
Characterization of Nitrogen Use by Neotropical Myrtaceae in Dry and Wet Forests of Southeast Brazil

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Material and Methods
2.1. Study Areas
2.2. Species Studied
2.3. Soil Sampling and Analysis
2.4. Collection of Plant Material and Analysis
2.4.1. In Vivo Activity of Nitrate Reductase (NADH) and Xylem Sap Collection
2.4.2. Sample Preparation and Nitrate (NO3−) Determination
2.4.3. Natural Abundance (δ15N Atom%), Ntotal (N%), and Ctotal (C%)
2.4.4. Free Amino Acids and N Transport
2.5. Statistical Tests
3. Results
3.1. Soil Chemical Properties
3.2. Soil Nitrogen
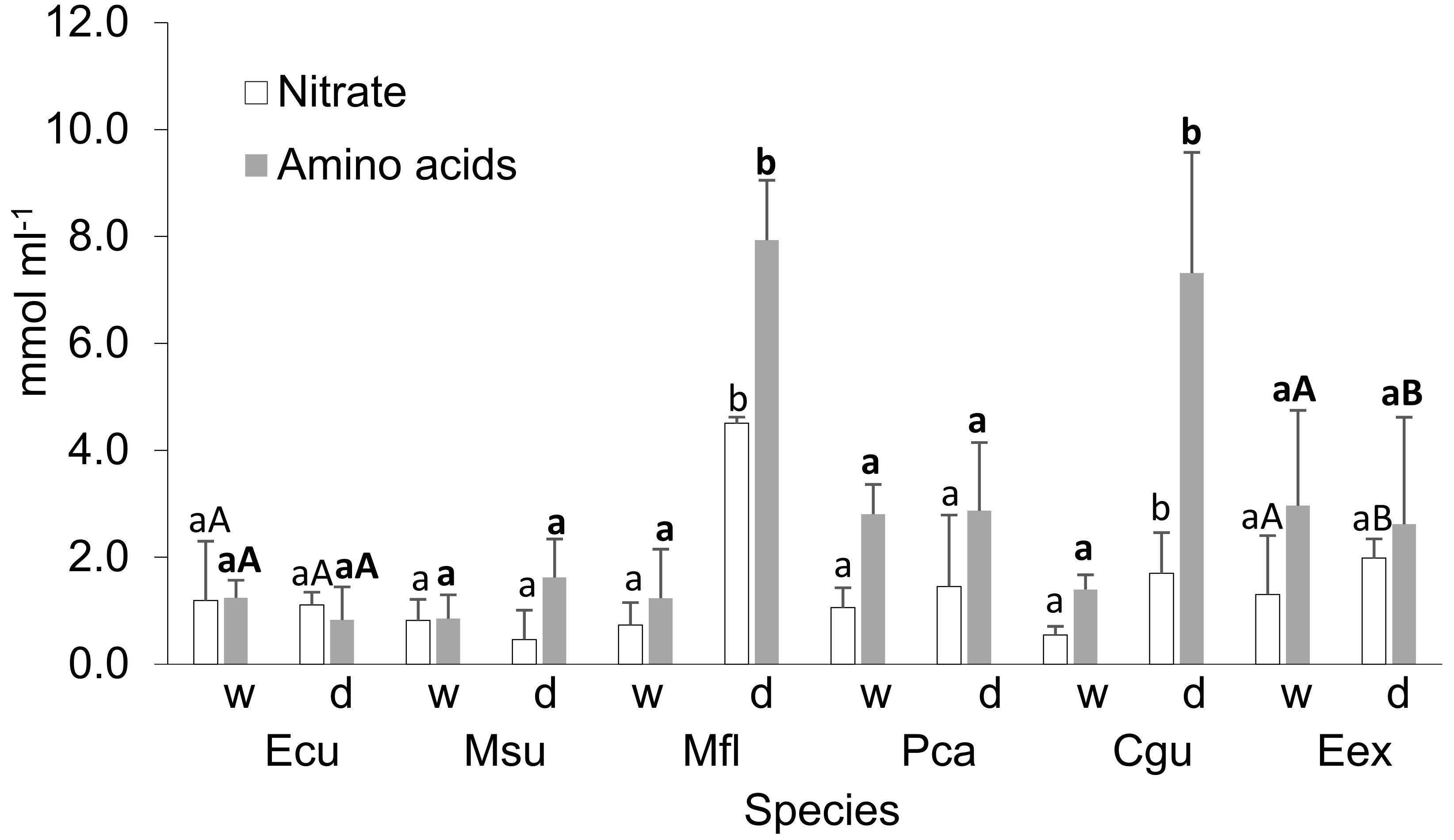
3.3. Plant Nitrogen
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Site | Forest Type | Coord. | Area | Altitude | Prec. | Etp | T | Season | Soil | |
|---|---|---|---|---|---|---|---|---|---|---|
| ha | m | mm | mm/year | °C | dry | wet | ||||
| JES | SW | 21°36′ S 47°47′ W | 9075 | 520–642 | 1470 | ~1320 | 14.2–32.5 | May–Oct | Nov–Apr | Dark Red Latosol (Oxisol) |
| SPCB | OD | 24°15′ S 47°98′ W | 37794 | 30–1003 | 1700–2400 | ~1400 | 17.0–31.7 | Apr–Sept | Oct–Mar | Haplic Cambisol (Inceptisol) |
| BRSG | SF | 22°44′ S 47°06′ W | 252 | 580–610 | 100–1360 | ~1241 | 15.5–30 | Apr–Sept | Oct–Mar | Red Argisol (Ultisol) |
References
- Fujii, K.; Shibata, M.; Kitajima, K.; Ichie, T.; Kitayama, K.; Turner, B.L. Plant–Soil Interactions Maintain Biodiversity and Functions of Tropical Forest Ecosystems. Ecol. Res. 2018, 33, 149–160. [Google Scholar] [CrossRef]
- Heineman, K.D.; Turner, B.L.; Dalling, J.W. Variation in Wood Nutrients along a Tropical Soil Fertility Gradient. New Phytol. 2016, 211, 440–454. [Google Scholar] [CrossRef] [PubMed]
- Ushio, M.; Aiba, S.; Takeuchi, Y.; Iida, Y.; Matsuoka, S.; Repin, R.; Kitayama, K. Plant–Soil Feedbacks and the Dominance of Conifers in a Tropical Montane Forest in Borneo. Ecol. Monogr. 2017, 87, 105–129. [Google Scholar] [CrossRef]
- Ueda, Y.; Konishi, M.; Yanagisawa, S. Molecular Basis of the Nitrogen Response in Plants. Soil Sci. Plant Nutr. 2017, 63, 329–341. [Google Scholar] [CrossRef]
- Hachiya, T.; Sakakibara, H. Interactions between Nitrate and Ammonium in Their Uptake, Allocation, Assimilation, and Signaling in Plants. J. Exp. Bot. 2016, 68, 2501–2512. [Google Scholar] [CrossRef]
- Cui, J.; Yu, C.; Qiao, N.; Xu, X.; Tian, Y.; Ouyang, H. Plant Preference for NH4+ versus NO3− at Different Growth Stages in an Alpine Agroecosystem. Field Crop. Res. 2017, 201, 192–199. [Google Scholar] [CrossRef]
