
Does Climate Change Affect the Yield of the Top Three Cereals and Food Security in the World?

 , , and
, , and
Abstract
1. Introduction
2. Current and Future Projected Areas under Cultivation and Total Production of Top Three Cereals




3. Significance and Requirements of Optimum Growing Conditions for the Top Three Cereals
3.1. Rice
3.2. Maize
3.3. Wheat

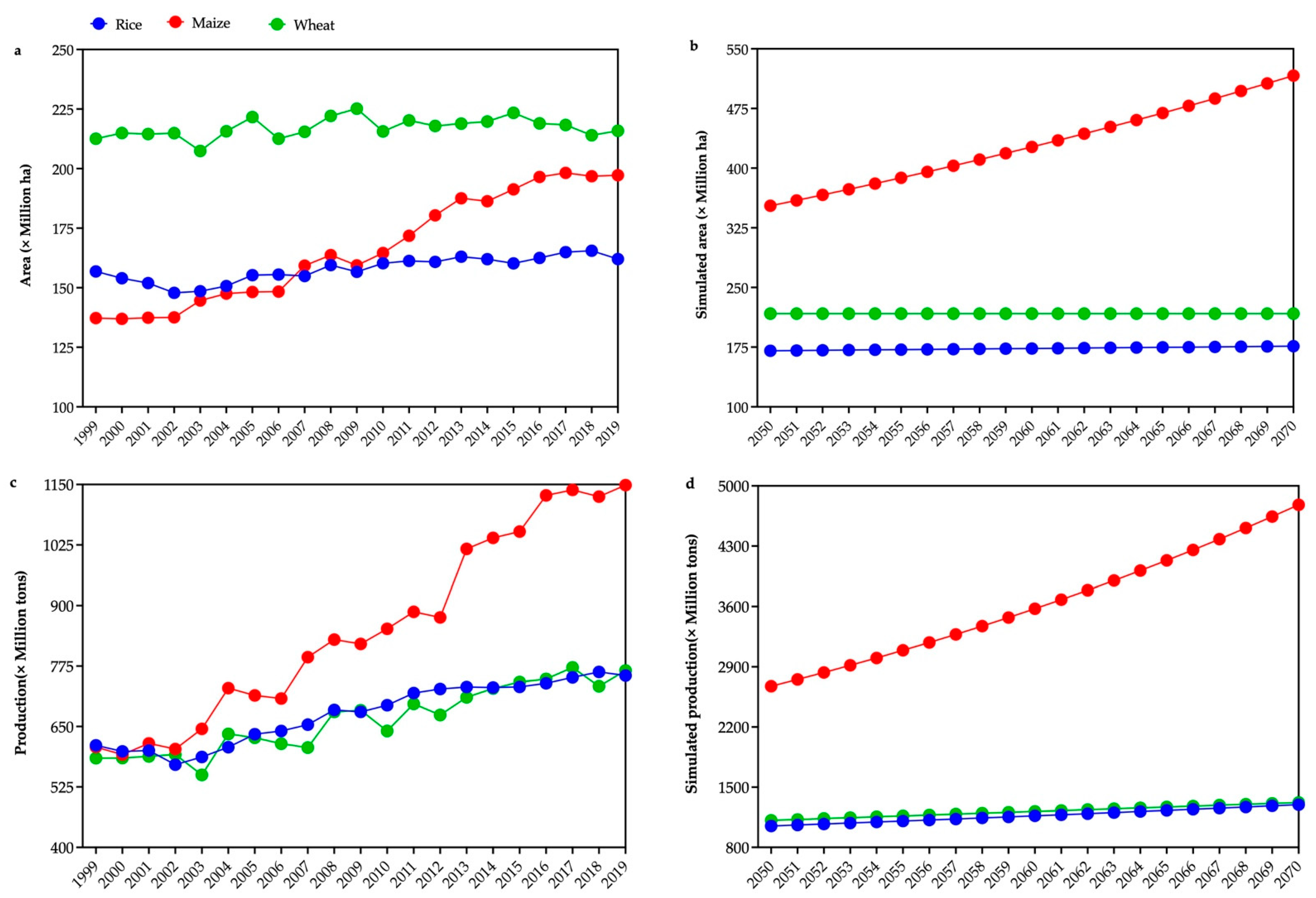{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Climatic Conditions | |||||||
| Crops | Irrigation (mm) | Temperature | Sunshine (h/Day) | Duration of Growth (d) | Photosynthesis Pathway | References | |
| (°C) | |||||||
| Rice | 500 to 60 (up to 829 mm [3]) | 22 to 31 | 4–6 | 90–120 | C3 | [39,59] | |
| Wheat | 60–90 | −3 to 23 | 4–6 | 120–180 | C3 | [47,59] | |
| Maize | 200–450 | 11 to 30 | 6–7 | 90–110 | C4 | [44,59] | |
| (b) Soil, Fertilizer, and Pesticide Requirements | |||||||
| Crops | Soil pH | Soil Type | N | P | K | Pesticide | References |
| (kg/ha) | (kg/ha) | (kg/ha) | |||||
| Rice | 5.0–6.5 | Flooded condition | 90–120 | 30–40 | 40–60 | Applied | [60,61] |
| Wheat | 5.5–6.5 | Sandy loam | 70–200 | 20–40 | 80–100 | Applied | [47,59,62,63] |
| Maize | 5.8–7.0 | Warm and silt loam | 125–160 | 55–80 | 85–110 | Applied | [44,45] |
4. Greenhouse Gas Emissions and Global Warming Potential from Multiple Crops
5. Cereal Crop Yield and Climate Change
6. Morphological, Physiological, and Biochemical Responses of Plants to Drought
7. Responses of the Plant Hormone in Abiotic Stresses
8. Strategies to Combat Climate Change and Increase Crop Yield
8.1. Conventional Breeding Techniques
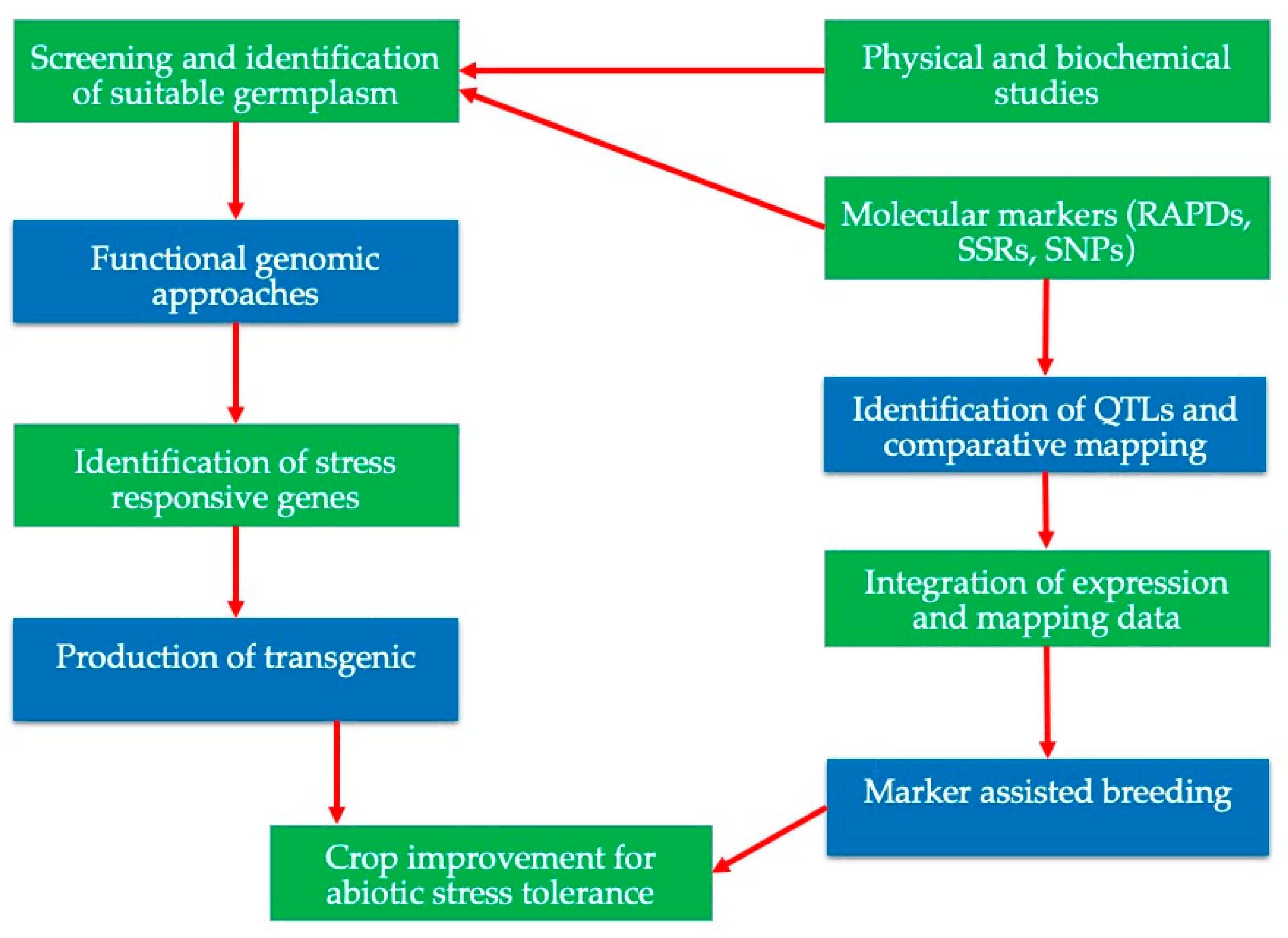
8.2. Modern Molecular Techniques and the Genomic Approach

8.3. Agronomic Best Management (ABM) Approaches
8.4. Cultivation of More Climate-Resilient Cereal Crops
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Bagale, S. Climate ready crops for drought stress: A review in Nepalese context. Rev. Food Agric. 2021, 2, 83–87. [Google Scholar] [CrossRef]
- Wang, J.; Vanga, S.K.; Saxena, R.; Orsat, V.; Raghavan, V. Effect of climate change on the yield of cereal crops: A review. Climate 2018, 6, 41. [Google Scholar] [CrossRef]
- Noya, I.; González-García, S.; Bacenetti, J.; Fiala, M.; Moreira, M.T. Environmental impacts of the cultivation-phase associated with agricultural crops for feed production. J. Clean. Prod. 2018, 172, 3721–3733. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Pye-Smith, C. Farming’s Climate-Smart Future: Placing Agriculture at the Heart of Climate-Change Polic. Wageningen, Netherlands: CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS) and the Technical Centre for Agricultural and Rural Cooperation (CTA). 2011. Available online: https://cgspace.cgiar.org/handle/10568/25164 (accessed on 17 December 2021).
- Hawkesford, M.J.; Araus, J.L.; Park, R.; Calderini, D.; Miralles, D.; Shen, T.; Zhang, J.; Parry, M.A. Prospects of doubling global wheat yields. Food Energy Secur. 2013, 2, 34–48. [Google Scholar] [CrossRef]
- Hussain, S.G. Assessing impacts of climate change on cereal production and food security in Bangladesh. In Climate Change and Food Security in South Asia; Springer: Berlin/Heidelberg, Germany, 2010; pp. 459–476. [Google Scholar]
- Naumann, G.; Alfieri, L.; Wyser, K.; Mentaschi, L.; Betts, R.; Carrao, H.; Spinoni, J.; Vogt, J.; Feyen, L. Global changes in drought conditions under different levels of warming. Geophys. Res. Lett. 2018, 45, 3285–3296. [Google Scholar] [CrossRef]
- Neupane, D.; Mayer, J.A.; Niechayev, N.A.; Bishop, C.D.; Cushman, J.C. Five-year field trial of the biomass productivity and water input response of cactus pear (Opuntia spp.) as a bioenergy feedstock for arid lands. GCB Bioenergy 2021, 13, 719–741. [Google Scholar] [CrossRef]
- Butler, E.E.; Huybers, P. Adaptation of US maize to temperature variations. Nat. Clim. Chang. 2013, 3, 68–72. [Google Scholar] [CrossRef]
