may have helped us identify which Th17 cell subpopulation was circulating in these patients, as Navarron-Compán et al. [30] proposed in chronic diseases.
- Unfortunately, we could not quantify the number of CD4/CD25/FoxP3 cells in the stimulated samples, which could have been an essential point for comparing with IL-17. The significant differences recorded in the basal levels suggest that upon higher circulating levels of IL-17, the number of T regulatory cells decreases.
- Some of these RPL patients did not continue visits to the fertility clinic for other procedures due to the costs, so we could not perform a follow-up to ascertain if they had any successful pregnancies.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pillarisetty, L.S.; Mahdy, H. Recurrent Pregnancy Loss; StatPearls Publishing: Treasure Island, FL, USA, 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK554460/ (accessed on 4 June 2024).
- Stephenson, M.D. Frequency of factors associated with habitual abortion in 197 couples. Fertil. Steril. 1996, 66, 24–29. [Google Scholar] [PubMed]
- Ford, H.B.; Schust, D.J. Recurrent pregnancy loss: Etiology, diagnosis, and therapy. Rev. Obstet. Gynecol. 2009, 2, 76–83. [Google Scholar] [PubMed]
- Cuadrado-Torroglosa, I.; García-Velasco, J.A.; Alecsandru, D. Maternal-Fetal Compatibility in Recurrent Pregnancy Loss. J. Clin. Med. 2024, 13, 2379. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Nakashima, A.; Ito, M.; Shima, T. Clinical implication of recent advances in our understanding of IL-17 and reproductive immunology. Expert. Rev. Clin. Immunol. 2011, 7, 649–657. [Google Scholar] [CrossRef]
- Fu, B.; Tian, Z.; Wei, H. TH17 cells in human recurrent pregnancy loss and pre-eclampsia. Cell. Mol. Immunol. 2014, 11, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Xu, Y.; Gong, G.; Zhang, Y. Roles of immune microenvironment in the female reproductive maintenance and regulation: Novel insights into the crosstalk of immune cells. Front. Immunol. 2023, 14, 1109122. [Google Scholar] [CrossRef]
- Cui, H.; Wang, N.; Li, H.; Bian, Y.; Wen, W.; Kong, X.; Wang, F. The dynamic shifts of IL-10-producing Th17 and IL-17-producing Treg in health and disease: Crosstalk between ancient "Yin-Yang" theory and modern immunology. Cell Commun. Signal. 2024, 22, 99. [Google Scholar] [CrossRef]
- Moura, G.A.; Rocha, Y.M.; Moura, F.L.D.; Freitas, J.O.; Rodrigues, J.P.V.; Gonçalves, V.P.; Nicolete, R. Immune system cells modulation in patients with reproductive issues: A systematic review approach. JBRA Assis. Reprod. 2024, 28, 78–89. [Google Scholar] [CrossRef]
- Najafi, S.; Hadinedoushan, H.; Eslami, G.; Aflatoonian, A. Association of IL-17A and IL-17 F gene polymorphisms with recurrent pregnancy loss in Iranian women. J. Assist. Reprod. Genet. 2014, 31, 1491–1496. [Google Scholar] [CrossRef]
- Zhang, M.; Xu, J.; Bao, X.; Niu, W.; Wang, L.; Du, L.; Zhang, N.; Sun, Y. Association between Genetic Polymorphisms in Interleukin Genes and Recurrent Pregnancy Loss—A Systematic Review and Meta-Analysis. PLoS ONE 2017, 12, e0169891. [Google Scholar] [CrossRef]
