Abstract
The activation of hepatic stellate cells (HSCs) is the key event of hepatic fibrosis. Furthermore, activated HSCs also play an important role in the progression of hepatocellular cancer (HCC). Bone morphogenetic protein 14 (BMP14) is a member of the TGF-β/BMP superfamily. So far, most studies have analyzed BMP14 in the context of bone and cartilage formation and homeostasis. The aim of this study was to assess the expression and function of BMP14 in liver fibrosis and HCC. The BMP14 expression increased during the in vitro activation of primary human HSCs and also in mouse models of liver fibrosis. In human HCC, as well as non-tumorous liver tissues, there was a significant correlation between the expression of BMP14 and alpha-smooth-muscle actin (α-SMA), an established marker for HSC activation. RNAi-mediated BMP14 suppression in activated HSCs resulted in the reduced expression of the transcription factors inhibitor of differentiation 1 (ID1) and ID2, known targets of BMP signaling. Interestingly, α-SMA and collagen expression was also reduced in BMP14-depleted cells, while treatment with recombinant BMP14 induced ID1, ID2, α-SMA and collagen expression. In human HCC cell lines, treatment with recombinant BMP14 induced proliferation, migratory activity and colony formation. In summary, our data indicate activated HSCs as a major cellular source of enhanced BMP14 expression in fibrotic liver disease and HCC, and show that BMP14 exhibits pro-fibrogenic as well as pro-tumorigenic effects. Future analyses will reveal the potential of this soluble growth factor as a therapeutic target or prognostic marker for the progression of fibrosis and HCC in patients with chronic liver disease.
1. Introduction
Chronic liver diseases can lead to liver inflammation (hepatitis) and fibrosis of the hepatic tissue. Generally, fibrosis is a response to tissue injury and can therefore be regarded as a physiological wound-healing reaction. However, if the tissue injury persists, the physiological reaction becomes disturbed and progresses with excessive production of extracellular matrix (ECM) proteins that increasingly accumulate [1,2]. Progressive fibrosis leads to impaired liver function and can ultimately progress to liver cirrhosis with organ failure. Furthermore, liver cirrhosis is the major risk factor for the development of hepatocellular carcinoma (HCC) [1,2]. HCC is one of the most aggressive tumors and the third leading cause of cancer-related deaths worldwide, with incidence still rising [3].
The activation of hepatic stellate cells (HSCs) is considered the key event of hepatic fibrosis [1]. Physiologically, these non-parenchymal liver cells stay quiescent in the space of Disse between the hepatocytes and lining sinusoidal endothelial cells. However, in response to liver injury, HSCs become activated to a myofibroblast-like cell type, and as such activated HSCs are the major cellular source of excessive ECM production in liver fibrosis. Moreover, activated HSCs infiltrate and form the HCC-stroma, and stromal HSCs support HCC progression [2,4,5]. The underlying mechanisms of these tumor-promoting effects are only incompletely understood. However, new insights of how HSC interact with HCC cells might lead to the identification of new therapeutic targets for the treatment of HCC, which are urgently needed. Furthermore, despite increasing knowledge regarding the molecular mechanisms of hepatic fibrosis, no approved therapy exists for liver fibrosis.
Bone morphogenetic proteins (BMPs) are secretory proteins and belong to the transforming growth factor (TGF)-β superfamily. The origin of the name is due to their initial identification as proteins that are involved in bone and cartilage formation. To date, more than 20 different BMP proteins are known, which influence many biological processes including proliferation, differentiation, motility or adhesion during embryogenesis. In the adult organism, BMPs play key roles in the homeostasis of different tissues [6]. A dysregulation of BMP signal transduction has also been described in different pathological processes, including organ fibrosis and different types of cancer [6].
BMP14, also known as growth differentiating factor 5 (GDF5), has so far mostly been studied in the context of cartilage and bone formation and homeostasis [7]. To date, only one study analyzed BMP14 in the context of chronic liver disease and found that the systemic overexpression of BMP14 in adipose tissues reduced high-fat diet-induced fatty liver disease in mice [8]. In addition, only a few studies exist regarding BMP14 and cancer. In oral squamous cell carcinoma, higher BMP14 immunoreactivity has been described in poorly differentiated forms and advanced stages [9]. Furthermore, BMP14 involvement has been suggested during the development of salivary gland neoplasia [9]. Moreover, it has been shown that TGF-ß produced by breast cancer cells induces in endothelial cells the expression of BMP14, which in turn stimulates angiogenesis [10]. The expression and function of BMP14 in HCC is unknown.
The aim of this study was to analyze the expression and role of BMP14 in hepatic fibrosis and cancer.
2. Materials and Methods
2.1. Cells and Cell Culture
The isolation and culture of primary human hepatic stellate cells (HSC) was performed as described [11]. Briefly, cells were isolated by a 2-step EGTA/collagenase perfusion procedure, and sequentially incubated with pronase. Arabinogalactan gradient ultracentrifugation was used to separate HSC from other non-parenchymal liver cells. Only liver tissues of patients that had no known liver disease, histologic evidence for liver fibrosis/inflammation and were classified as non-cancerous by a pathologist were used. The in vitro activation of HSC was achieved via cell culture on uncoated tissue culture dishes [11]. During HSC activation, the medium was changed daily for the first four days in culture, and afterwards every two days. In addition to primary human HSC, the hepatic stellate cell line LX-2 was used [12]. The HCC cell lines HepG2 (ATCC HB-8065), Hep3B (ATCC HB-8064) and PLC (ATCC CRL-8024) were cultured as described [5]. All cells and cell lines were cultured in DMEM (Sigma, Deisenhofen, Germany) supplemented with 10% fetal bovine serum (PAN, Aidenbach, Germany) and 1% penicillin/streptomycin at 37 °C and 8% CO2.