- Jin, Z.; Zhuang, Q.; Wang, J.; Archontoulis, S.V.; Zobel, Z.; Kotamarthi, V.R. The combined and separate impacts of climate extremes on the current and future US rainfed maize and soybean production under elevated CO2. Glob. Chang. Biol. 2017, 23, 2687–2704. [Google Scholar] [CrossRef] [PubMed]
- Basso, B.; Ritchie, J.T. Evapotranspiration in high-yielding maize and under increased vapor pressure deficit in the US Midwest. Agric. Environ. Lett. 2018, 3, 170039. [Google Scholar] [CrossRef]
- Abendroth, L.J.; Miguez, F.E.; Castellano, M.J.; Carter, P.R.; Messina, C.D.; Dixon, P.M.; Hatfield, J.L. Lengthening of maize maturity time is not a widespread climate change adaptation strategy in the US Midwest. Glob. Chang. Biol. 2021, 27, 2426–2440. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2007: Impacts, Adaptation and Vulnerability: Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007; Volume 4. [Google Scholar]
- FAO. The State of Food Insecurity in the World 2014: Strengthening the Enabling Enviroment for Food Security and Nutrition; Food & Agriculture Org.: Rome, Italy, 2015. [Google Scholar]
- Wu, J.-Z.; Zhang, J.; Ge, Z.-M.; Xing, L.-W.; Han, S.-Q.; Chen, S.; Kong, F.-T. Impact of climate change on maize yield in China from 1979 to 2016. J. Integr. Agric. 2021, 20, 289–299. [Google Scholar] [CrossRef]
- Mashizha, T.M. Building adaptive capacity: Reducing the climate vulnerability of smallholder farmers in Zimbabwe. Bus. Strategy Dev. 2019, 2, 166–172. [Google Scholar] [CrossRef]
- Bannayan, M.; Paymard, P.; Ashraf, B. Vulnerability of maize production under future climate change: Possible adaptation strategies. J. Sci. Food Agric. 2016, 96, 4465–4474. [Google Scholar] [CrossRef] [PubMed]
- Soglo, Y.Y.; Nonvide, G.M.A. Climate change perceptions and responsive strategies in Benin: The case of maize farmers. Clim. Chang. 2019, 155, 245–256. [Google Scholar] [CrossRef]
- Akram, N. Is climate change hindering economic growth of Asian economies. Asia-Pac. Dev. J. 2013, 19, 1–18. [Google Scholar] [CrossRef]
- Bevan, M.; Waugh, R. Applying plant genomics to crop improvement. BioMed Cent. 2007, 8, 302. [Google Scholar]
- Hunter, D.; Borelli, T.; Beltrame, D.M.; Oliveira, C.N.; Coradin, L.; Wasike, V.W.; Wasilwa, L.; Mwai, J.; Manjella, A.; Samarasinghe, G.W. The potential of neglected and underutilized species for improving diets and nutrition. Planta 2019, 250, 709–729. [Google Scholar] [CrossRef] [PubMed]
- Hendre, P.S.; Muthemba, S.; Kariba, R.; Muchugi, A.; Fu, Y.; Chang, Y.; Song, B.; Liu, H.; Liu, M.; Liao, X. African Orphan Crops Consortium (AOCC): Status of developing genomic resources for African orphan crops. Planta 2019, 250, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Zenda, T.; Liu, S.; Dong, A.; Duan, H. Advances in Cereal Crop Genomics for Resilience under Climate Change. Life 2021, 11, 502. [Google Scholar] [CrossRef]
- Mabhaudhi, T.; Chimonyo, V.G.P.; Hlahla, S.; Massawe, F.; Mayes, S.; Nhamo, L.; Modi, A.T. Prospects of orphan crops in climate change. Planta 2019, 250, 695–708. [Google Scholar] [CrossRef]
- Scheben, A.; Yuan, Y.; Edwards, D. Advances in genomics for adapting crops to climate change. Curr. Plant Biol. 2016, 6, 2–10. [Google Scholar] [CrossRef]
- Pourkheirandish, M.; Golicz, A.A.; Bhalla, P.L.; Singh, M.B. Global role of crop genomics in the face of climate change. Front. Plant Sci. 2020, 11, 922. [Google Scholar] [CrossRef]
- Assefa, Y.; Prasad, P.V.; Foster, C.; Wright, Y.; Young, S.; Bradley, P.; Stamm, M.; Ciampitti, I.A. Major management factors determining spring and winter canola yield in North America. Crop Sci. 2018, 58, 1–16. [Google Scholar] [CrossRef]
- Roser, M.; Ritchie, H. Land Use. 2013. Published online at OurWorldInData.org. Available online: https://ourworldindata.org/land-use (accessed on 19 July 2021).
