
A Proposal for a Hybrid Model Based on the Weibull Growth Equation in the Adjustment of Growth Curves applied to Pine Forest Species in Northern Mexico †



 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
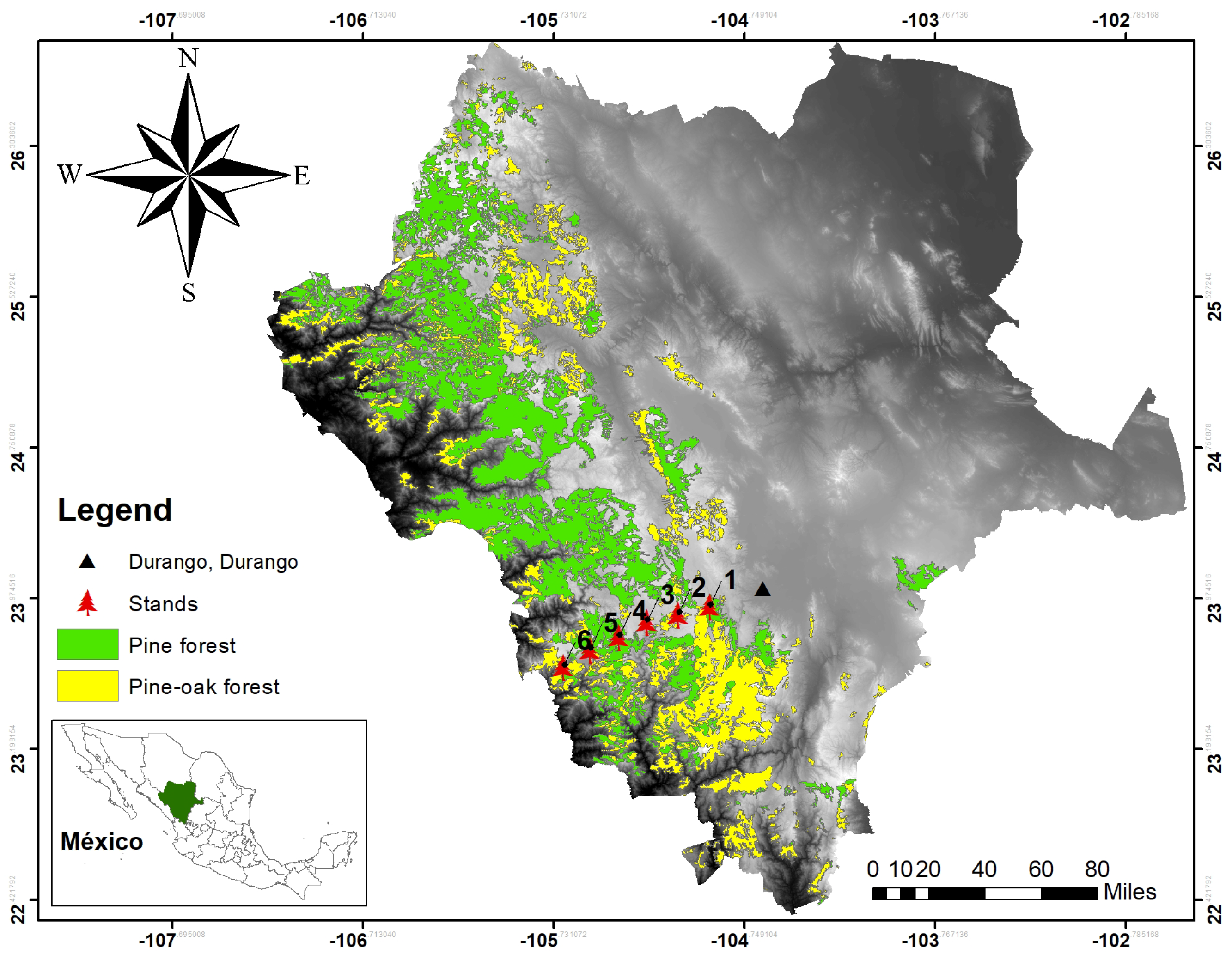
2.1. Study Area
2.2. Base-Aged-Specific Model (BAS Model)
2.3. Algebraic Difference Approach (ADA) and Generalized Algebraic Difference Approach (GADA)
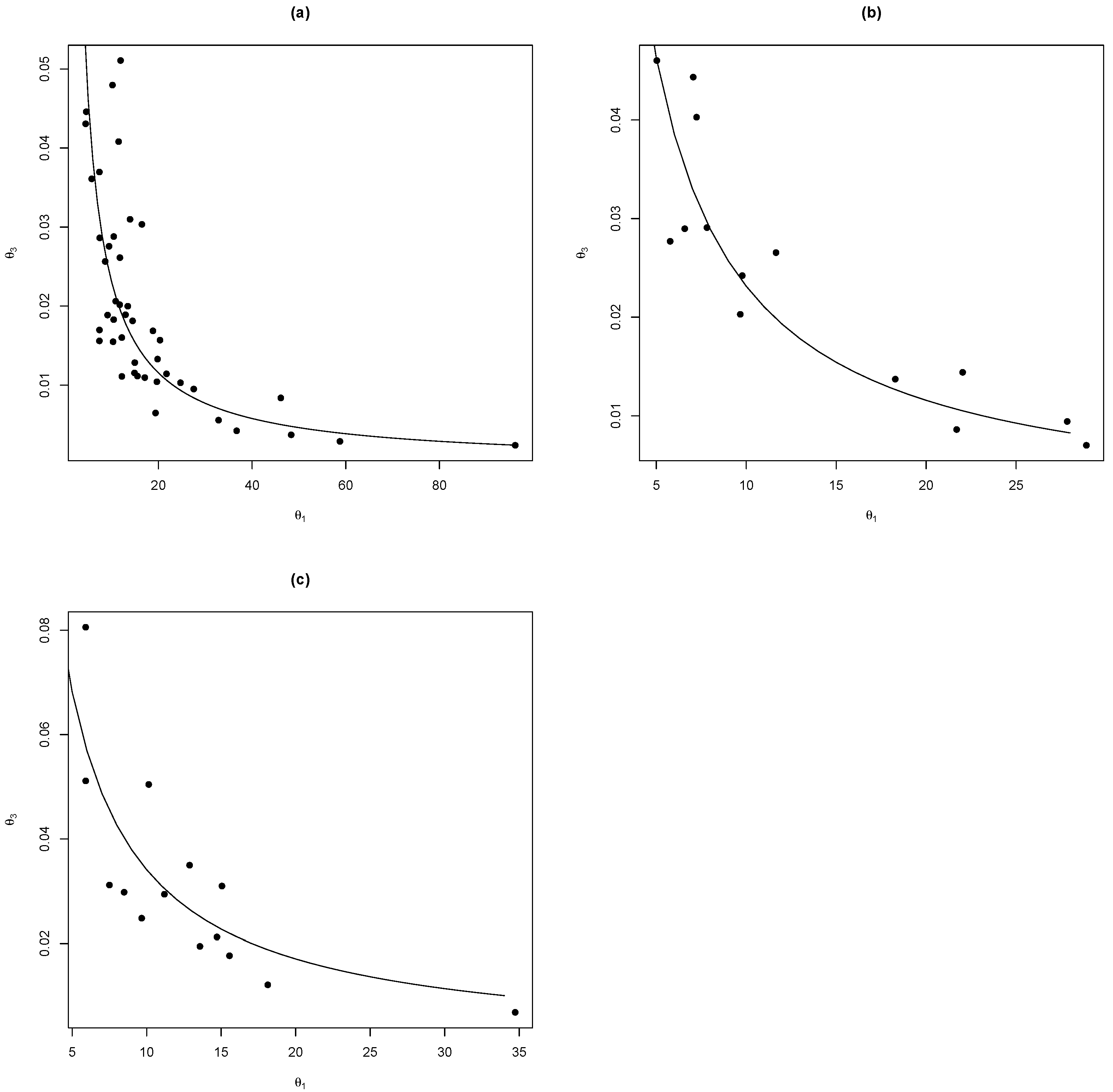
2.3.1. Chapman–Richards (Hereinafter C-R) GADA Model
2.3.2. Hossfeld GADA Model
2.4. Proposed Methodology Based on the Algebraic Difference Approach (ADA)
2.5. Mean Absolute Percentage Error (MAPE)
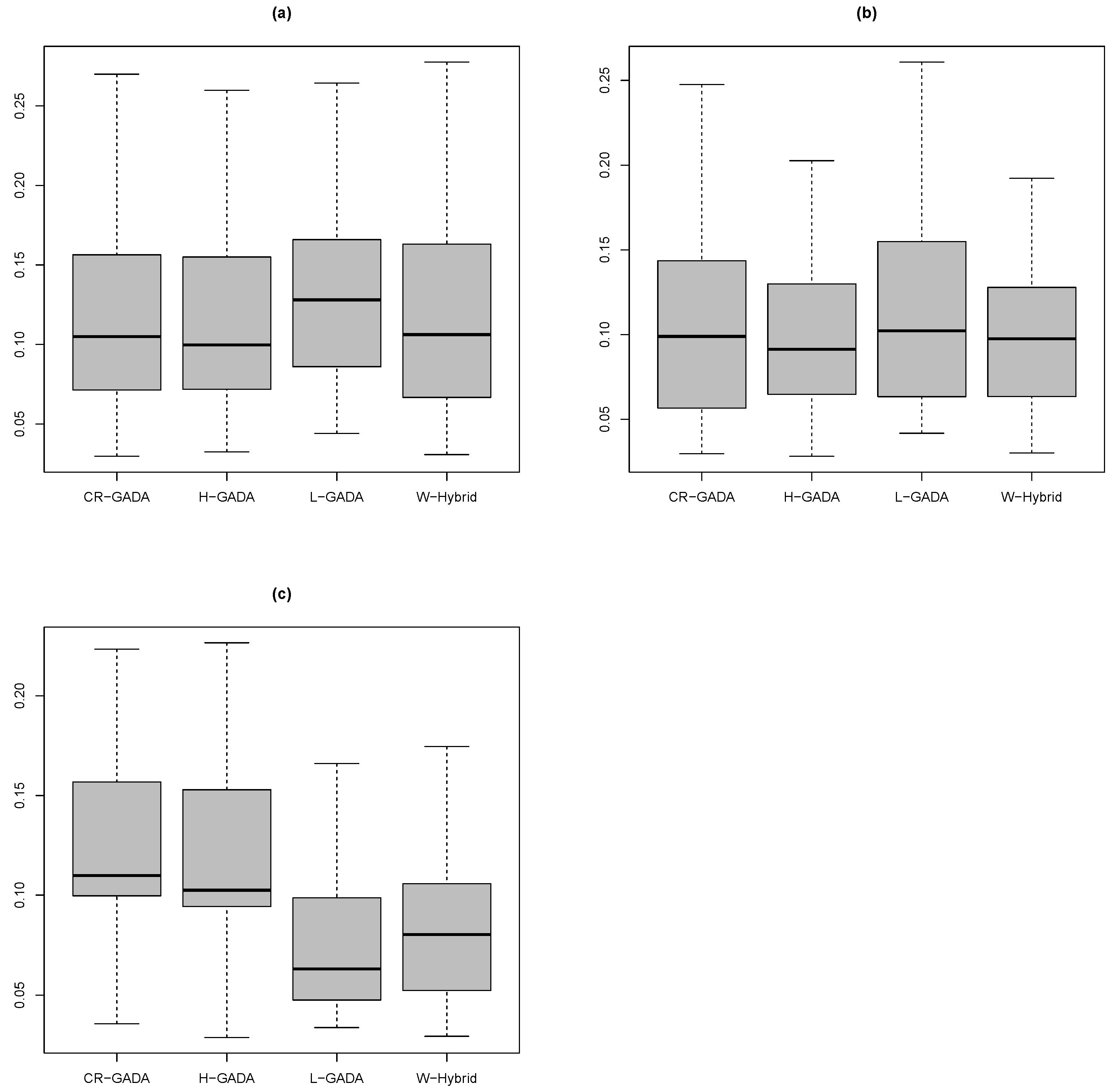
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanclay, J.K. Modelling Forest Growth and Yield: Applications to Mixed Tropical Forests; CAB International: Wallingford, UK, 1994; p. 537. [Google Scholar]
- Mitchell, K.J. Sylver: Modelling the impact of silviculture on yield, lumber value, and economic return. For. Chron. 1988, 64, 127–131. [Google Scholar] [CrossRef]