Human liver tissues samples for cell isolation were obtained from the charitable state-controlled foundation HTCR (Human Tissue and Cell Research) with the patient’s informed consent [13]. The sampling and handling of patient material was carried out according to the ethical principles of the Declaration of Helsinki.
For stimulation experiments, cells were serum-starved for at least 12 h before stimulation, and then treated with recombinant human BMP14 (Peprotech, Hamburg, Germany) at a concentration of 100 ng/mL. In experiments with the BMP receptor inhibitor dorsomorphin homologue 1 (DMH1), cells were pre-incubated with 5 nM DMH1 (Sigma-Aldrich, Deisenhofen, Germany) for 15 min. Transfection with BMP14 si-RNA pools was performed as described [14], by using the Lipofectamine RNAiMAX transfection reagent (Life Technologies, Darmstadt, Germany) and si-RNA pools against BMP14 mRNA (siTOOLs Biotech GmbH, Planegg, Germany). Si-RNA pools are complex mixtures of up to 60 different defined siRNAs that are all directed against the target gene, while each of them is applied at a low concentration. This leads to a robust knockdown of the gene of interest, while off-target effects are significantly decreased [15].
2.2. Mouse Models of Liver Fibrosis
Two established mouse models of liver fibrosis were analyzed: chemically induced liver injury by application of thioacetamide (TAA), and cholestasis-induced liver injury by bile duct ligation (BDL). In the TAA model [16], TAA (Sigma, Deisenhofen, Germany) was administered in the drinking water (0.3 g TAA/L) for a duration of 14 weeks. BDL was performed as described in [17]. Briefly, the common bile duct was exposed and ligated twice, and subsequently the abdominal cavity was closed. Control mice were sham-operated. Two weeks after BDL or sham operation the animals were sacrificed. In both models, each group consisted of n = 6–8 mice/group.
All mice were fed with a standard laboratory chow and had access to water ad libitum. The animal studies were approved by the Committee for Animal Health and Care of the local government (54-2532.1-49/13), and conformed to international guidelines on the ethical use of animals.
2.3. Human Tissue Samples
Human liver tissues samples were obtained from the charitable state-controlled foundation HTCR (Human Tissue and Cell Research) with the patient’s informed consent [13]. Sampling and handling of patient material were carried out according to the ethical principles of the Declaration of Helsinki.
2.4. Analysis of mRNA Expression
RNA isolation and reverse transcription were performed as described [17]. Quantitative real-time PCR was performed by applying the LightCycler 480 System (Roche Diagnostics, Mannheim, Germany) as described [17] and the following sets of specific primers: α-SMA human (forward: 5′-CGT GGC TAT TCC TTC GTT AC-3′, reverse: 5′-TGC CAG CAG ACT CCA TCC-3′), β-actin human (forward: 5′-CTA CGT CGC CCT GGA CTT CGA GC-3′, reverse: 5′-GAT GGA GCC GCC GAT CCA CAC GG-3′), β-actin murine (forward: 5′-AGG CCA ACC GTG AAA AGA T-3′, reverse: 5′-GGC GTG AGG GAG AGC ATA-3′), BMP14 human (forward: 5′-TGT ACA GGA CGC TGT CCG A-3′, reverse: 5′-ACA CGT ACC TCT GCT TCC TGA-3′), BMP14 murine (forward: 5′-TTG ACA AAG GGC AAG ATG ACC GA-3′, reverse: 5′-AAC ACC TCC CAG CCA GAT CC-3′), COL1A1 human (forward: 5′-CGG CTC CTG CTC CTC TT-3′, reverse: 5′-GGG GCA GTT CTT GGT CTC-3′), ID1 human (forward: 5′-GTA TCT GCT TCG GGC TTC CA-3′, reverse: 5′-TGA TTC TTG GCG ACT GGC T-3′) and ID2 human (forward: 5′-CAC GGA TAT CAG CAT CCT GTC CT-3′, reverse 5′-CAA GTA AGA GAA CAC CCT GGG AAG A-3′).
2.5. Analysis of Protein Expression
Protein extraction from cells and Western blotting were performed as described [18] by applying the following primary antibodies: rabbit anti-αSMA (ab32575, 1:1000, Abcam, Cambridge, UK), goat anti-BMP14 (AF853, 1:2000, Novus Biologicals, Wiesbaden, Germany), rabbit anti-GAPDH (#2118, 1:1000, Cell Signaling Technology, Danvers, MA, USA) and rabbit anti-ID1 (sc-1331-04, 1:100, Santa Cruz Biotechnology, Heidelberg, Germany). Donkey anti-goat (sc-2304, 1:5000, Santa Cruz Biotechnology, Heidelberg, Germany) and mouse anti-rabbit (sc-2357, 1:10,000, Santa Cruz Biotechnology, Heidelberg, Germany) were used as secondary antibodies.