- USDA. World Agricultural Production. United States Department of Agriculture, Foreign Agricultural Service, Circular Series WAP 7–21. 2021. Available online: https://apps.fas.usda.gov/psdonline/circulars/production.pdf (accessed on 19 July 2021).
- FAOSTAT. Crops. Food and Agriculture Organization of the United Nations. 2021. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 19 July 2021).
- Maskey, R. Population growth and food production: Issues, problems and prospects. Outlook Agric. 2001, 30, 155–160. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar]
- Ma, L.; Hu, C.; Lin, R.; Han, Y. ARIMA model forecast based on EViews software. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Proceedings of the 2018 International Conference on Air Pollution and Environmental Engineering (APEE 2018), Hong Kong, China, 26–28 October 2018; IOP Publishing: Bristol, UK, 2018; Volume 208, p. 012017. [Google Scholar]
- Datta, A.; Ullah, H.; Ferdous, Z. Water management in rice. In Rice Production Worldwide; Springer: Berlin/Heidelberg, Germany, 2017; pp. 255–277. [Google Scholar]
- Bandumula, N. Rice production in Asia: Key to global food security. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 1323–1328. [Google Scholar] [CrossRef]
- Smith, C.W.; Dilday, R.H. Rice: Origin, History, Technology, and Production; John Wiley & Sons: Hoboken, NJ, USA, 2002; Volume 3. [Google Scholar]
- Yao, F.; Huang, J.; Cui, K.; Nie, L.; Xiang, J.; Liu, X.; Wu, W.; Chen, M.; Peng, S. Agronomic performance of high-yielding rice variety grown under alternate wetting and drying irrigation. Field Crops Res. 2012, 126, 16–22. [Google Scholar] [CrossRef]
- Kandil, E. Response of some maize hybrids (Zea mays L.) to different levels of nitrogenous fertilization. J. Appl. Sci. Res. 2013, 9, 1902–1908. [Google Scholar]
- Piperno, D.R.; Flannery, K.V. The earliest archaeological maize (Zea mays L.) from highland Mexico: New accelerator mass spectrometry dates and their implications. Proc. Natl. Acad. Sci. USA 2001, 98, 2101–2103. [Google Scholar] [CrossRef] [PubMed]
- Hilaire, L. Corn: An American Native. Spanning Gap Newsl. Del. Water Gap Natl. Recreat. Area 2000, 22, 2. [Google Scholar]
- Zamir, M.; Yasin, G.; Javeed, H.; Ahmad, A.; Tanveer, A.; Yaseen, M. Effect of different sowing techniques and mulches on the growth and yield behavior of spring planted maize (Zea mays L.). Cercet. Agron. Mold. 2013, 46, 7782. [Google Scholar] [CrossRef]
- Woldesenbet, M.; Haileyesus, A. Effect of nitrogen fertilizer on growth, yield and yield components of maize (Zea mays L.) in Decha district, Southwestern Ethiopia. Int. J. Res.-Granthaalayah 2016, 4, 95–100. [Google Scholar] [CrossRef]
- Yin, G.; Gu, J.; Zhang, F.; Hao, L.; Cong, P.; Liu, Z. Maize yield response to water supply and fertilizer input in a semi-arid environment of Northeast China. PLoS ONE 2014, 9, e86099. [Google Scholar]
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef]
- Ngwako, S.; Mashiqa, P. The effect of irrigation on the growth and yield of winter wheat (Triticum aestivum L.) cultivars. Int. J. Agric. Crop Sci. 2013, 5, 976–982. [Google Scholar]
- Liu, B.; Martre, P.; Ewert, F.; Porter, J.R.; Challinor, A.J.; Müller, C.; Ruane, A.C.; Waha, K.; Thorburn, P.J.; Aggarwal, P.K. Global wheat production with 1.5 and 2.0 °C above pre-industrial warming. Glob. Chang. Biol. 2019, 25, 1428–1444. [Google Scholar] [CrossRef] [PubMed]
- Schleussner, C.-F.; Deryng, D.; Müller, C.; Elliott, J.; Saeed, F.; Folberth, C.; Liu, W.; Wang, X.; Pugh, T.A.; Thiery, W. Crop productivity changes in 1.5 °C and 2 °C worlds under climate sensitivity uncertainty. Environ. Res. Lett. 2018, 13, 064007. [Google Scholar] [CrossRef]
- Bhatt, R.; Hossain, A.; Hasanuzzaman, M. Adaptation strategies to mitigate the evapotranspiration for sustainable crop production: A perspective of rice-wheat cropping system. In Agronomic Crops; Springer: Berlin/Heidelberg, Germany, 2019; pp. 559–581. [Google Scholar]
- Bhattacharyya, R.; Das, T.; Das, S.; Dey, A.; Patra, A.; Agnihotri, R.; Ghosh, A.; Sharma, A. Four years of conservation agriculture affects topsoil aggregate-associated 15nitrogen but not the 15nitrogen use efficiency by wheat in a semi-arid climate. Geoderma 2019, 337, 333–340. [Google Scholar] [CrossRef]
- Dey, A.; Dwivedi, B.S.; Bhattacharyya, R.; Datta, S.P.; Meena, M.C.; Jat, R.K.; Gupta, R.K.; Jat, M.L.; Singh, V.K.; Das, D. Effect of conservation agriculture on soil organic and inorganic carbon sequestration and lability: A study from a rice–wheat cropping system on a calcareous soil of the eastern Indo-Gangetic Plains. Soil Use Manag. 2020, 36, 429–438. [Google Scholar] [CrossRef]
- Bhattacharyya, R.; Das, T.; Sudhishri, S.; Dudwal, B.; Sharma, A.; Bhatia, A.; Singh, G. Conservation agriculture effects on soil organic carbon accumulation and crop productivity under a rice–wheat cropping system in the western Indo-Gangetic Plains. Eur. J. Agron. 2015, 70, 11–21. [Google Scholar] [CrossRef]
- Dey, A.; Dwivedi, B.; Meena, M.; Datta, S. Dynamics of soil carbon and nitrogen under conservation agriculture in rice-wheat cropping system. Indian J. Fertil. 2018, 14, 12–26. [Google Scholar]
- Parihar, C.M.; Singh, A.; Jat, S.L.; Dey, A.; Nayak, H.S.; Mandal, B.N.; Saharawat, Y.S.; Jat, M.L.; Yadav, O.P. Soil quality and carbon sequestration under conservation agriculture with balanced nutrition in intensive cereal-based system. Soil Tillage Res. 2020, 202, 104653. [Google Scholar] [CrossRef]
- Sapkota, T.B.; Majumdar, K.; Jat, M.L.; Kumar, A.; Bishnoi, D.K.; McDonald, A.; Pampolino, M. Precision nutrient management in conservation agriculture based wheat production of Northwest India: Profitability, nutrient use efficiency and environmental footprint. Field Crops Res. 2014, 155, 233–244. [Google Scholar] [CrossRef]
- West, T.O.; Post, W.M. Soil organic carbon sequestration rates by tillage and crop rotation: A global data analysis. Soil Sci. Soc. Am. J. 2002, 66, 1930–1946. [Google Scholar] [CrossRef]
- Mazzoncini, M.; Sapkota, T.B.; Barberi, P.; Antichi, D.; Risaliti, R. Long-term effect of tillage, nitrogen fertilization and cover crops on soil organic carbon and total nitrogen content. Soil Tillage Res. 2011, 114, 165–174. [Google Scholar] [CrossRef]
- Faostat, F. Production Agricultural Commodities; Food and Agriculture Organization of the United Nations: Roma, Italy, 2016. [Google Scholar]