- Rajsz, A.; Wojtuń, B.; Mróz, L.; Żołnierz, L.; Kempers, A.J. Nitrate Reductase Activity in High-Mountain Plants: A Test across Species, Growth Form and Habitat Type. J. Plant Ecol. 2018, 12, 519–530. [Google Scholar] [CrossRef]
- Tilman, D.; Wedin, D. Dynamics of Nitrogen Competition Between Successional Grasses. Ecology 1991, 72, 1038–1049. [Google Scholar] [CrossRef]
- Cantón, F.R.; Suárez, M.F.; Cánovas, F.M. Molecular Aspects of Nitrogen Mobilization and Recycling in Trees. Photosynth. Res. 2005, 83, 265–278. [Google Scholar] [CrossRef]
- Sudduth, E.B.; Perakis, S.S.; Bernhardt, E.S. Nitrate in Watersheds: Straight from Soils to Streams? J. Geophys. Res. Biogeosci. 2013, 118, 291–302. [Google Scholar] [CrossRef]
- Du, E.; de Vries, W.; Galloway, J.N.; Hu, X.; Fang, J. Changes in Wet Nitrogen Deposition in the United States between 1985 and 2012. Environ. Res. Lett. 2014, 9, 95004. [Google Scholar] [CrossRef]
- Pereira-Silva, E.F.L.; Casals, P.; Sodek, L.; Delitti, W.B.C.; Vallejo, V.R. Post-Fire Nitrogen Uptake and Allocation by Two Resprouting Herbaceous Species with Contrasting Belowground Traits. Environ. Exp. Bot. 2019, 159, 157–167. [Google Scholar] [CrossRef]
- Pereira-Silva, E.F.L.; Hardt, E.; Fernandes, A.O. The Soil-Plant Relationship of Nitrogen Use in Three Tropical Tree Species. Web Ecol. 2012, 12, 57–64. [Google Scholar] [CrossRef]
- Stewart, G.R.; Pate, J.S.; Unkovich, M. Characteristics of Inorganic Nitrogen Assimilation of Plants in Fire-prone Mediterranean-type Vegetation. Plant Cell Environ. 1993, 16, 351–363. [Google Scholar] [CrossRef]
- Usman, S.; Singh, S.P.; Rawat, Y.S.; Bargali, S.S. Fine Root Decomposition and Nitrogen Mineralisation Patterns in Quercus Leucotrichophora and Pinus Roxburghii Forests in Central Himalaya. For. Ecol. Manag. 2000, 131, 191–199. [Google Scholar] [CrossRef]
- Schimann, H.; Ponton, S.; Hättenschwiler, S.; Ferry, B.; Lensi, R.; Domenach, A.-M.; Roggy, J.-C. Differing Nitrogen Use Strategies of Two Tropical Rainforest Late Successional Tree Species in French Guiana: Evidence from 15N Natural Abundance and Microbial Activities. Soil Biol. Biochem. 2008, 40, 487–494. [Google Scholar] [CrossRef]
- Pereira-Silva, E.F.L.; Hardt, E.; Biral, M.B.; Keller, V.C.; Delitti, W.B.C. Effects of Recent Fire on Soil Conditions and Nutrient Use of a Native and an Invasive Grass in the Brazilian Savanna. Écoscience 2019, 26, 359–370. [Google Scholar] [CrossRef]