2.6. Functional in Vitro Analysis
The proliferation of cells was determined using a colorimetric XTT assay (Roche Diagnostics, Mannheim, Germany) according to the manufacturer’s instructions as described in [5]. Briefly, HCC cells were seeded at a density of 4000 cells/well into a 96-well plate and allowed to adhere. Before stimulation, cells were serum-starved for at least 12 h. The following day, cells were treated with 100 ng/mL BMP14 in serum-free DMEM, and allowed to proliferate for 24 h. The supplied XTT was a tetrazolium salt that was converted to a red formazan dye, which is only formed by metabolically active cells and thus correlates with the cell number.
The migratory activity of HCC cells towards BMP14 was quantified using the Cultrex 96 Well Cell Migration assay (Trevigen, Gaithersburg, MD, USA) as described previously [19] and according to the manufacturer’s instructions. Briefly, HCC cells were seeded in serum-free DMEM at a density of 40,000 cells/well into the upper compartment of the provided 96-well plate, containing a polyethylene terephthalate (PET) membrane with a pore size of 8 µm. The lower compartment was filled with serum-free DMEM supplemented with BMP14 (100 ng/mL). Cells were allowed to migrate for 4 h at 37 °C. Migrated cells were detached from the membrane in the lower compartment using the supplied cell dissociation solution. Quantification was achieved by calcein AM, which was internalized and subsequently cleaved to generate free calcein that can be detected fluorometrically.
Clonogenic assays were performed in order to analyze stem cell behavior and attachment-dependent colony formation, as well as the growth of cancer cells, as described previously [20]. Briefly, cells were seeded at a low density (1000 cells/6-well), treated with 100 ng/mL BMP14 and incubated at 37 °C for 12 days. BMP14 was renewed every 2–3 days. Cells were fixed with glutaraldehyde (6% v/v) and simultaneously stained with crystal violet (0.35% w/v). After washing with tap water, plates could be microscopically analyzed. Colony number and size were calculated, applying the CellSens Dimension Software (Olympus Soft Imaging Solutions GmbH, Münster, Germany).
2.7. Histological Analysis
For histochemical and immunofluorescence staining, standard 5 µm sections of formalin-fixed and paraffin-embedded tissue blocks were used.
Immunofluorescence staining was performed by applying the following primary antibodies: rabbit anti-αSMA (ab32575, 1:500, Abcam, Cambridge, UK) and goat anti-BMP14 (AF853, 1:250, Novus Biologicals, Wiesbaden, Germany). Cy3-conjugated donkey anti-rabbit IgG (711-165-152, 1:1000, Jackson ImmunoResearch, Cambridgeshire, UK) and Alexa Fluor 488-conjugated donkey anti-goat IgG (A11055, 1:1000, Invitrogen, Carlsbad, CA, USA) were used as secondary antibodies. Microscopic images were taken, analyzed and processed using an OlympusTM IX83 microscope with the CellSens Dimension Software (Olympus Soft Imaging Solutions GmbH, Münster, Germany).
Sirius red/fast green staining was performed as described previously [17]. Briefly, sections were deparaffinized and rehydrated by a decreasing ethanol gradient. Slices were then immersed in a solution of sirius red (Polysciences, Hirschberg, Germany) and fast green (Sigma, Deisenhofen, Germany; both 1 mg/mL in aqueous solution of picric acid) for 30 min. Sirius red and fast green are used in combination in order to distinguish collagen from its surrounding materials. Sirius red specifically binds to collagen (regardless of type), and fast green stains to all non-collagenous proteins. After dehydration, sections were mounted and microscopically analyzed.
2.8. Statistical Analysis
Calculations were performed by applying the statistical computer package GraphPad Prism Software version 6.01 (GraphPad Software, San Diego, CA, USA). All analyses were performed at least in triplicate. Values are presented as mean ± SEM. Student’s unpaired t-test or, when appropriate, one-way ANOVA were used for comparison between groups, and a p-value < 0.05 was considered statistically significant.
3. Results
3.1. BMP14 Expression in Hepatic Fibrosis and Hepatic Stellate Cells
BMP14 expression was already detectable in quiescent human hepatic stellate cells (HSCs), but the mRNA and protein expression further significantly increased during the course of in vitro activation (Figure 1A). Next, we examined the expression of BMP14 in two experimental models of liver fibrosis in mice, namely, surgically induced cholestasis by bile duct ligation (BDL) [17] and thioacetamide (TAA) [16]. Sirius red/fast green staining showed significant hepatic extracellular matrix deposition in both models (Figure 1B). In both models, we also found a significantly increased expression of BMP14 (Figure 1C). The in silico analysis of 110 human liver tissue samples, applying the GEPIA database [21], showed a significant correlation between the expression of BMP14 and α-smooth muscle actin (α-SMA), a marker for activated HSC [22] (Figure 1D). Furthermore, co-immunofluorescence staining revealed a co-localization of BMP14 and α-SMA signals in human cirrhotic liver tissue (Figure 1E). Together, these data indicate increased BMP14 expression during HSC activation and activated HSC as the cellular source of BMP14 in hepatic fibrosis.
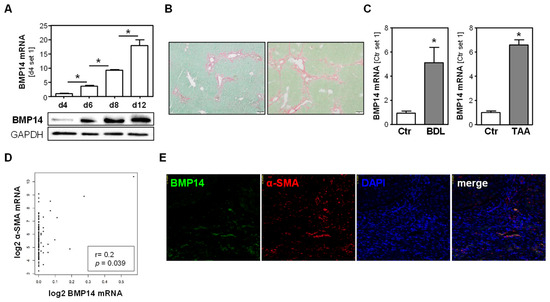
Figure 1.