- Guo, Z.; Chai, M.; Zhan, Z.; Chen, Z. Spatiotemporal variation of soil PH in the past 30 years of Guangdong Province, China. In Proceedings of the 2011 19th International Conference on Geoinformatics, IEEE, Shanghai, China, 24–26 June 2011; pp. 1–5. [Google Scholar]
- Santos, E.S.; Abreu, M.M.; Magalhães, M.C.; Viegas, W.; Amâncio, S.; Cordovil, C. Nutrients levels in paddy soils and flood waters from Tagus-Sado basin: The impact of farming system. In Proceedings of the EGU General Assembly Conference Abstracts, Vienna, Austria, 23–28 April 2017; p. 17129. [Google Scholar]
- Acevedo, E.; Silva, P.; Silva, H. Wheat growth and physiology. Bread Wheat Improv. Prod. 2002, 30, 39–70. [Google Scholar]
- Hergert, G.W.; Shaver, T.M. Fertilizing Winter Wheat; UNL-W Est Central Research and Extension Center: North Plate, NE, USA, 2009; p. 69101. [Google Scholar]
- Simionescu, M.; Bilan, Y.; Gędek, S.; Streimikiene, D. The effects of greenhouse gas emissions on cereal production in the European Union. Sustainability 2019, 11, 3433. [Google Scholar] [CrossRef]
- Brunetière, J.-R.; Alexandre, S.; d’Aubreby, M.; Debiesse, G.; Guérin, A.-J.; Perret, B.; Schwartz, D. Le Facteur 4 en France: La Division par 4 des Émissions de Gaz à Effet de Serre à l’Horizon 2050; Rapport final. Technical Report; Conseil général de l’Environnement et du Développement durable: Puteaux, France, 2009. [Google Scholar]
- Stavi, I.; Lal, R. Agriculture and greenhouse gases, a common tragedy. A review. Agron. Sustain. Dev. 2013, 33, 275–289. [Google Scholar] [CrossRef]
- Gan, H.; Liu, J.; Huang, W.; Yang, Y.; Jiang, Y.; Cai, Y. Estimation of agricultural greenhouse gas emissions from 1987 to 2015 in Fujian Province, China. In IOP Conference Series: Earth and Environmental Science, Proceedings of the Fourth International Conference on Energy Engineering and Environmental Protection; Xiamen, China, 19–21 November 2019, IOP Publishing: Bristol, UK, 2020; Volume 467, p. 012158. [Google Scholar]
- Alhassan, A.-R.M.; Yang, C.; Ma, W.; Li, G. Influence of conservation tillage on Greenhouse gas fluxes and crop productivity in spring-wheat agroecosystems on the Loess Plateau of China. PeerJ 2021, 9, e11064. [Google Scholar] [CrossRef] [PubMed]
- Holka, M.; Bieńkowski, J. Carbon Footprint and Life-Cycle Costs of Maize Production in Conventional and Non-Inversion Tillage Systems. Agronomy 2020, 10, 1877. [Google Scholar] [CrossRef]
- Camargo, G.G.; Ryan, M.R.; Richard, T.L. Energy use and greenhouse gas emissions from crop production using the farm energy analysis tool. BioScience 2013, 63, 263–273. [Google Scholar] [CrossRef]
- Ma, B.; Liang, B.C.; Biswas, D.K.; Morrison, M.J.; McLaughlin, N.B. The carbon footprint of maize production as affected by nitrogen fertilizer and maize-legume rotations. Nutr. Cycl. Agroecosyst 2012, 94, 15–31. [Google Scholar] [CrossRef]
- Linquist, B.; Van Groenigen, K.J.; Adviento-Borbe, M.A.; Pittelkow, C.; Van Kessel, C. An agronomic assessment of greenhouse gas emissions from major cereal crops. Glob. Chang. Biol. 2012, 18, 194–209. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Zhang, Y.; McDaniel, M.D.; Sun, L.; Su, W.; Fan, X.; Liu, S.; Xiao, X. Water-saving irrigation is a ‘win-win’ management strategy in rice paddies–With both reduced greenhouse gas emissions and enhanced water use efficiency. Agric. Water Manag. 2020, 228, 105889. [Google Scholar] [CrossRef]
- Adviento-Borbe, M.; Haddix, M.; Binder, D.; Walters, D.; Dobermann, A. Soil greenhouse gas fluxes and global warming potential in four high-yielding maize systems. Glob. Chang. Biol. 2007, 13, 1972–1988. [Google Scholar] [CrossRef]
- Bhatia, A.; Pathak, H.; Aggarwal, P.K.; Jain, N. Trade-off between productivity enhancement and global warming potential of rice and wheat in India. Nutr. Cycl. Agroecosyst. 2010, 86, 413–424. [Google Scholar] [CrossRef]
- Das, S.; Adhya, T.K. Effect of combine application of organic manure and inorganic fertilizer on methane and nitrous oxide emissions from a tropical flooded soil planted to rice. Geoderma 2014, 213, 185–192. [Google Scholar] [CrossRef]
- Dendooven, L.; Patiño-Zúñiga, L.; Verhulst, N.; Luna-Guido, M.; Marsch, R.; Govaerts, B. Global warming potential of agricultural systems with contrasting tillage and residue management in the central highlands of Mexico. Agric. Ecosyst. Environ. 2012, 152, 50–58. [Google Scholar] [CrossRef]
- Jain, N.; Arora, P.; Tomer, R.; Mishra, S.V.; Bhatia, A.; Pathak, H.; Chakraborty, D.; Kumar, V.; Dubey, D.; Harit, R. Greenhouse gases emission from soils under major crops in Northwest India. Sci. Total Environ. 2016, 542, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Laratte, B.; Guillaume, B.; Kim, J.; Birregah, B. Modeling cumulative effects in life cycle assessment: The case of fertilizer in wheat production contributing to the global warming potential. Sci. Total Environ. 2014, 481, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Yang, X.; Gao, C.; Wu, P.; Liu, J.; Xu, Y.; Shen, Q.; Zou, J.; Guo, S. Net annual global warming potential and greenhouse gas intensity in Chinese double rice-cropping systems: A 3-year field measurement in long-term fertilizer experiments. Glob. Chang. Biol. 2011, 17, 2196–2210. [Google Scholar] [CrossRef]
- Tuomisto, H.; Hodge, I.; Riordan, P.; Macdonald, D. Comparing global warming potential, energy use and land use of organic, conventional and integrated winter wheat production. Ann. Appl. Biol. 2012, 161, 116–126. [Google Scholar] [CrossRef]
- Stocker, T. Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Fatima, Z.; Ahmed, M.; Hussain, M.; Abbas, G.; Ul-Allah, S.; Ahmad, S.; Ahmed, N.; Ali, M.A.; Sarwar, G.; ul Haque, E. The fingerprints of climate warming on cereal crops phenology and adaptation options. Sci. Rep. 2020, 10, 18013. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2016, 6, 166–171. [Google Scholar] [CrossRef]
- Feng, Q.; Ma, H.; Jiang, X.; Wang, X.; Cao, S. What has caused desertification in China? Sci. Rep. 2015, 5, 15998. [Google Scholar] [CrossRef]
- Bevacqua, E.; Vousdoukas, M.I.; Zappa, G.; Hodges, K.; Shepherd, T.G.; Maraun, D.; Mentaschi, L.; Feyen, L. More meteorological events that drive compound coastal flooding are projected under climate change. Commun. Earth Environ. 2020, 1, 47. [Google Scholar] [CrossRef]
- Corwin, D.L. Climate change impacts on soil salinity in agricultural areas. Eur. J. Soil Sci. 2021, 72, 842–862. [Google Scholar] [CrossRef]
- FAOSTAT. Crops. Food and Agriculture Organization of the United Nations. 2021. Available online: http://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 19 July 2021).