- de Araújo, V.B.F.; Justino, G.C.; da Silva, C.E.M.; Vera, S.A.A.; Lins, L.; Sodek, L.; de Camargos, L.S.; Pereira, L.O.V.; de Carvalho Gonçalves, J.F. Soil Nitrogen Recovery and Seasonal Changes of Xylem Sap Amino Acids of Amazonian Tree Species Following Pasture Abandonment. Plant Ecol. 2019, 220, 633–648. [Google Scholar] [CrossRef]
- Rubio-Covarrubias, O.A.; Brown, P.H.; Weinbaum, S.A.; Johnson, R.S.; Cabrera, R.I. Evaluating Foliar Nitrogen Compounds as Indicators of Nitrogen Status in Prunus Persica Trees. Sci. Hortic. 2009, 120, 27–33. [Google Scholar] [CrossRef]
- Chapin, F.S.; Follett, J.M.; O’Connor, K.F. Growth, Phosphate Absorption, and Phosphorus Chemical Fractions in Two Chionochloa Species. J. Ecol. 1982, 70, 305–321. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Koba, K.; Koyama, L.A.; Hobbie, S.E.; Weiss, M.S.; Inagaki, Y.; Shaver, G.R.; Giblin, A.E.; Hobara, S.; Nadelhoffer, K.J.; et al. Nitrate Is an Important Nitrogen Source for Arctic Tundra Plants. Proc. Natl. Acad. Sci. USA 2018, 115, 201715382. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, R.A.; Lea, P.J. Lysine Metabolism in Higher Plants. Amino Acids 2001, 20, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Stock, W.D.; Lewis, O.A.M. Extraction of Nitrate Reductase from Members of the South African Proteaceae. S. Afr. J. Bot. 1982, 1, 124–126. [Google Scholar] [CrossRef][Green Version]
- Schmidt, S.; Stewart, G.R. Transport, Storage and Mobilization of Nitrogen by Trees and Shrubs in the Wet/Dry Tropics of Northern Australia. Tree Physiol. 1998, 18, 403–410. [Google Scholar] [CrossRef][Green Version]
- Aidar, M.P.M.; Schmidt, S.; Moss, G.; Stewart, G.R.; Joly, C.A. Nitrogen Use Strategies of Neotropical Rainforest Trees in Threatened Atlantic Forest. Plant Cell Environ. 2003, 26, 389–399. [Google Scholar] [CrossRef]
- Wyse, S.V. Nitrate Reductase Activity in Plant Species of Varied Spatial Association with Acidic Soils beneath Agathis Australis. N. Z. J. Bot. 2014, 52, 213–223. [Google Scholar] [CrossRef]
- Craine, J.M.; Elmore, A.J.; Aidar, M.P.M.; Bustamante, M.; Dawson, T.E.; Hobbie, E.A.; Kahmen, A.; Mack, M.C.; McLauchlan, K.K.; Michelsen, A.; et al. Global Patterns of Foliar Nitrogen Isotopes and Their Relationships with Climate, Mycorrhizal Fungi, Foliar Nutrient Concentrations, and Nitrogen Availability. New Phytol. 2009, 183, 980–992. [Google Scholar] [CrossRef]
