
Autochthonous Biostimulants as a Promising Biological Tool to Promote Lettuce Growth and Development under Salinity Conditions †

 ,
,  and
and 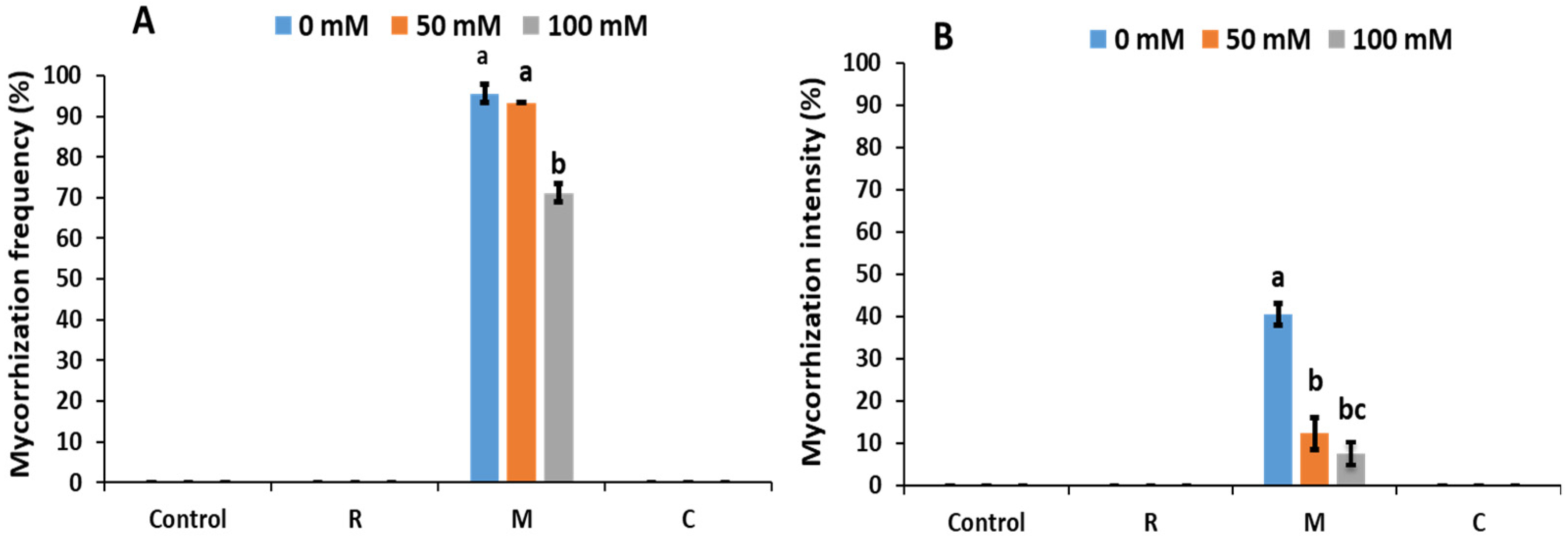{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Materials and Cultivation Conditions
2.2. Treatments and Study Design
2.3. Mycorrhization Parameters
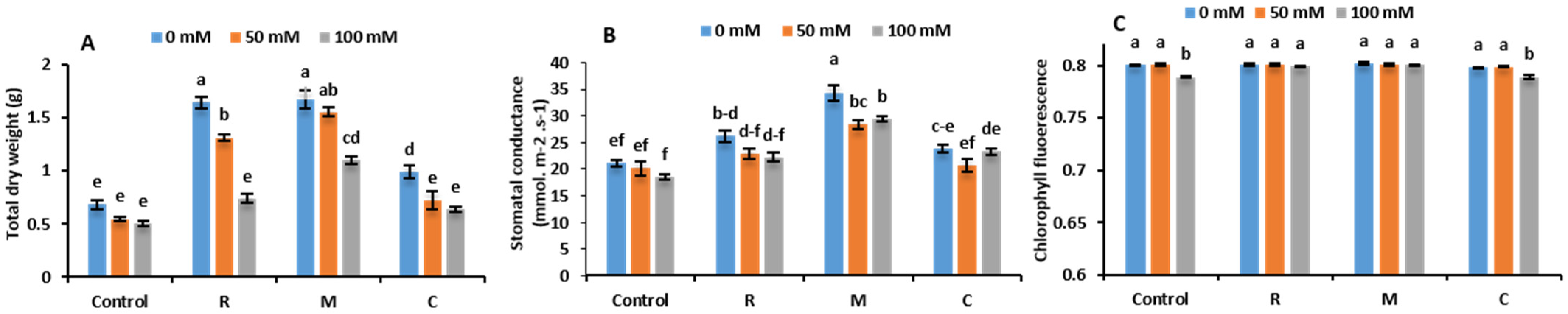
2.4. Growth Assessment
2.5. Photosynthetic Efficiency and Gas Exchanges
3. Results
3.1. Mycorrhizal Symbiosis
3.2. Growth Assessment, Photosynthetic Efficiency and Gas Exchanges
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meddich, A.; Jaiti, F.; Bourzik, W.; El Asli, A.; Hafidi, M. Use of mycorrhizal fungi as a strategy for improving the drought tolerance in date palm (Phoenix dactylifera). Sci. Hortic. 2015, 192, 468–474. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthodes d’estimation ayant une signification fonctionnelle. Mycorhizes Physiol. Génétique 1986, 217–220. [Google Scholar] [CrossRef]
- Santander, C.; Aroca, R.; Cartes, P.; Vidal, G.; Cornejo, P. Aquaporins and cation transporters are differentially regulated by two arbuscular mycorrhizal fungi strains in lettuce cultivars growing under salinity conditions. Plant Physiol. Biochem. 2021, 158, 396–409. [Google Scholar] [CrossRef]
- Khalloufi, M.; Martínez-Andújar, C.; Lachaâl, M.; Karray-Bouraoui, N.; Pérez-Alfocea, F.; Albacete, A. The interaction between foliar GA3 application and arbuscular mycorrhizal fungi inoculation improves growth in salinized tomato (Solanum lycopersicum L.) plants by modifying the hormonal balance. J. Plant Physiol. 