- Mahmood, N.; Ahmad, B.; Hassan, S.; Bakhsh, K. Impact of temperature ADN precipitation on rice productivity in rice-wheat cropping system of Punjab province. J. Anim. Plant Sci. 2012, 22, 993–997. [Google Scholar]
- Janjua, P.Z.; Samad, G.; Khan, N.U.; Nasir, M. Impact of climate change on wheat production: A case study of Pakistan [with comments]. Pak. Dev. Rev. 2010, 49, 799–822. [Google Scholar] [CrossRef]
- Boonwichai, S.; Shrestha, S.; Babel, M.S.; Weesakul, S.; Datta, A. Evaluation of climate change impacts and adaptation strategies on rainfed rice production in Songkhram River Basin, Thailand. Sci. Total Environ. 2019, 652, 189–201. [Google Scholar] [CrossRef]
- Castro, H.F.; Classen, A.T.; Austin, E.E.; Norby, R.J.; Schadt, C.W. Soil microbial community responses to multiple experimental climate change drivers. Appl. Environ. Microbiol. 2010, 76, 999–1007. [Google Scholar] [CrossRef]
- Muehe, E.M.; Wang, T.; Kerl, C.F.; Planer-Friedrich, B.; Fendorf, S. Rice production threatened by coupled stresses of climate and soil arsenic. Nat. Commun. 2019, 10, 4985. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.; Ruedy, R.; Sato, M.; Lo, K. Global surface temperature change. Rev. Geophys. 2010, 48. [Google Scholar] [CrossRef]
- Matthews, R.; Kropff, M.; Horie, T.; Bachelet, D. Simulating the impact of climate change on rice production in Asia and evaluating options for adaptation. Agric. Syst. 1997, 54, 399–425. [Google Scholar] [CrossRef]
- Li, X.; Takahashi, T.; Suzuki, N.; Kaiser, H.M. Impact of climate change on maize production in Northeast and Southwest China and risk mitigation strategies. Apcbee Procedia 2014, 8, 11–20. [Google Scholar] [CrossRef][Green Version]
- Leng, G.; Huang, M. Crop yield response to climate change varies with crop spatial distribution pattern. Sci. Rep. 2017, 7, 1463. [Google Scholar] [CrossRef] [PubMed]
- Nagy, J. Effect of irrigation on maize yield (Zea mays L.). Acta Agrar. Debr. 2003, 30–35. [Google Scholar] [CrossRef]
- Lobell, D.B.; Burke, M.B. On the use of statistical models to predict crop yield responses to climate change. Agric. For. Meteorol. 2010, 150, 1443–1452. [Google Scholar] [CrossRef]
- Lobell, D.B.; Bänziger, M.; Magorokosho, C.; Vivek, B. Nonlinear heat effects on African maize as evidenced by historical yield trials. Nat. Clim. Chang. 2011, 1, 42–45. [Google Scholar] [CrossRef]
- Herrero, M.P.; Johnson, R. High temperature stress and pollen viability of maize 1. Crop Sci. 1980, 20, 796–800. [Google Scholar] [CrossRef]
- Lobell, D.B.; Gourdji, S.M. The influence of climate change on global crop productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- de Fraiture, C.; Smakhtin, V.; Bossio, D.; McCornick, P.; Hoanh, C.; Noble, A.; Molden, D.; Gichuki, F.; Giordano, M.; Finlayson, M. Facing climate change by securing water for food, livelihoods and ecosystems. J. Semi-Arid. Trop. Agric. Res. 2007, 4, 1–21. [Google Scholar]
- Harkness, C.; Semenov, M.A.; Areal, F.; Senapati, N.; Trnka, M.; Balek, J.; Bishop, J. Adverse weather conditions for UK wheat production under climate change. Agric. For. Meteorol. 2020, 282, 107862. [Google Scholar] [CrossRef] [PubMed]
- DEFRA. Farming Statistics Provisional Crop. Areas, Yields and Livestock Populations at June 2018—United Kingdom; DEFRA: London, UK, 2018.
- FAOSTAT F. Available online: http://www.fao.org/faostat/en/#home (accessed on 30 June 2018).
- Hernandez-Ochoa, I.M.; Asseng, S.; Kassie, B.T.; Xiong, W.; Robertson, R.; Pequeno, D.N.L.; Sonder, K.; Reynolds, M.; Babar, M.A.; Milan, A.M. Climate change impact on Mexico wheat production. Agric. For. Meteorol. 2018, 263, 373–387. [Google Scholar] [CrossRef]
- Demirhan, H. Impact of increasing temperature anomalies and carbon dioxide emissions on wheat production. Sci. Total Environ. 2020, 741, 139616. [Google Scholar] [CrossRef]
- Tao, F.; Zhang, Z.; Xiao, D.; Zhang, S.; Rötter, R.P.; Shi, W.; Liu, Y.; Wang, M.; Liu, F.; Zhang, H. Responses of wheat growth and yield to climate change in different climate zones of China, 1981–2009. Agric. For. Meteorol. 2014, 189, 91–104. [Google Scholar] [CrossRef]
- Zewdie, A. Impacts of climate change on food security: A literature review in Sub Saharan Africa. J. Earth Sci. Clim. Chang. 2014, 5, 225. [Google Scholar]
- Li, M. Climate Change to Adversely Impact Grain Production in China by 2030; IFPRI: Washington, DC, USA, 2018. [Google Scholar]
- Gammans, M.; Mérel, P.; Ortiz-Bobea, A. Negative impacts of climate change on cereal yields: Statistical evidence from France. Environ. Res. Lett. 2017, 12, 054007. [Google Scholar] [CrossRef]
- Trostle, R. Global Agricultural Supply and Demand: Factors Contributing to the Recent Increase. Food Commodity Prices Outlook Report WRS-0801, ERS; USDA: Washington, DC, USA, 2008.