- McKane, R.B.; Johnson, L.C.; Shaver, G.R.; Nadelhoffer, K.J.; Rastetter, E.B.; Fry, B.; Giblin, A.E.; Kielland, K.; Kwiatkowski, B.L.; Laundre, J.A.; et al. Resource-Based Niches Provide a Basis for Plant Species Diversity and Dominance in Arctic Tundra. Nature 2002, 415, 68–71. [Google Scholar] [CrossRef]
- Rascher, K.G.; Hellmann, C.; Máguas, C.; Werner, C. Community Scale 15N Isoscapes: Tracing the Spatial Impact of an Exotic N2-Fixing Invader. Ecol. Lett. 2012, 15, 484–491. [Google Scholar] [CrossRef]
- Craine, J.M.; Brookshire, E.N.J.; Cramer, M.D.; Hasselquist, N.J.; Koba, K.; Marin-Spiotta, E.; Wang, L. Ecological Interpretations of Nitrogen Isotope Ratios of Terrestrial Plants and Soils. Plant Soil 2015, 396, 1–26. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen Uptake, Assimilation and Remobilization in Plants: Challenges for Sustainable and Productive Agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M. Transporters Involved in Source to Sink Partitioning of Amino Acids and Ureides: Opportunities for Crop Improvement. J. Exp. Bot. 2014, 65, 1865–1878. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M.; Masclaux-Daubresse, C. Source and Sink Mechanisms of Nitrogen Transport and Use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J.; Fan, X.; Shen, Q.; Smith, S.J. Amino Acids and Nitrate as Signals for the Regulation of Nitrogen Acquisition. J. Exp. Bot. 2007, 59, 111–119. [Google Scholar] [CrossRef]
- Kusano, M.; Fukushima, A.; Redestig, H.; Saito, K. Metabolomic Approaches toward Understanding Nitrogen Metabolism in Plants. J. Exp. Bot. 2011, 62, 1439–1453. [Google Scholar] [CrossRef]
- Govaerts, R.; Sobral, M.; Ashton, P.; Barrie, F.; Holst, B.K.; Landrum, L.L.; Matsumoto, K.; Mazine, F.F.; Lughadha, E.N.; Proneça, C.; et al. World Checklist of Myrtaceae; Royal Botanic Gardens: Richmond, BC, Canada, 2008; ISBN 978−1-84246-391-8. [Google Scholar]
- Binkley, D.; Matson, P. Ion Exchange Resin Bag Method for Assessing Forest Soil Nitrogen Availability. Soil Sci. Soc. Am. J. 1983. [Google Scholar] [CrossRef]
- Hardt, E.; Pereira-Silva, E.F.L.; Dos Santos, R.F.; Tamashiro, J.Y.; Ragazzi, S.; Lins, D.B.D.S. The Influence of Natural and Anthropogenic Landscapes on Edge Effects. Landsc. Urban Plan. 2013, 120. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid Colorimetric Determination of Nitrate in Plant Tissue by Nitration of Salicylic Acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- McCullough, H. The Determination of Ammonia in Whole Blood by a Direct Colorimetric Method. Clin. Chim. Acta 1967, 17, 297–304. [Google Scholar] [CrossRef]