- Richards, F.J. A flexible growth function for empirical use. J. Exp. Botany 1959, 10, 290–300. [Google Scholar] [CrossRef]
- Hossfeld, J.W. Mathematik für Forstmänner, Ökonomen und Cameralisten. Gotha 1822, 4, 310. [Google Scholar]
- Weibull, W. A Statistical Distribution Function of Wide Applicability. J. Appl. Mech. 1951, 18, 293–297. [Google Scholar] [CrossRef]
- Cieszewski, C.J. Comparing Fixed- and Variable-Base-Age Site Equations Having Single Versus Multiple Asymptotes. For. Sci. 2002, 48, 7–23. [Google Scholar]
- Tofallis, C. A Better Measure of Relative Prediction Accuracy for Model Selection and Model Estimation. J. Oper. Res. Soc. 2015, 66, 1352–1362. [Google Scholar] [CrossRef]
- Bailey, R.L.; Clutter, J.L. Base-Age Invariant Polymorphic Site Curves. For. Sci. 1974, 20, 155–159. [Google Scholar]
- Bailey, R.L.; Clutter, J.L. Modeling and Prediction of Forest Growth Variables based on Multilevel nonlinear Mixed Models. For. Sci. 2001, 47, 311–321. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BAS Model | Function | Reference |
|---|---|---|
| Chapman–Richards | [3] | |
| Hossfeld | [4] | |
| Weibull | [5] |
| Model | C-R GADA | Hossfeld GADA | ||||