- Vahid, S.A.; Ghaebi, M.; Ahmadi, M.; Nouri, M.; Danaei, S.; Aghebati-Maleki, L.; Ardehaie, R.M.; Yousefi, B.; Hakimi, P.; Hojjat-Farsangi, M.; et al. Altered T-cell subpopulations in recurrent pregnancy loss patients with cellular immune abnormalities. J. Cell. Physiol. 2019, 234, 4924–4933. [Google Scholar] [CrossRef]
- Farshchi, M.; Abdollahi, E.; Saghafi, N.; Hosseini, A.; Fallahi, S.; Rostami, S.; Rostami, P.; Rafatpanah, H.; Habibagahi, M. Evaluation of Th17 and Treg cytokines in patients with unexplained recurrent pregnancy loss. J. Clin. Transl. Res. 2022, 8, 256–265. [Google Scholar]
- Ali, S.; Majid, S.; Ali, N.; Banday, M.Z.; Taing, S.; Wani, S.; Almuqbil, M.; Alshehri, S.; Shamim, K.; Rehman, M.U. Immunogenetic Role of IL17A Polymorphism in the Pathogenesis of Recurrent Miscarriage. J. Clin. Med. 2022, 11, 7448. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, M.; Ye, S.; Liu, Y.; Zhao, X.; Wang, Y. Association of IL-17 and IL-27 polymorphisms with susceptibility to recurrent pregnancy loss and pre-eclampsia: A systematic review and meta-analysis. Immun. Inflamm. Dis. 2023, 11, e1057. [Google Scholar] [CrossRef]
- Keshavarz Motamed, A.; Zarei, Z.H.; Mirfakhraee, H.; Shariatinia, F.; Akbari, M.; Ziagham, S.; Igder, S.; Zarei, N. Association of Interleukin-17A rs2275913 Polymorphism with Recurrent Miscarriage: A Systematic Review and Meta-Analysis Study. Int. J. Fertil. Steril. 2023, 18, 7–11. [Google Scholar]
- Li, D.; Uskenbayeva, N.; Fang, L.; Xu, Y.; Yan, H.; Zhang, K.; Wang, J. Genetic polymorphism of IL-17 influences susceptibility to recurrent pregnancy loss in a Chinese population. Medicine 2024, 103, e38333. [Google Scholar] [CrossRef]
- Gao, J.F.; Zhang, H.; Lv, J.; Wang, L.; Fan, Y.Y. Associations of the IL-17A rs2275913 and IL-17F rs763780 polymorphisms with the risk of digestive system neoplasms: A meta-analysis. Int. Immunopharmacol. 2019, 67, 248–259. [Google Scholar] [CrossRef]
- Li, J.; Tian, H.; Jiang, H.J.; Han, B. Interleukin-17 SNPs and serum levels increase ulcerative colitis risk: A meta-analysis. World J. Gastroenterol. 2014, 20, 15899–15909. [Google Scholar] [CrossRef]
- Stavros, S.; Panagopoulos, P.; Machairiotis, N.; Potiris, A.; Mavrogianni, D.; Sfakianakis, A.; Drakaki, E.; Christodoulaki, C.; Panagiotopoulos, D.; Sioutis, D.; et al. Association between cytokine polymorphisms and recurrent pregnancy loss: A review of current evidence. Int. J. Gynecol. Obstet. 2024, 167, 45–57. [Google Scholar] [CrossRef]
- Conesa, A.; Fernández-Mestre, M.; Padrón, D.; Toro, F.; Silva, N.; Tassinari, P.; Blanca, I.; Martin, M.P.; Carrington, M.; Layrisse, Z. Distribution of killer cell immunoglobulin-like receptor genes in the mestizo population from Venezuela. Tissue Antigens 2010, 75, 724–729. [Google Scholar] [CrossRef]
- del Fortes, P.M.; Gill, G.; Paredes, M.E.; Gamez, L.E.; Palacios, M.; Blanca, I.; Tassinari, P. Allele and haplotype frequencies at human leukocyte antigen class I and II genes in Venezuela’s population. Ann. Biol. Clin. 2012, 70, 175–181. [Google Scholar]