- Jarrett, H.W.; Cooksy, K.D.; Ellis, B.; Anderson, J.M. The Separation of O-Phthalaldehyde Derivatives of Amino Acids by Reversed-Phase Chromatography on Octylsilica Columns. Anal. Biochem. 1986, 153, 189–198. [Google Scholar] [CrossRef]
- Puiatti, M.; Sodek, L. Waterlogging Affects Nitrogen Transport in the Xylem of Soybean. Plant Physiol. Biochem. 1999, 37, 767–773. [Google Scholar] [CrossRef]
- Laine, M.; Rütting, T.; Alakukku, L.; Palojärvi, A.; Strömmer, R. Process Rates of Nitrogen Cycle in Uppermost Topsoil after Harvesting in No-Tilled and Ploughed Agricultural Clay Soil. Nutr. Cycl. Agroecosyst. 2018, 110, 39–49. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Morgan, J.A.; Blumenthal, D.; Follett, R.F. Water Limitation and Plant Inter-Specific Competition Reduce Rhizosphere-Induced C Decomposition and Plant N Uptake. Soil Biol. Biochem. 2010, 42, 1073–1082. [Google Scholar] [CrossRef]
- Niboyet, A.; Le Roux, X.; Dijkstra, P.; Hungate, B.A.; Barthes, L.; Blankinship, J.C.; Brown, J.R.; Field, C.B.; Leadley, P.W. Testing Interactive Effects of Global Environmental Changes on Soil Nitrogen Cycling. Ecosphere 2011, 2, art56. [Google Scholar] [CrossRef]
- Rothstein, D.E.; Zak, D.R.; Pregitzer, K.S. Nitrate Deposition in Northern Hardwood Forests and the Nitrogen Metabolism of Acer Saccharum Marsh. Oecologia 1996, 108, 338–344. [Google Scholar] [CrossRef]
- Prado, C.H.B.A.; De Moraes, J.A.P.V. Photosynthetic Capacity and Specific Leaf Mass in Twenty Woody Species of Cerrado Vegetation under Field Conditions. Photosynthetica 1997, 33, 103–112. [Google Scholar] [CrossRef]
- Havill, D.C.; Lee, J.A.; De-Felice, J. Some Factors Limiting Nitrate Utilization in Acidic and Calcareous Grasslands. New Phytol. 1977, 78, 649–659. [Google Scholar] [CrossRef]
- SMIRNOFF, N.; TODD, P.; STEWART, G.R. The Occurrence of Nitrate Reduction in the Leaves of Woody Plants. Ann. Bot. 1984, 54, 363–374. [Google Scholar] [CrossRef]
- Gebauer, G.; Rehder, H.; Wollenweber, B. Nitrate, Nitrate Reduction and Organic Nitrogen in Plants from Different Ecological and Taxonomic Groups of Central Europe. Oecologia 1988, 75, 371–385. [Google Scholar] [CrossRef]