- Conforti, P. Looking Ahead in World Food and Agriculture: Perspectives to 2050; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- Wheeler, T.; Von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Sage, R.F.; Way, D.A.; Kubien, D.S. Rubisco, Rubisco activase, and global climate change. J. Exp. Bot. 2008, 59, 1581–1595. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol 2009, 11, 100–105. [Google Scholar]
- Chaves, M.; Oliveira, M. Mechanisms underlying plant resilience to water deficits: Prospects for water-saving agriculture. J. Exp. Bot. 2004, 55, 2365–2384. [Google Scholar] [CrossRef]
- Chaves, M.; Flexas, J.; Gulías, J.; Loreto, F.; Medrano, H. Photosyntesis under water deficits, flooding and salinity. Terr. Photosyntesis Chang. Environ. 2012, 299–311. [Google Scholar]
- Chaves, M.M.; Costa, J.M.; Saibo, N.J.M. Recent advances in photosynthesis under drought and salinity. Adv. Bot. Res. 2011, 57, 49–104. [Google Scholar]
- Gray, J.E.; Hetherington, A.M. Plant development: YODA the stomatal switch. Curr. Biol. 2004, 14, R488–R490. [Google Scholar] [CrossRef]
- Assmann, S.M. Signal transduction in guard cells. Annu. Rev. Cell Biol. 1993, 9, 345–375. [Google Scholar] [CrossRef]
- Cornic, G.; Massacci, A. Leaf photosynthesis under drought stress. In Photosynthesis and the Environment; Springer: Berlin/Heidelberg, Germany, 1996; pp. 347–366. [Google Scholar]
- Zargar, S.M.; Gupta, N.; Nazir, M.; Mahajan, R.; Malik, F.A.; Sofi, N.R.; Shikari, A.B.; Salgotra, R. Impact of drought on photosynthesis: Molecular perspective. Plant Gene 2017, 11, 154–159. [Google Scholar] [CrossRef]
- Pandey, V.; Shukla, A. Acclimation and Tolerance Strategies of Rice under Drought Stress. Rice Sci. 2015, 22, 147–161. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef]
- Allahverdiyev, T. Impact of soil water deficit on some physiological parameters of durum and bread wheat genotypes. Poljopr. I Sumar. 2016, 62, 131. [Google Scholar] [CrossRef]
- Elazab, A.; Serret, M.D.; Araus, J.L. Interactive effect of water and nitrogen regimes on plant growth, root traits and water status of old and modern durum wheat genotypes. Planta 2016, 244, 125–144. [Google Scholar] [CrossRef]
- Shangguan, Z.; Shao, M.; Dyckmans, J. Effects of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat. J. Plant Physiol. 2000, 156, 46–51. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Saifullah, Ahmad, A. Water stress and nitrogen management effects on gas exchange, water relations, and water use efficiency in wheat. J. Plant Nutr. 2011, 34, 1867–1882. [Google Scholar] [CrossRef]
- Zhao, L.; Xiao, H.; Liu, X. Relationships between carbon isotope discrimination and yield of spring wheat under different water and nitrogen levels. J. Plant Nutr. 2007, 30, 947–963. [Google Scholar] [CrossRef]
- Kamara, A.; Menkir, A.; Badu-Apraku, B.; Ibikunle, O. The influence of drought stress on growth, yield and yield components of selected maize genotypes. J. Agric. Sci. 2003, 141, 43–50. [Google Scholar] [CrossRef]
- Hura, T.; Hura, K.; Grzesiak, M.; Rzepka, A. Effect of long-term drought stress on leaf gas exchange and fluorescence parameters in C 3 and C 4 plants. Acta Physiol. Plant. 2007, 29, 103–113. [Google Scholar] [CrossRef]
- Wilkinson, S.; Kudoyarova, G.R.; Veselov, D.S.; Arkhipova, T.N.; Davies, W.J. Plant hormone interactions: Innovative targets for crop breeding and management. J. Exp. Bot. 2012, 63, 3499–3509. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Bai, L.; Chang, J. Song C-p: Chloroplast protein PLGG1 is involved in abscisic acid-regulated lateral root development and stomatal movement in Arabidopsis. Biochem. Biophys. Res. Commun. 2018, 495, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Dong, B.; Shiran, B.; Talbot, M.J.; Edlington, J.E.; Hughes, T.; White, R.G.; Gubler, F.; Dolferus, R. Control of abscisic acid catabolism and abscisic acid homeostasis is important for reproductive stage stress tolerance in cereals. Plant Physiol. 2011, 156, 647–662. [Google Scholar] [CrossRef] [PubMed]
- Sperotto, R.A.; Ricachenevsky, F.K.; Duarte, G.L.; Boff, T.; Lopes, K.L.; Sperb, E.R.; Grusak, M.A.; Fett, J.P. Identification of up-regulated genes in flag leaves during rice grain filling and characterization of Os NAC5, a new ABA-dependent transcription factor. Planta 2009, 230, 985–1002. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Lv, B.; Li, J.; Luo, L.; Lu, S.; Zhang, X.; Ma, H.; Ming, F. The NAC family transcription factor OsNAP confers abiotic stress response through the ABA pathway. Plant Cell Physiol. 2014, 55, 604–619. [Google Scholar] [CrossRef]
- Liang, C.; Wang, Y.; Zhu, Y.; Tang, J.; Hu, B.; Liu, L.; Ou, S.; Wu, H.; Sun, X.; Chu, J. OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 10013–10018. [Google Scholar] [CrossRef]
- Du, H.; Wang, N.; Cui, F.; Li, X.; Xiao, J.; Xiong, L. Characterization of the β-carotene hydroxylase gene DSM2 conferring drought and oxidative stress resistance by increasing xanthophylls and abscisic acid synthesis in rice. Plant Physiol. 2010, 154, 1304–1318. [Google Scholar] [CrossRef]
- Kuromori, T.; Seo, M.; Shinozaki, K. ABA transport and plant water stress responses. Trends Plant Sci. 2018, 23, 513–522. [Google Scholar] [CrossRef]
- Peleg, Z.; Reguera, M.; Tumimbang, E.; Walia, H.; Blumwald, E. Cytokinin-mediated source/sink modifications improve drought tolerance and increase grain yield in rice under water-stress. Plant Biotechnol. J. 2011, 9, 747–758. [Google Scholar] [CrossRef]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef]
- Wang, C.; Yang, A.; Yin, H.; Zhang, J. Influence of water stress on endogenous hormone contents and cell damage of maize seedlings. J. Integr. Plant Biol. 2008, 50, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Van den Broeck, L.; Inzé, D. The pivotal role of ethylene in plant growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5. [Google Scholar] [CrossRef]
- Gomez-Roldan, V.; Fermas, S.; Brewer, P.B.; Puech-Pagès, V.; Dun, E.A.; Pillot, J.-P.; Letisse, F.; Matusova, R.; Danoun, S.; Portais, J.-C. Strigolactone inhibition of shoot branching. Nature 2008, 455, 189–194. [Google Scholar] [CrossRef]
- Xue-Xuan, X.; Hong-Bo, S.; Yuan-Yuan, M.; Gang, X.; Jun-Na, S.; Dong-Gang, G.; Cheng-Jiang, R. Biotechnological implications from abscisic acid (ABA) roles in cold stress and leaf senescence as an important signal for improving plant sustainable survival under abiotic-stressed conditions. Crit. Rev. Biotechnol. 2010, 30, 222–230. [Google Scholar] [CrossRef]
- Argueso, C.T.; Ferreira, F.J.; Kieber, J.J. Environmental perception avenues: The interaction of cytokinin and environmental response pathways. Plant Cell Environ. 2009, 32, 1147–1160. [Google Scholar] [CrossRef]
- Divi, U.K.; Krishna, P. Brassinosteroid: A biotechnological target for enhancing crop yield and stress tolerance. New Biotechnol. 2009, 26, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Fleury, D.; Jefferies, S.; Kuchel, H.; Langridge, P. Genetic and genomic tools to improve drought tolerance in wheat. J. Exp. Bot. 2010, 61, 3211–3222. [Google Scholar] [CrossRef]