2017, 214, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Ben-Laouane, R.; Baslam, M.; Ait-El-mokhtar, M.; Anli, M.; Boutasknit, A.; Ait-Rahou, Y.; Toubali, S.; Mitsui, T.; Oufdou, K.; Wahbi, S.; et al. Potential of native arbuscular mycorrhizal fungi, rhizobia, and/or green compost as alfalfa (Medicago sativa) enhancers under salinity. Microorganisms 2020, 8, 1695. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouhaddou, R.; Ben-Laouane, R.; Slimani, A.; Boutasknit, A.; Anli, M.; Oufdou, K.; Baslam, M.; Meddich, A. Autochthonous Biostimulants as a Promising Biological Tool to Promote Lettuce Growth and Development under Salinity Conditions. Environ. Sci. Proc. 2022, 16, 41. https://doi.org/10.3390/environsciproc2022016041
Ouhaddou R, Ben-Laouane R, Slimani A, Boutasknit A, Anli M, Oufdou K, Baslam M, Meddich A. Autochthonous Biostimulants as a Promising Biological Tool to Promote Lettuce Growth and Development under Salinity Conditions. Environmental Sciences Proceedings. 2022; 16(1):41. https://doi.org/10.3390/environsciproc2022016041
Chicago/Turabian StyleOuhaddou, Redouane, Raja Ben-Laouane, Aiman Slimani, Abderrahim Boutasknit, Mohamed Anli, Khalid Oufdou, Marouane Baslam, and Abdelilah Meddich. 2022. "Autochthonous Biostimulants as a Promising Biological Tool to Promote Lettuce Growth and Development under Salinity Conditions" Environmental Sciences Proceedings 16, no. 1: 41. https://doi.org/10.3390/environsciproc2022016041
APA StyleOuhaddou, R., Ben-Laouane, R., Slimani, A., Boutasknit, A., Anli, M., Oufdou, K., Baslam, M., & Meddich, A. (2022). Autochthonous Biostimulants as a Promising Biological Tool to Promote Lettuce Growth and Development under Salinity Conditions. Environmental Sciences Proceedings, 16(1), 41. https://doi.org/10.3390/environsciproc2022016041