Expression of BMP14 in hepatic fibrosis. (A) mRNA and protein expression of BMP14 during the course of in vitro activation of primary human hepatic stellate cells (phHSC). (B) Histological sirius red/fast green staining of livers from two different models of hepatic fibrosis (bile duct ligation (BDL; left) and chronic thioacetamide administration (TAA; right); size bar: 100 µm). (C) BMP14 mRNA expression in BDL mice compared to sham-operated controls (Ctr) (left panel), and TAA mice compared to control (Ctr) mice (right panel). (D) Correlation of BMP14 and α-SMA RNA expression levels (log2(TPM)) in n = 110 human liver tissues. “Genotype-Tissue Expression” (GTEx)-derived data were used applying the GEPIA database [21]. (E) Co-immunofluorescence staining of BMP14 (green) and α-SMA (red) in human cirrhotic liver tissue (20× magnification) (*: p < 0.05).
3.2. Effect of BMP14 in Activated Hepatic Stellate Cells
To gain more insight into the role of BMP14 in activated HSCs, we first studied the effect of BMP14 depletion in activated human HSC in vitro. Transfection with si-RNA pools [15] directed against BMP14 led to a significant downregulation of BMP14 mRNA and protein as compared to HSC transfected with control si-RNA pools (Figure 2A,B). In BMP14-depleted cells, we observed a significant downregulation of the expression of the transcription factors’ inhibitor of differentiation 1 (ID1) (Figure 2C,G) and ID2 (Figure 2D), which are both known BMP target genes [23,24]. ID1 has been suggested to be involved in liver injury and fibrogenesis [25], and ID2 has been shown to play a role in HSC differentiation and proliferation [26]. Interestingly, we observed a slight but significant downregulation of α-SMA as well as collagen type I alpha 1 (COL1A1) expression in BMP14-depleted HSC (Figure 2E–G).
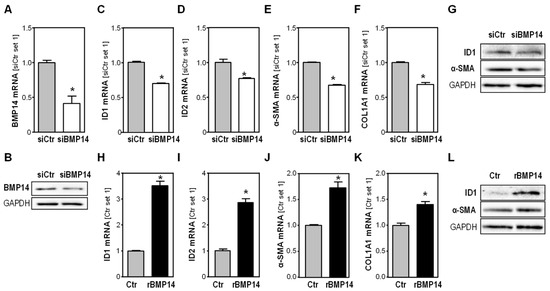
Figure 2.
Effect of BMP14 depletion in activated human hepatic stellate cells (HSC). (A) BMP14 mRNA expression and (B) BMP14 protein expression in activated human HSC transfected with si-RNA pools against BMP14 (siBMP14) or si-control pools (siCtr). Effect of BMP14 depletion in HSC on (C) ID1 mRNA expression, (D) ID2 mRNA expression, (E) α-SMA mRNA expression and (F) COL1A1 mRNA expression. (G) Protein expression of ID1 and α-SMA. Effect of rBMP14 (100 ng/mL) on activated human HSC. (H) ID1 mRNA expression, (I) ID2 mRNA expression, (J) α-SMA mRNA expression and (K) COL1A1 mRNA expression. (L) Protein expression of ID1 and α-SMA (*: p < 0.05).
In a complementary approach, we stimulated activated human HSCs with recombinant BMP14 (rBMP14) and observed a significant induction of ID1 and ID2 expression (Figure 2H,I,L). Furthermore, rBMP14 induced the expression of both α-SMA and COL1A1 in HSCs (Figure 2J–L).
Together, these data indicate that BMP14 in HSCs induces the pro-fibrogenic phenotype of these cells.
3.3. BMP14 Expression in HCC Cells and Tissues and Effect of BMP14 on HCC Cells
In addition to their critical role in hepatic fibrosis, activated HSCs infiltrate and form the HCC-stroma and affect HCC progression [2,4,5]. Interestingly, BMP14 mRNA and protein expression was significantly higher in activated human HSCs compared to different human HCC cell lines (Figure 3A,B). Furthermore, the in silico analysis of 190 human HCC tissue samples, applying the Xena database [27], showed a significant correlation between the expression of BMP14 and α-SMA, indicating that activated HSCs are the major cellular source of BMP14 in HCC (Figure 3C).
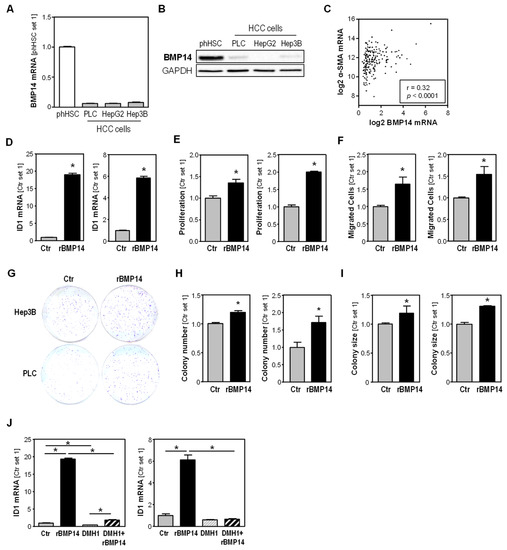
Figure 3.