- Khan, M.A. Current status of genomic based approaches to enhance drought tolerance in rice (Oryza sativa L.), an overview. Mol. Plant Breed. 2012, 3. [Google Scholar]
- Blum, A. Plant Breeding for Stress Environments; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Raza, A.; Mehmood, S.S.; Ashraf, F.; Khan, R.S.A. Genetic diversity analysis of Brassica species using PCR-based SSR markers. Gesunde Pflanz. 2019, 71, 1–7. [Google Scholar] [CrossRef]
- Raza, A.; Shaukat, H.; Ali, Q.; Habib, M. Assessment of RAPD markers to analyse the genetic diversity among sunflower (Helianthus annuus L.) genotypes. Turk. J. Agric.-Food Sci. Technol. 2018, 6, 107–111. [Google Scholar] [CrossRef]
- Lopes, M.S.; El-Basyoni, I.; Baenziger, P.S.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A. Exploiting genetic diversity from landraces in wheat breeding for adaptation to climate change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D. Abiotic Stress Adaptation in Plants: Physiological, Molecular and Genomic Foundation. Ann. Bot. 2011, 107, vii. [Google Scholar] [CrossRef]
- Shivhare, R.; Lata, C. Assessment of pearl millet genotypes for drought stress tolerance at early and late seedling stages. Acta Physiol. Plant. 2019, 41, 39. [Google Scholar] [CrossRef]
- Bhatnagar-Mathur, P.; Vadez, V.; Sharma, K.K. Transgenic approaches for abiotic stress tolerance in plants: Retrospect and prospects. Plant Cell Rep. 2008, 27, 411–424. [Google Scholar] [CrossRef]
- Lata, C.; Prasad, M. Role of DREBs in regulation of abiotic stress responses in plants. J. Exp. Bot. 2011, 62, 4731–4748. [Google Scholar] [CrossRef]
- Yadav, D.; Shavrukov, Y.; Bazanova, N.; Chirkova, L.; Borisjuk, N.; Kovalchuk, N.; Ismagul, A.; Parent, B.; Langridge, P.; Hrmova, M. Constitutive overexpression of the TaNF-YB4 gene in transgenic wheat significantly improves grain yield. J. Exp. Bot. 2015, 66, 6635–6650. [Google Scholar] [CrossRef] [PubMed]
- Henry, R.J. Genomics of grain quality in cereals. Crop Breed. Appl. Biotechnol. 2021, 21. [Google Scholar] [CrossRef]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M.; Zheng, T.; Fuentes, R.R.; Zhang, F. Genomic variation in 3010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Zhao, H.; Ren, L.; Song, W.; Zeng, B.; Guo, J.; Wang, B.; Liu, Z.; Chen, J.; Li, W. Genome-wide genetic changes during modern breeding of maize. Nat. Genet. 2012, 44, 812–815. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Shi, C.; Thudi, M.; Mariac, C.; Wallace, J.; Qi, P.; Zhang, H.; Zhao, Y.; Wang, X.; Rathore, A. Pearl millet genome sequence provides a resource to improve agronomic traits in arid environments. Nat. Biotechnol. 2017, 35, 969–976. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, Y.; Li, C.; Wang, A.; Zhao, Q.; Li, W.; Guo, Y.; Deng, L.; Zhu, C.; Fan, D. Genome-wide association study of flowering time and grain yield traits in a worldwide collection of rice germplasm. Nat. Genet. 2012, 44, 32–39. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Singh, R.B.; Pujarula, V.L.; Bollam, S.; Pusuluri, M.; Chellapilla, T.S.; Yadav, R.S.; Gupta, R. Genome-wide association studies and genomic selection in Pearl Millet: Advances and prospects. Front. Genet. 2020, 10, 1389. [Google Scholar] [CrossRef]
- Pham, A.-T.; Maurer, A.; Pillen, K.; Brien, C.; Dowling, K.; Berger, B.; Eglinton, J.K.; March, T.J. Genome-wide association of barley plant growth under drought stress using a nested association mapping population. BMC Plant Biol. 2019, 19, 134. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Huang, X.; Zhi, H.; Zhao, Y.; Zhao, Q.; Li, W.; Chai, Y.; Yang, L.; Liu, K.; Lu, H. A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet (Setaria italica). Nat. Genet. 2013, 45, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.P.; Ramu, P.; Deshpande, S.P.; Hash, C.T.; Shah, T.; Upadhyaya, H.D.; Riera-Lizarazu, O.; Brown, P.J.; Acharya, C.B.; Mitchell, S.E. Population genomic and genome-wide association studies of agroclimatic traits in sorghum. Proc. Natl. Acad. Sci. USA 2013, 110, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Bohra, A.; Chand Jha, U.; Godwin, I.D.; Kumar Varshney, R. Genomic interventions for sustainable agriculture. Plant Biotechnol. J. 2020, 18, 2388–2405. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Bashir, T.; Hashem, A.; Abd_Allah, E.F. Systems biology approach in plant abiotic stresses. Plant Physiol. Biochem. 2017, 121, 58–73. [Google Scholar] [CrossRef]
- Singh, R.K.; Prasad, A.; Muthamilarasan, M.; Parida, S.K.; Prasad, M. Breeding and biotechnological interventions for trait improvement: Status and prospects. Planta 2020, 252, 54. [Google Scholar] [CrossRef]
- Choudhary, M.; Singh, V.; Muthusamy, V.; Wani, S.H. Harnessing crop wild relatives for crop improvement. Indian J. 2017, 6, 73–85. [Google Scholar] [CrossRef]
- Sharma, D.; Khulbe, R.K.; Pal, R.S.; Bettanaika, J.; Kant, L. Wild Progenitor and Landraces Led Genetic Gain in the Modern-Day Maize (Zea mays L.). In Landraces-Traditional Variety and Natural Breed; IntechOpen: London, UK, 2021. [Google Scholar]
- Ananda, G.K.; Myrans, H.; Norton, S.L.; Gleadow, R.; Furtado, A.; Henry, R.J. Wild sorghum as a promising resource for crop improvement. Front. Plant Sci. 2020, 11, 1108. [Google Scholar] [CrossRef]
- Kole, C.; Muthamilarasan, M.; Henry, R.; Edwards, D.; Sharma, R.; Abberton, M.; Batley, J.; Bentley, A.; Blakeney, M.; Bryant, J. Application of genomics-assisted breeding for generation of climate resilient crops: Progress and prospects. Front. Plant Sci. 2015, 6, 563. [Google Scholar] [CrossRef]
- Lata, C.; Gupta, S.; Prasad, M. Foxtail millet: A model crop for genetic and genomic studies in bioenergy grasses. Crit. Rev. Biotechnol. 2013, 33, 328–343. [Google Scholar] [CrossRef]
- Lata, C.; Shivhare, R. Engineering cereal crops for enhanced abiotic stress tolerance. In Proceedings of the Indian National Science Academy, New Delhi, Indian, 4 December 2021; Volume 87, pp. 63–83. [Google Scholar]
- Olmstead, S.M. Climate change adaptation and water resource management: A review of the literature. Energy Econ. 2014, 46, 500–509. [Google Scholar] [CrossRef]
- Surendran, U.; Raja, P.; Jayakumar, M.; Ramasubramoniam, S. Use of efficient water saving techniques for production of rice in India under climate change scenario: A critical review. J. Clean. Prod. 2021, 309, 127272. [Google Scholar] [CrossRef]
- Cooley, H.; Christian-Smith, J.; Gleick, P.H. More with Less: Agricultural Water Conservation and Efficiency in California. Oakl. Calif. 2008, 30, 2011. [Google Scholar]
- Lampayan, R.M.; Rejesus, R.M.; Singleton, G.R.; Bouman, B.A. Adoption and economics of alternate wetting and drying water management for irrigated lowland rice. Field Crops Res. 2015, 170, 95–108. [Google Scholar] [CrossRef]
- Dawe, D. Increasing water productivity in rice-based systems in Asia–past trends, current problems, and future prospects. Plant Prod. Sci. 2005, 8, 221–230. [Google Scholar] [CrossRef]