- Miller, A.J.; Cramer, M.D. Root Nitrogen Acquisition and Assimilation. Plant Soil 2005, 274, 1–36. [Google Scholar] [CrossRef]
- Peuke, A.D. Correlations in Concentrations, Xylem and Phloem Flows, and Partitioning of Elements and Ions in Intact Plants. A Summary and Statistical Re-Evaluation of Modelling Experiments in Ricinus Communis. J. Exp. Bot. 2009, 61, 635–655. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B. The World-Wide ‘Fast–Slow’ Plant Economics Spectrum: A Traits Manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Tanaka-Oda, A.; Kenzo, T.; Inoue, Y.; Yano, M.; Koba, K.; Ichie, T. Variation in Leaf and Soil Δ15N in Diverse Tree Species in a Lowland Dipterocarp Rainforest, Malaysia. Trees 2016, 30, 509–522. [Google Scholar] [CrossRef]
- Galili, G. The Aspartate-Family Pathway of Plants: Linking Production of Essential Amino Acids with Energy and Stress Regulation. Plant Signal. Behav. 2011, 6, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Okumoto, S.; Pilot, G. Amino Acid Export in Plants: A Missing Link in Nitrogen Cycling. Mol. Plant 2011, 4, 453–463. [Google Scholar] [CrossRef] [PubMed]

| Species | Authors | Acr | n | Forest Type |
|---|---|---|---|---|
| Myrcia lingua | (O. Berg) Mattos & D. Legrand | Mli | 3 | SW |
| Campomanesia pubescens | (DC.) O. Berg | Cpu | 3 | SW |
| Eugenia cuprea | (O. Berg) Mattos | Ecu | 3 | OD |
| Marlierea suaveolens | Cambess. | Msu | 3 | OD |
| Myrciaria floribunda | (H. West ex Willd.) O. Berg | Mfl | 3 | SF |
| Plinia cauliflora | (DC.) Kausel | Pca | 3 | SF |
| Campomanesia guaviroba | (DC.) Kiaersk. | Cgu | 3 | SF |
| Eugenia excelsa | O. Berg | Eex | 3 | SF |
| Soil Chemical Properties | Forest Type | SW | OD | SF | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | 11 | 23 | 12 | |||||||||||
| Unit | ||||||||||||||
| pH | CaCI2 | 3.8 | ± | 0.02 | a | 3.7 | ± | 0.03 | b | 4.3 | ± | 0.34 | c | |
| K+ | mmolc/dm3 | 1.3 | ± | 0.13 | a | 3.5 | ± | 0.31 | b | 2.2 | ± | 0.77 | b | |
| Ca+2 | 1.4 | ± | 0.36 | a | 18.2 | ± | 2.2 | b | 38.2 | ± | 18.3 | b | ||
| Mg+2 | 1.3 | ± | 0.27 | a | 8.9 | ± | 0.92 | b | 10.9 | ± | 5.0 | b | ||
| H+ + Al+3 | 53.9 | ± | 0.73 | a | 98.6 | ± | 3.3 | b | 101.2 | ± | 27.8 | b | ||
| Al+3 | 13.9 | ± | 0.34 | a | 18.2 | ± | 1.6 | b | ||||||
| S | mg/dm3 | 9.1 | ± | 0.80 | a | 8.1 | ± | 0.90 | a | 7.9 | ± | 2.1 | a | |
| P | 3.3 | ± | 0.75 | a | 14.7 | ± | 1.5 | b | 23.6 | ± | 4.6 | c | ||