- Bryc, K.; Velez, C.; Karafet, T.; Moreno-Estrada, A.; Reynolds, A.; Auton, A.; Hammer, M.; Bustamante, C.D.; Ostrer, H. Colloquium paper: Genome-wide patterns of population structure and admixture among Hispanic/Latino populations. Proc. Natl. Acad. Sci. USA 2010, 107 (Suppl. S2), 8954–8961. [Google Scholar] [CrossRef]
- De Oliveira, T.C.; Secolin, R.; Lopes-Cendes, I. A review of ancestrality and admixture in Latin America and the Caribbean focusing on native American and African descendant populations. Front. Genet. 2023, 14, 1091269. [Google Scholar] [CrossRef]
- Niafar, M.; Samaie, V.; Soltani-Zangbar, M.S.; Motavalli, R.; Dolati, S.; Danaii, S.; Mehdizadeh, A.; Yousefi, M. The association of Treg and Th17 cells development factors and anti-TPO autoantibodies in patients with recurrent pregnancy loss. BMC Res. Notes 2023, 16, 302. [Google Scholar] [CrossRef]
- Peña, M.J.; De Sanctis, C.V.; De Sanctis, J.B.; Garmendia, J.V. Frequency of Gene Polymorphisms in Admixed Venezuelan Women with Recurrent Pregnancy Loss: Microsomal Epoxy Hydroxylase (rs1051740) and Enos (rs1799983). Curr. Issues Mol. Biol. 2024, 46, 3460–3469. [Google Scholar] [CrossRef]
- Xie, Z.; Ding, X.; Wang, Y.; Zhang, M. The rs2275913 polymorphism of the interleukin-17A gene is associated with the risk of ovarian endometriosis. J. Obstet. Gynaecol. 2023, 43, 2199852. [Google Scholar] [CrossRef]
- Li, H.; Zhou, Z.; Tai, W.; Feng, W.; Zhang, D.; Gu, X.; Yang, R. Decreased Frequency of IL-17F rs763780 Site Allele G is Associated With Genetic Susceptibility to Immune Thrombocytopenia in a Chinese Population. Clin. Appl. Thromb. Hemost. 2017, 23, 466–471. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, J.Y.; Hur, S.E.; Kim, C.J.; Na, B.J.; Lee, M.; Gilman-Sachs, A.; Kwak-Kim, J. An imbalance in interleukin-17-producing T and Foxp3⁺ regulatory T cells in women with idiopathic recurrent pregnancy loss. Hum. Reprod. 2011, 26, 2964–2971. [Google Scholar] [CrossRef]
- Navarro-Compán, V.; Puig, L.; Vidal, S.; Ramírez, J.; Llamas-Velasco, M.; Fernández-Carballido, C.; Almodóvar, R.; Pinto, J.A.; Galíndez-Aguirregoikoa, E.; Zarco, P.; et al. The paradigm of IL-23-independent production of IL-17F and IL-17A and their role in chronic inflammatory diseases. Frontier. Immunol. 2023, 14, 1191782. [Google Scholar] [CrossRef]
- Mills, K.H. Induction, function and regulation of IL-17-producing T cells. Eur. J. Immunol. 2008, 38, 2636–2649. [Google Scholar] [CrossRef]
- Purvis, H.A.; Stoop, J.N.; Mann, J.; Woods, S.; Kozijn, A.E.; Hambleton, S.; Robinson, J.H.; Isaacs, J.D.; Anderson, A.E.; Hilkens, C.M. Low-strength T-cell activation promotes Th17 responses. Blood 2010, 116, 4829–4837. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Zhou, Y.; Yu, J.; Mao, L.; Bosco, M.J.; Wang, J.; Lu, Y.; Mao, L.; Wu, X.; Wang, F.; et al. Establishment of the Reference Intervals of Lymphocyte Function in Healthy Adults Based on IFN-γ Secretion Assay upon Phorbol-12-Myristate-13-Acetate/Ionomycin Stimulation. Front. Immunol. 2018, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Pandya, A.D.; Al-Jaderi, Z.; Høglund, R.A.; Holmøy, T.; Harbo, H.F.; Norgauer, J.; Maghazachi, A.A. Identification of human NK17/NK1 cells. PLoS ONE 2011, 6, e26780. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Controls | RPL | |