|---|---|---|---|---|---|---|
| Species | ||||||
| Pinus engelmannii | 0.003 | 0.843 | 0.236 | 513.109 | 0.032 | 0.882 |
| Pinus strobiformis | 0.009 | 1.019 | −0.124 | 38.051 | −0.325 | 0.983 |
| Pinus arizonica | 0.004 | 0.011 | 2.681 | 68.413 | 1.639 | 0.794 |
| Model | C-R GADA | Hossfeld GADA | Hybrid Weibull | |||
|---|---|---|---|---|---|---|
| Specie | ||||||
| Pinus engelmannii | 0.110 | 0.062 | 0.112 | 0.064 | 0.107 | 0.055 |
| Pinus strobiformis | 0.093 | 0.046 | 0.094 | 0.047 | 0.098 | 0.051 |
| Pinus arizonica | 0.113 | 0.050 | 0.107 | 0.046 | 0.084 | 0.038 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, M.d.A.; Hernández-Velazco, M.R.; Brazão-Protázio, J.M.; Escobar-Flores, J.G.; López-Sánchez, C.A.; Hernández-Díaz, J.C.; Wehenkel, C. A Proposal for a Hybrid Model Based on the Weibull Growth Equation in the Adjustment of Growth Curves applied to Pine Forest Species in Northern Mexico. Environ. Sci. Proc. 2021, 3, 107. https://doi.org/10.3390/IECF2020-08036