BMP14 expression in HSC compared to HCC cells, and effects of BMP14 on HCC cells. (A) BMP14 mRNA expression and (B) BMP14 protein expression in primary human HSC compared to HCC cell lines PLC, HepG2 and Hep3B. (C) Correlation of BMP14 and α-SMA mRNA expression levels (log2(TPM)) in n = 190 tumorous liver tissue, applying the TCGA Liver Cancer (LIHC)-derived data and applying the Xena Functional Genomics Explorer database. Effect of recombinant BMP14 (rBMP14) on HCC cell lines Hep3B (left panel) and PLC (right panel). (D) ID1 mRNA expression, (E) cell proliferation and (F) migratory activity towards BMP14 after 4 h. (G–I) Anchorage-dependent clonogenic assay after BMP14 treatment. (G) Representative images. Analysis of (H) colony number and (I) colony size. (J) Effect of the specific BMP receptor inhibitor DMH1 on BMP14-induced ID1 mRNA expression in Hep3B cells (left panel) and PLC cells (right panel). (*: p < 0.05).
To gain insight into the effect of BMP14 on HCC cells, we treated Hep3B and PLC cells with rBMP14 and observed a significant induction of ID1 expression in stimulated cells (Figure 3D). In addition to its pro-fibrotic role, ID1 has been shown to promote HCC progression via inducing growth and metastatic ability [28,29]. In functional in vitro analyses, we found that rBMP14 induced the proliferation (Figure 3E) as well as the migratory activity of HCC cells (Figure 3F). Furthermore, we found that rBMP14 induced colony formation and growth in clonogenic assays (Figure 3G–I).
In order to gain a first insight into the BMP receptor(s) (BMPR) that are involved in the effect of BMP14 on HCC cells, we applied the specific receptor inhibitor dorsomorphin homologue 1 (DMH1) prior to BMP14 stimulation. DMH1 selectively inhibits the type I BMPRs activin receptor-like kinase-2 (ALK2), also known as Activin A receptor, type I (ACVR1) and Alk3 (BMPR1a) [30]. In both HCC cell lines, we observed that the BMP14-induced upregulation of ID1 mRNA expression by BMP14 was nearly completely abrogated by pre-incubation with DMH1 (Figure 3J).
Together, these data indicate that BMP14 secretion contributes to the tumor-promoting effect of HSCs in HCC, and that the BMP14-induced effects in these cells are dependent on Alk3 and/or Alk2 signaling.
4. Discussion
The aim of this study was to gain insights into the expression and role of BMP14 in hepatic fibrosis and cancer. So far, BMP14 has mostly been studied in the context of bone and cartilage formation and homeostasis.
BMP14 exerts an osteogenesis-promoting effect, and is expressed in the limb at sites where the joint cavity is created during development. BMP14 exhibits robust expression up until the fetal day 14 arthroplasty stage, but decreases during later stages of development [31]. BMP14 mutations have been associated with varying types of skeletal dysplasia [32,33,34]. Furthermore, BMP14 appears to have important functions in cartilage remodeling and repair. BMP14 expression in the damaged synovium increases during cartilage repair, and BMP14 showed a potent stimulation of ECM production in cultures of human osteoarthritis chondrocytes. Conversely, reduced BMP14 levels may be a precipitating factor in the development of osteoarthritis [32,35,36]. Moreover, BMP14 deficiencies are associated with delayed fracture repair and impaired bone healing [37].
Here, we show the novel finding that BMP14 is expressed in the liver and, furthermore, we found that BMP14 is upregulated during hepatic fibrosis. As a cellular source of enhanced BMP14 expression in liver fibrosis, we identified activated HSCs. Furthermore, we found that BMP14 exhibits pro-fibrotic effects in HSCs. Similar to cartilage injury in joints, the upregulation of BMP14 upon liver injury may serve as a molecular mechanism to support tissue repair. However, if the hepatocellular injury persists, such as in chronic liver injury, the physiological reaction becomes disturbed, leading to progressive hepatic fibrosis and, herewith, also increasing HCC risk. Moreover, besides indirectly promoting hepatocancerogenesis, we observed pro-tumorigenic effects of BMP14 on HCC cells. Together, these findings indicate BMP14 as a potential novel therapeutic target for both hepatic fibrosis and HCC formation and progression.
RNA interference is becoming more and more exploited for the targeted depletion of genes in HSCs and for the treatment of liver fibrosis and cancer [38,39], and here BMP14 also appears as a potential target. The inhibition of BMP14 receptors could be another potential therapeutic strategy. A previous study applying biosensor analysis revealed ligand–receptor interactions for BMP14 to Alk3 (BMPR1a) and Alk6 (BMPR1b), with preferential binding to Alk6 [40]. Furthermore, Nishitoh et al. found that BMP14 binds to Alk6 in ROB-C26 cells (rat osteoprogenitor-like cell line) and COS-1 cells (fibroblast-like monkey kidney cells) [41]. Here, we found that the chemical inhibitor DMH1 almost completely abrogated the BMP14-induced upregulation of ID1 expression in HCC cells, suggesting that at least parts of the pro-tumorigenic effects of BMP14 on HCC cells are mediated via Alk3 and/or Alk2. Further studies are required to assess the receptors involved in the pro-fibrotic and pro-tumorigenic effects of BMP14 in the liver.
Generally, the role of BMPs in liver fibrosis and cancer is complex, and previous studies of other BMP family members have shown that the effects of these growth factors can be diverse.