| OM | g/dm3 | 28.6 | ± | 1.1 | a | 44.3 | ± | 3.3 | b | 65.3 | ± | 5.7 | c | |
| OC | 16.6 | ± | 0.9 | a | 25.7 | ± | 1.8 | b | 37.9 | ± | 3.2 | c | ||
| CEC | mmolc/dm3 | 57.8 | ± | 1.3 | a | 130.3 | ± | 5.0 | b | 152.4 | ± | 28.6 | b | |
| SB | % | 3.9 | ± | 0.72 | a | 31.6 | ± | 3.3 | b | 51.3 | ± | 22.8 | b | |
| V | 6.6 | ± | 0.97 | a | 23.5 | ± | 1.9 | b | 33.6 | ± | 13.5 | b | ||
| NH4+ | µg/g | wet | 3.0 | ± | 0.09 | aAA | 2.7 | ± | 0.08 | bAA | 13.3 | ± | 1.5 | cAA |
| dry | 1.1 | ± | 0.07 | aAB | 1.4 | ± | 0.11 | bAB | 8.7 | ± | 1.6 | cAB | ||
| NO3− | wet | 1.0 | ± | 0.20 | aBA | 1.1 | ± | 0.34 | aBA | 3.1 | ± | 0.61 | bBA | |
| dry | 0.06 | ± | 0.01 | aBB | 0.03 | ± | 0.02 | aBB | 10.3 | ± | 1.5 | bAB | ||
| F | S | Leaves | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acr | NRA | NO3− | δ15N | Ntotal | C:N | |||||||||||||||||
| pkat g−1 FW | mol g−1 FW | ‰ | % | |||||||||||||||||||
| Mli | SW | wet | 13.5 | ± | 1.0 | a | −0.2 | ± | 0.12 | a | 1.0 | ± | 0.01 | a | 39.9 | ± | 1.8 | a | ||||
| dry | 10.1 | ± | 2.9 | a | −0.17 | ± | 0.12 | a | 1.2 | ± | 0.1 | a | 37.8 | ± | 2.7 | |||||||
| Cpu | SW | wet | 59.5 | ± | 4.3 | aB | −0.11 | ± | 0.08 | a | 0.3 | ± | 0.14 | aB | 46.9 | ± | 2.2 | aB | ||||
| dry | 49.8 | ± | 5.4 | aA | −0.19 | ± | 0.01 | a | 0.4 | ± | 0.04 | aB | 43.1 | ± | 7.1 | aB | ||||||
| Ecu | OD | wet | 98.9 | ± | 62.3 | aA | 6.0 | ± | 1.0 | aA | 1.1 | ± | 1.3 | aA | 2.1 | ± | 0.7 | aA | 22.4 | ± | 6.6 | aA |
| dry | 38.0 | ± | 11.8 | aA | 6.9 | ± | 0.9 | aA | 0.5 | ± | 0.6 | aB | 1.9 | ± | 0.2 | aA | 24.6 | ± | 2.7 | aA | ||
| Msu | OD | wet | 104.2 | ± | 10.8 | a | 5.0 | ± | 0.9 | a | 0.7 | ± | 1.2 | a | 1.5 | ± | 0.1 | a | 30.4 | ± | 3.1 | a |
| dry | 87.2 | ± | 34.0 | a | 4.8 | ± | 0.9 | a | 1.0 | ± | 1.1 | a | 2.1 | ± | 1.0 | a | 25.6 | ± | 10.2 | a | ||
| Mfl | SF | wet | 134.4 | ± | 105.9 | a | 6.6 | ± | 2.0 | a | 2.2 | ± | 1.2 | a | 1.9 | ± | 0.1 | a | 24.0 | ± | 1.8 | a |
| dry | 106.4 | ± | 86.9 | a | 5.3 | ± | 1.2 | a | 1.4 | ± | 1.7 | a | 2.0 | ± | 0.7 | a | 23.7 | ± | 7.3 | a | ||
| Pca | SF | wet | 114.5 | ± | 55.0 | a | 5.1 | ± | 4.9 | a | 3.0 | ± | 1.3 | a | 2.1 | ± | 0.2 | a | 22.0 | ± | 1.7 | a |
| dry | 205.9 | ± | 10.2 | b | 7.4 | ± | 1.3 | a | 2.5 | ± | 0.6 | a | 2.0 | ± | 0.4 | a | 22.8 | ± | 3.4 | a | ||
| Cgu | SF | wet | 26.7 | ± | 6.4 | aA | 10.1 | ± | 0.4 | a | 2.4 | ± | 0.2 | a | 2.4 | ± | 0.4 | aA | 19.4 | ± | 2.9 | aA |