|---|---|---|
| n | 50 | 50 |
| Age (years) | 34.3 ± 6.5 | 34.1 ± 4.5 |
| # Pregnancies (%) | 1 (10%) | 2 (50%) |
| 2 (60%) | 3 (35%) | |
| 3 (30%) | >3 (15%) | |
| # Miscarriages (%) | 0 | 2 (40%) |
| >2 (60%) | ||
| Duration of pregnancy (weeks) | 37.3 ± 2.2 | 8.1 ± 2.5 |
| Polymorphism | Control | RPL | p | OR |
|---|---|---|---|---|
| rs2275913 | ||||
| Genotype | ||||
| GG | 40 | 41 | 0.9 | 1.0 |
| GA | 8 | 8 | 0.9 | |
| AA | 2 | 1 | ||
| G | 48 | 50 | 0.8 | 1.0 |
| A | 10 | 9 | 0.8 | |
| rs763780 | ||||
| Genotype | ||||
| AA | 47 | 47 | 0.9 | 1.0 |
| GA | 1 | 2 | 0.9 | |
| GG | 2 | 1 | ||
| A | 48 | 50 | 0.9 | 1.0 |
| G | 3 | 3 | 0.9 |
| Control | RPL | p | |
|---|---|---|---|
| Total leukocytes | 6700 ± 814 | 6766 ± 727 | 0.7 |
| Total lymphocytes | 2105 ± 241 | 2359 ± 239 | >0.001 |
| CD3 T lymphocytes | 1530 ± 208 | 1614 ± 186 | 0.03 |
| NK cells (CD56/CD16) | 186 ± 37 | 261 ± 36 | >0.001 |
| NKT cells (CD3/CD56) | 34 ± 11 | 35 ± 14 | 0.7 |
| Index T cell/NK cell | 8.5 ± 1.6 | 6.2 ± 0.8 | >0.001 |
| CD3/CD45RA | 839 ± 134 | 1266 ± 165 | >0.001 |
| CD3/CD45RO | 928 ± 123 | 1076 ± 151 | >0.001 |
| HLA DR+ cells | 153 ± 43 | 247 ± 49 | >0.001 |
| CD3/HLA DR+ | 22 ± 8 | 82 ± 52 | >0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garmendia, J.V.; Blanca, I.; Peña, M.J.; De Sanctis, C.V.; De Sanctis, J.B. Unlocking the Puzzle: Investigating the Role of Interleukin 17 Genetic Polymorphisms, Circulating Lymphocytes, and Serum Levels in Venezuelan Women with Recurrent Pregnancy Loss. Immuno 2024, 4, 301-311. https://doi.org/10.3390/immuno4040019
Garmendia JV, Blanca I, Peña MJ, De Sanctis CV, De Sanctis JB. Unlocking the Puzzle: Investigating the Role of Interleukin 17 Genetic Polymorphisms, Circulating Lymphocytes, and Serum Levels in Venezuelan Women with Recurrent Pregnancy Loss. Immuno. 2024; 4(4):301-311. https://doi.org/10.3390/immuno4040019
Chicago/Turabian StyleGarmendia, Jenny Valentina, Isaac Blanca, María Johanna Peña, Claudia Valentina De Sanctis, and Juan Bautista De Sanctis. 2024. "Unlocking the Puzzle: Investigating the Role of Interleukin 17 Genetic Polymorphisms, Circulating Lymphocytes, and Serum Levels in Venezuelan Women with Recurrent Pregnancy Loss" Immuno 4, no. 4: 301-311. https://doi.org/10.3390/immuno4040019
APA StyleGarmendia, J. V., Blanca, I., Peña, M. J., De Sanctis, C. V., & De Sanctis, J. B. (2024). Unlocking the Puzzle: Investigating the Role of Interleukin 17 Genetic Polymorphisms, Circulating Lymphocytes, and Serum Levels in Venezuelan Women with Recurrent Pregnancy Loss. Immuno, 4(4), 301-311. https://doi.org/10.3390/immuno4040019