For example, BMP7 is upregulated after liver injury and exhibits anti-fibrotic effects, which can be attributed to a decreased expression of α-SMA and/or collagen [42]. In contrast, BMP4 shows pro-fibrotic effects in experimental models of liver fibrosis [43]. BMP4 expression has also been shown to be upregulated in patients with cirrhosis and HCC, and further to correlate with cancer progression [44]. Moreover, Arndt et al. have demonstrated that mice with global BMP6 knockout revealed increased hepatic inflammation and fibrosis in two different experimental models of non-alcoholic steatohepatitis (NASH), indicating an anti-fibrogenic role of this BMP family member [45]. For BMP9, both pro- and anti-fibrogenic effects have been described. Desroches-Castan et al. found that BMP9 gene deletion triggers hepatic perisinusoidal fibrosis, indicating a protective effect of BMP9 [46]. As an underlying molecular mechanism, their findings indicated that BMP9 prevented the loss of fenestration of sinusoidal endothelial cells. On the other hand, Li et al. described how, in mouse models, recombinant BMP9 overexpression accelerated liver fibrosis, while BMP9 knockdown attenuated liver fibrogenesis [47].
In HCC, BMP2 and BMP10 were shown to exhibit anti-tumorigenic effects, thereby inhibiting HCC progression [48,49]. In HCC patients, BMP4 expression was found to be upregulated and to correlate with cancer progression [44], pointing to a protumorigenic effect of BMP4 in HCC. Furthermore, BMP9 has been shown to promote the growth of HCC cell lines [50]. Another study found that BMP9 induces angiogenesis via the ID1 signaling axis, and therefore acts as a pro-tumorigenic [51].
Together, these previous studies have indicated the complexity of the BMP system in diseased livers, since individual members of the large BMP protein family can even have opposite effects on critical pathological steps of chronic liver diseases.
The data of our study suggest that BMP14 secreted by activated HSCs acts on both HSCs themselves and HCC cells, and herewith promotes fibrosis as well as tumorigenicity of HCC cells (Figure 4). In future studies, the role of BMP14 and its molecular mechanisms of action need to be validated in vivo. Our study may be the basis for the further evaluation of the potential of BMP14 as a new therapeutic target against liver fibrosis and cancer. Furthermore, this soluble growth factor may also have potential as a diagnostic marker in patients with chronic liver disease.
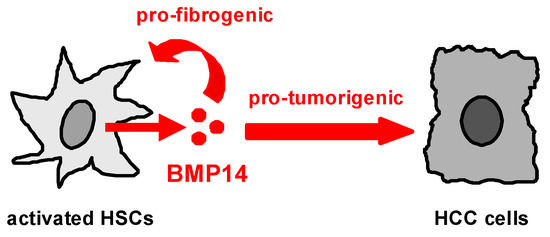
Figure 4.
Proposed mechanism of action of enhanced BMP14 expression in the (fibrotic) liver. BMP14 is expressed and secreted by activated hepatic stellate cells (HSCs). It acts pro-fibrogenically on the HSCs itself, and exerts pro-tumorigenic effects on HCC cells.
Author Contributions
J.S. performed the experiments. J.S. and C.H. analyzed the data. W.E.T. and A.B. provided material and methods. J.S., A.B. and C.H. designed the project. J.S. and C.H. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by funding from the German Cancer Aid (Deutsche Krebshilfe; 70115020) to A.B. and C.H.
Institutional Review Board Statement
The animal studies were approved by the Committee for Animal Health and Care of the local government (54-2532.1-49/13), and conformed to international guidelines on the ethical use of animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We acknowledge the Human Tissue and Cell Research (HTCR) Foundation and HTCR-Services for providing primary human cells for in vitro studies. The authors thank Petra Wenzeler and Jennifer Czekalla for excellent technical assistance.
Conflicts of Interest
All contributing authors declare no conflict of interests.