| dry | 52.2 | ± | 65.0 | aA | 10.2 | ± | 0.1 | a | 3.3 | ± | 1.2 | a | 2.0 | ± | 0.02 | aA | 23.1 | ± | 0.2 | aA | ||
| Eex | SF | wet | 110.6 | ± | 93.9 | aA | 8.4 | ± | 1.9 | aA | 2.6 | ± | 0.4 | aA | 2.0 | ± | 0.2 | aA | 23.2 | ± | 2.3 | aA |
| dry | 24.5 | ± | 16.3 | aA | 9.9 | ± | 6.1 | aA | 2.8 | ± | 1.0 | aA | 2.1 | ± | 0.4 | aA | 22.5 | ± | 4.6 | aA | ||
| F | S | Xylem sap | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acr | Nitrogen Transported (%) | |||||||||||||||
| NO3− | ASP | GLU | ASN | GLN | ARG | CIT | ALA | PHE | MET | VAL | LEU | LYS | Others | |||
| Ecu | OD | wet | 3.3 | 4.4 | 8.8 | 4.2 | 27.8 | 0.2 | 43.6 | 7.6 | ||||||
| dry | 3.7 | 2.6 | 5.9 | 0.8 | 64.8 | 0.2 | 18.8 | 3.2 | ||||||||
| Msu | OD | wet | 3.6 | 8.2 | 11.7 | 3.8 | 44.5 | 18.1 | 10.0 | |||||||
| dry | 0.7 | 5.8 | 9.4 | 6.1 | 11.0 | 1.3 | 58.6 | 7.0 | ||||||||
| Mfl | SF | wet | 0.6 | 2.7 | 7.7 | 0.5 | 50.1 | 11.0 | 25.5 | 2.0 | ||||||
| dry | 0.6 | 4.6 | 6.5 | 6.2 | 41.3 | 2.4 | 20.1 | 18.4 | ||||||||
| Pca | SF | wet | 0.6 | 2.5 | 7.9 | 1.1 | 54.0 | 10.9 | 16.8 | 6.2 | ||||||
| dry | 0.8 | 6.1 | 18.2 | 0.0 | 18.3 | 3.4 | 15.8 | 25.2 | 12.2 | |||||||
| Cgu | SF | wet | 0.8 | 0.0 | 4.1 | 23.7 | 0.0 | 20.9 | 15.8 | 12.8 | 21.9 | |||||
| dry | 0.7 | 2.7 | 10.7 | 0.0 | 8.3 | 3.7 | 20.9 | 48.4 | 4.6 | |||||||
| Eex | SF | wet | 0.9 | 3.6 | 10.5 | 0.8 | 11.9 | 67.3 | 5.1 | |||||||
| dry | 0.9 | 5.0 | 24.7 | 0.2 | 8.3 | 3.0 | 48.2 | 9.5 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira-Silva, E.F.L.; Joly, C.; Sodek, L.; Hardt, E.; Aidar, M. Characterization of Nitrogen Use by Neotropical Myrtaceae in Dry and Wet Forests of Southeast Brazil. Earth 2022, 3, 1290-1304. https://doi.org/10.3390/earth3040073
Pereira-Silva EFL, Joly C, Sodek L, Hardt E, Aidar M. Characterization of Nitrogen Use by Neotropical Myrtaceae in Dry and Wet Forests of Southeast Brazil. Earth. 2022; 3(4):1290-1304. https://doi.org/10.3390/earth3040073
Chicago/Turabian StylePereira-Silva, Erico Fernando Lopes, Carlos Joly, Ladaslav Sodek, Elisa Hardt, and Marcos Aidar. 2022. "Characterization of Nitrogen Use by Neotropical Myrtaceae in Dry and Wet Forests of Southeast Brazil" Earth 3, no. 4: 1290-1304. https://doi.org/10.3390/earth3040073
APA StylePereira-Silva, E. F. L., Joly, C., Sodek, L., Hardt, E., & Aidar, M. (2022). Characterization of Nitrogen Use by Neotropical Myrtaceae in Dry and Wet Forests of Southeast Brazil. Earth, 3(4), 1290-1304. https://doi.org/10.3390/earth3040073