- Brouwer, C.; Prins, K.; Kay, M.; Heibloem, M. Irrigation Water Management: Irrigation Methods Training Manual No. 5; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Zhu, G.H.; Zhuang, C.X.; Wang, Y.Q.; Jiang, L.R.; Peng, X.X. Differential expression of rice genes under different nitrogen forms and their relationship with sulfur metabolism. J. Integr. Plant Biol. 2006, 48, 1177–1184. [Google Scholar] [CrossRef]
- Fageria, N.; Dos Santos, A.; Moraes, M. Influence of urea and ammonium sulfate on soil acidity indices in lowland rice production. Commun. Soil Sci. Plant Anal. 2010, 41, 1565–1575. [Google Scholar] [CrossRef]
- Ju, X.T.; Kou, C.L.; Christie, P.; Dou, Z.; Zhang, F. Changes in the soil environment from excessive application of fertilizers and manures to two contrasting intensive cropping systems on the North China Plain. Environ. Pollut. 2007, 145, 497–506. [Google Scholar] [CrossRef]
- Liu, T.; Ren, T.; White, P.J.; Cong, R.; Lu, J. Storage nitrogen co-ordinates leaf expansion and photosynthetic capacity in winter oilseed rape. J. Exp. Bot. 2018, 69, 2995–3007. [Google Scholar] [CrossRef]
- Alexander, A.; Singh, V.K.; Mishra, A. Overexpression of differentially expressed AhCytb6 gene during plant-microbe interaction improves tolerance to N2 deficit and salt stress in transgenic tobacco. Sci. Rep. 2021, 11, 13435. [Google Scholar] [CrossRef] [PubMed]
- Ghatak, A.; Chaturvedi, P.; Bachmann, G.; Valledor, L.; Ramšak, Ž; Bazargani, M.M.; Bajaj, P.; Jegadeesan, S.; Li, W.; Sun, X. Physiological and proteomic signatures reveal mechanisms of superior drought resilience in pearl millet compared to wheat. Front. Plant Sci. 2021, 11, 600278. [Google Scholar] [CrossRef]
- Kumar, A.; Tomer, V.; Kaur, A.; Kumar, V.; Gupta, K. Millets: A solution to agrarian and nutritional challenges. Agric. Food Secur. 2018, 7, 31. [Google Scholar] [CrossRef]
- Li, P.; Brutnell, T.P. Setaria viridis and Setaria italica, model genetic systems for the Panicoid grasses. J. Exp. Bot. 2011, 62, 3031–3037. [Google Scholar] [CrossRef]
- Habiyaremye, C.; Matanguihan, J.B.; D’Alpoim Guedes, J.; Ganjyal, G.M.; Whiteman, M.R.; Kidwell, K.K.; Murphy, K.M. Proso millet (Panicum miliaceum L.) and its potential for cultivation in the Pacific Northwest, US. A review. Front. Plant Sci. 2017, 7, 1961. [Google Scholar] [CrossRef] [PubMed]
- Aubry, S.; Brown, N.J.; Hibberd, J.M. The role of proteins in C3 plants prior to their recruitment into the C4 pathway. J. Exp. Bot. 2011, 62, 3049–3059. [Google Scholar] [CrossRef]
- Sage, R.; Christin, P.; Edwards, E. The lineages of C4 photosynthesis on planet Earth. J. Exp. Bot. 2011, 62, 3155–3169. [Google Scholar] [CrossRef] [PubMed]
- Lychuk, T.E.; Hill, R.L.; Izaurralde, R.C.; Momen, B.; Thomson, A.M. Evaluation of climate change impacts and effectiveness of adaptation options on crop yield in the Southeastern United States. Field Crops Res. 2017, 214, 228–238. [Google Scholar] [CrossRef]
- Sage, R.F.; Zhu, X.-G. Exploiting the engine of C4 photosynthesis. J. Exp. Bot. 2011, 62, 2989–3000. [Google Scholar] [CrossRef] [PubMed]
- Lychuk, T.; Moulin, A.; Lemke, R.; Gossen, B.; Leeson, J.; Kirk, A.; Johnson, E.; Olfert, O.; Brandt, S.; Thomas, A. Effects of crop inputs, diversity, environment, and terrain on yield in an 18-yr study in the semi-arid Canadian Prairies. Can. J. Plant Sci. 2017, 97, 715–730. [Google Scholar]
- Bandyopadhyay, T.; Muthamilarasan, M.; Prasad, M. Millets for next generation climate-smart agriculture. Front. Plant Sci. 2017, 8, 1266. [Google Scholar] [CrossRef] [PubMed]

| Crops | Area (Million Hectare) | Production (Million Tons) | ||||
|---|---|---|---|---|---|---|
| 2019/20 | 2020/21 (Pre.) | 2021/22 (Pro.) | 2019/20 | 2020/21 (Pre.) | 2021/22 (Pro.) | |
| Rice | 160.39 | 162.56 | 162.90 | 497.74 | 504.94 | 506.04 |
| Maize | 194.05 | 197.28 | 199.64 | 1117.56 | 1120.65 | 1194.80 |
| Wheat | 216.20 | 221.86 | 224.49 | 763.49 | 775.82 | 792.40 |
| Crop | Global Warming Potential (kg CO2 eq. ha−1) | Carbon Equivalent Emission (kg C ha−1) |
|---|---|---|
| Rice | 2890–17,000 | 956–4600 |
| Wheat | 2000–18,000 | 545–4900 |
| Maize | 3427–17,600 | 935–4800 |
| Millet | 3218 | 878 |
| Rice–Wheat * | 7137–18,000 | 2000–4900 |
| Wheat–Maize * | 12,880–18,850 | 3512–5100 |
| Crop | Findings | References |
|---|---|---|
| Rice | Increase in leaf rolling, biomass and root traits severely affected, decrease in elongation and expansion growth, and number of tillers as well as physiological traits, i.e., photosynthesis, transpiration, leaf area index, and water use efficiency | [127] |
| Wheat | Exposure to drought at anthesis reduces fertility by increasing pollen sterility, number of tillers and kernels per ear, and ultimately reduced yield | [128] |
| Decreased photosynthesis rate, transpiration rate, stomatal conductance, mesophyll conductance, photosynthetic pigment content, leaf area, dry weight, and relative water content | [129] | |
| Well water conditions lead to an increase in aerial biomass, root dry biomass, and root length. However, water stress studies found a negative correlation between aerial biomass and root dry biomass, root length, and root weight density | [130] | |
| Reduced photochemical quenching, the efficiency of PSII, and potential photosynthetic quantum conversion of leaves | [131] | |
| Decreased rate of photosynthetic gas exchange parameters, leaf water potential, and osmotic potential | [132] | |
| Increased ground dry matter and grain yield under well water conditions. However, foliar and grain carbon isotope discrimination decreased upon stress | [133] | |
| Maize | Decreased plant height, stem diameter, leaf area, number of leaves per plant, cob length, and shoot fresh and dry weight per plant. Total biomass accumulation at silking, grain filling, and maturity, reduced by 37, 34, and 21%, respectively | [134] |
| Triticale, field bean, maize, and amaranth | Field bean and maize acclimatized more effectively compared to triticale and amaranth, due to the synthesis of phenolic compounds that act as photo protectors to avoid damage to PSII | [135] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neupane, D.; Adhikari, P.; Bhattarai, D.; Rana, B.; Ahmed, Z.; Sharma, U.; Adhikari, D. Does Climate Change Affect the Yield of the Top Three Cereals and Food Security in the World? Earth 2022, 3, 45-71. https://doi.org/10.3390/earth3010004
Neupane D, Adhikari P, Bhattarai D, Rana B, Ahmed Z, Sharma U, Adhikari D. Does Climate Change Affect the Yield of the Top Three Cereals and Food Security in the World? Earth. 2022; 3(1):45-71. https://doi.org/10.3390/earth3010004
Chicago/Turabian StyleNeupane, Dhurba, Pramila Adhikari, Dwarika Bhattarai, Birendra Rana, Zeeshan Ahmed, Umanath Sharma, and Debraj Adhikari. 2022. "Does Climate Change Affect the Yield of the Top Three Cereals and Food Security in the World?" Earth 3, no. 1: 45-71. https://doi.org/10.3390/earth3010004
APA StyleNeupane, D., Adhikari, P., Bhattarai, D., Rana, B., Ahmed, Z., Sharma, U., & Adhikari, D. (2022). Does Climate Change Affect the Yield of the Top Three Cereals and Food Security in the World? Earth, 3(1), 45-71. https://doi.org/10.3390/earth3010004