References
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Dhar, D.; Baglieri, J.; Kisseleva, T.; Brenner, D.A. Mechanisms of liver fibrosis and its role in liver cancer. Exp. Biol. Med. 2020, 245, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Barry, A.E.; Baldeosingh, R.; Lamm, R.; Patel, K.; Zhang, K.; Dominguez, D.A.; Kirton, K.J.; Shah, A.P.; Dang, H. Hepatic Stellate Cells and Hepatocarcinogenesis. Front. Cell Dev. Biol. 2020, 8, 709. [Google Scholar] [CrossRef]
- Amann, T.; Bataille, F.; Spruss, T.; Muhlbauer, M.; Gabele, E.; Scholmerich, J.; Kiefer, P.; Bosserhoff, A.K.; Hellerbrand, C. Activated hepatic stellate cells promote tumorigenicity of hepatocellular carcinoma. Cancer Sci. 2009, 100, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Puerto, M.C.; Iyengar, P.V.; Garcia de Vinuesa, A.; Ten Dijke, P.; Sanchez-Duffhues, G. Bone morphogenetic protein receptor signal transduction in human disease. J. Pathol. 2019, 247, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Guo, J.; Yao, X.; Guo, Z.; Guo, F. Growth differentiation factor 5 in cartilage and osteoarthritis: A possible therapeutic candidate. Cell Prolif. 2021, 54, e12998. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, W.; Wu, X.; Wu, J.; Sun, C.; Luo, F.; Pei, Z. Systemic Overexpression of GDF5 in Adipocytes but Not Hepatocytes Alleviates High-Fat Diet-Induced Nonalcoholic Fatty Liver in Mice. Can. J. Gastroenterol. Hepatol. 2021, 2021, 8894685. [Google Scholar] [CrossRef]
- Enescu, A.S.; Margaritescu, C.L.; Craitoiu, M.M.; Enescu, A.; Craitoiu, S. The involvement of growth differentiation factor 5 (GDF5) and aggrecan in the epithelial-mesenchymal transition of salivary gland pleomorphic adenoma. Rom. J. Morphol. Embryol. 2013, 54, 969–976. [Google Scholar]
- Margheri, F.; Schiavone, N.; Papucci, L.; Magnelli, L.; Serrati, S.; Chilla, A.; Laurenzana, A.; Bianchini, F.; Calorini, L.; Torre, E.; et al. GDF5 regulates TGFss-dependent angiogenesis in breast carcinoma MCF-7 cells: In vitro and in vivo control by anti-TGFss peptides. PLoS ONE 2012, 7, e50342. [Google Scholar] [CrossRef]
- Muhlbauer, M.; Bosserhoff, A.K.; Hartmann, A.; Thasler, W.E.; Weiss, T.S.; Herfarth, H.; Lock, G.; Scholmerich, J.; Hellerbrand, C. A novel MCP-1 gene polymorphism is associated with hepatic MCP-1 expression and severity of HCV-related liver disease. Gastroenterology 2003, 125, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Hui, A.Y.; Albanis, E.; Arthur, M.J.; O’Byrne, S.M.; Blaner, W.S.; Mukherjee, P.; Friedman, S.L.; Eng, F.J. Human hepatic stellate cell lines, LX-1 and LX-2: New tools for analysis of hepatic fibrosis. Gut 2005, 54, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Thasler, W.E.; Weiss, T.S.; Schillhorn, K.; Stoll, P.T.; Irrgang, B.; Jauch, K.W. Charitable State-Controlled Foundation Human Tissue and Cell Research: Ethic and Legal Aspects in the Supply of Surgically Removed Human Tissue for Research in the Academic and Commercial Sector in Germany. Cell Tissue Bank. 2003, 4, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, P.; Koch, A.; Fritz, V.; Hartmann, A.; Bosserhoff, A.K.; Hellerbrand, C. Wild type Kirsten rat sarcoma is a novel microRNA-622-regulated therapeutic target for hepatocellular carcinoma and contributes to sorafenib resistance. Gut 2018, 67, 1328–1341. [Google Scholar] [CrossRef]
- Hannus, M.; Beitzinger, M.; Engelmann, J.C.; Weickert, M.T.; Spang, R.; Hannus, S.; Meister, G. siPools: Highly complex but accurately defined siRNA pools eliminate off-target effects. Nucleic Acids Res. 2014, 42, 8049–8061. [Google Scholar] [CrossRef]
- Mazagova, M.; Wang, L.; Anfora, A.T.; Wissmueller, M.; Lesley, S.A.; Miyamoto, Y.; Eckmann, L.; Dhungana, S.; Pathmasiri, W.; Sumner, S.; et al. Commensal microbiota is hepatoprotective and prevents liver fibrosis in mice. FASEB J. 2015, 29, 1043–1055. [Google Scholar] [CrossRef]
- Sommer, J.; Dorn, C.; Gabele, E.; Bataille, F.; Freese, K.; Seitz, T.; Thasler, W.E.; Buttner, R.; Weiskirchen, R.; Bosserhoff, A.; et al. Four-And-A-Half LIM-Domain Protein 2 (FHL2) Deficiency Aggravates Cholestatic Liver Injury. Cells 2020, 9, 248. [Google Scholar] [CrossRef]
- Mahli, A.; Seitz, T.; Beckroge, T.; Freese, K.; Thasler, W.E.; Benkert, M.; Dietrich, P.; Weiskirchen, R.; Bosserhoff, A.; Hellerbrand, C. Bone Morphogenetic Protein-8B Expression is Induced in Steatotic Hepatocytes and Promotes Hepatic Steatosis and Inflammation In Vitro. Cells 2019, 8, 457. [Google Scholar] [CrossRef]
- Dorn, C.; Weiss, T.S.; Heilmann, J.; Hellerbrand, C. Xanthohumol, a prenylated chalcone derived from hops, inhibits proliferation, migration and interleukin-8 expression of hepatocellular carcinoma cells. Int. J. Oncol. 2010, 36, 435–441. [Google Scholar] [CrossRef]