Souza MdA, Hernández-Velazco MR, Brazão-Protázio JM, Escobar-Flores JG, López-Sánchez CA, Hernández-Díaz JC, Wehenkel C. A Proposal for a Hybrid Model Based on the Weibull Growth Equation in the Adjustment of Growth Curves applied to Pine Forest Species in Northern Mexico. Environmental Sciences Proceedings. 2021; 3(1):107. https://doi.org/10.3390/IECF2020-08036
Chicago/Turabian StyleSouza, Marcos de Almeida, Miguel Ricardo Hernández-Velazco, João Marcelo Brazão-Protázio, Jonathan Gabriel Escobar-Flores, Carlos Antonio López-Sánchez, Jose Ciro Hernández-Díaz, and Christian Wehenkel. 2021. "A Proposal for a Hybrid Model Based on the Weibull Growth Equation in the Adjustment of Growth Curves applied to Pine Forest Species in Northern Mexico" Environmental Sciences Proceedings 3, no. 1: 107. https://doi.org/10.3390/IECF2020-08036
APA StyleSouza, M. d. A., Hernández-Velazco, M. R., Brazão-Protázio, J. M., Escobar-Flores, J. G., López-Sánchez, C. A., Hernández-Díaz, J. C., & Wehenkel, C. (2021). A Proposal for a Hybrid Model Based on the Weibull Growth Equation in the Adjustment of Growth Curves applied to Pine Forest Species in Northern Mexico. Environmental Sciences Proceedings, 3(1), 107. https://doi.org/10.3390/IECF2020-08036