- Freese, K.; Seitz, T.; Dietrich, P.; Lee, S.M.L.; Thasler, W.E.; Bosserhoff, A.; Hellerbrand, C. Histone Deacetylase Expressions in Hepatocellular Carcinoma and Functional Effects of Histone Deacetylase Inhibitors on Liver Cancer Cells In Vitro. Cancers 2019, 11, 1587. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Koyama, Y.; Liu, X.; Xu, J.; Ma, H.Y.; Liang, S.; Kim, I.H.; Brenner, D.A.; Kisseleva, T. Promising Therapy Candidates for Liver Fibrosis. Front. Physiol. 2016, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Korchynskyi, O.; ten Dijke, P. Identification and functional characterization of distinct critically important bone morphogenetic protein-specific response elements in the Id1 promoter. J. Biol. Chem. 2002, 277, 4883–4891. [Google Scholar] [CrossRef] [PubMed]
- Hollnagel, A.; Oehlmann, V.; Heymer, J.; Ruther, U.; Nordheim, A. Id genes are direct targets of bone morphogenetic protein induction in embryonic stem cells. J. Biol. Chem. 1999, 274, 19838–19845. [Google Scholar] [CrossRef]
- Wiercinska, E.; Wickert, L.; Denecke, B.; Said, H.M.; Hamzavi, J.; Gressner, A.M.; Thorikay, M.; ten Dijke, P.; Mertens, P.R.; Breitkopf, K.; et al. Id1 is a critical mediator in TGF-beta-induced transdifferentiation of rat hepatic stellate cells. Hepatology 2006, 43, 1032–1041. [Google Scholar] [CrossRef]
- Tajima, K.; Terai, S.; Takami, T.; Kawaguchi, K.; Okita, K.; Sakaida, I. Importance of inhibitor of DNA binding/differentiation 2 in hepatic stellate cell differentiation and proliferation. Hepatol. Res. 2007, 37, 647–655. [Google Scholar] [CrossRef]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repecka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef]
- Yin, X.; Tang, B.; Li, J.H.; Wang, Y.; Zhang, L.; Xie, X.Y.; Zhang, B.H.; Qiu, S.J.; Wu, W.Z.; Ren, Z.G. ID1 promotes hepatocellular carcinoma proliferation and confers chemoresistance to oxaliplatin by activating pentose phosphate pathway. J. Exp. Clin. Cancer Res. 2017, 36, 166. [Google Scholar] [CrossRef]
- Ao, J.; Meng, J.; Zhu, L.; Nie, H.; Yang, C.; Li, J.; Gu, J.; Lin, Q.; Long, W.; Dong, X.; et al. Activation of androgen receptor induces ID1 and promotes hepatocellular carcinoma cell migration and invasion. Mol. Oncol. 2012, 6, 507–515. [Google Scholar] [CrossRef]
- Hao, J.; Ho, J.N.; Lewis, J.A.; Karim, K.A.; Daniels, R.N.; Gentry, P.R.; Hopkins, C.R.; Lindsley, C.W.; Hong, C.C. In vivo structure-activity relationship study of dorsomorphin analogues identifies selective VEGF and BMP inhibitors. ACS Chem. Biol. 2010, 5, 245–253. [Google Scholar] [CrossRef]
- Rhee, D.K.; Marcelino, J.; Baker, M.; Gong, Y.; Smits, P.; Lefebvre, V.; Jay, G.D.; Stewart, M.; Wang, H.; Warman, M.L.; et al. The secreted glycoprotein lubricin protects cartilage surfaces and inhibits synovial cell overgrowth. J. Clin. Investig. 2005, 115, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Takahata, Y.; Hagino, H.; Kimura, A.; Urushizaki, M.; Yamamoto, S.; Wakamori, K.; Murakami, T.; Hata, K.; Nishimura, R. Regulatory Mechanisms of Prg4 and Gdf5 Expression in Articular Cartilage and Functions in Osteoarthritis. Int. J. Mol. Sci. 2022, 23, 4672. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.T.; Lin, K.; Nandedkar, M.; Camargo, M.; Cervenka, J.; Luyten, F.P. A human chondrodysplasia due to a mutation in a TGF-beta superfamily member. Nat. Genet. 1996, 12, 315–317. [Google Scholar] [CrossRef]
- Al-Yahyaee, S.A.; Al-Kindi, M.N.; Habbal, O.; Kumar, D.S. Clinical and molecular analysis of Grebe acromesomelic dysplasia in an Omani family. Am. J. Med. Genet. A 2003, 121A, 9–14. [Google Scholar] [CrossRef]
- Storm, E.E.; Kingsley, D.M. GDF5 coordinates bone and joint formation during digit development. Dev. Biol. 1999, 209, 11–27. [Google Scholar] [CrossRef]
- Coleman, C.M.; Vaughan, E.E.; Browe, D.C.; Mooney, E.; Howard, L.; Barry, F. Growth differentiation factor-5 enhances in vitro mesenchymal stromal cell chondrogenesis and hypertrophy. Stem Cells Dev. 2013, 22, 1968–1976. [Google Scholar] [CrossRef] [PubMed]
- Coleman, C.M.; Scheremeta, B.H.; Boyce, A.T.; Mauck, R.L.; Tuan, R.S. Delayed fracture healing in growth differentiation factor 5-deficient mice: A pilot study. Clin. Orthop. Relat. Res. 2011, 469, 2915–2924. [Google Scholar] [CrossRef]
- Zhao, Z.; Lin, C.Y.; Cheng, K. siRNA- and miRNA-based therapeutics for liver fibrosis. Transl. Res. 2019, 214, 17–29. [Google Scholar] [CrossRef]
- Omar, R.; Yang, J.; Liu, H.; Davies, N.M.; Gong, Y. Hepatic Stellate Cells in Liver Fibrosis and siRNA-Based Therapy. Rev. Physiol. Biochem. Pharmacol. 2016, 172, 1–37. [Google Scholar] [CrossRef]
- Nickel, J.; Kotzsch, A.; Sebald, W.; Mueller, T.D. A single residue of GDF-5 defines binding specificity to BMP receptor IB. J. Mol. Biol. 2005, 349, 933–947. [Google Scholar] [CrossRef]
- Nishitoh, H.; Ichijo, H.; Kimura, M.; Matsumoto, T.; Makishima, F.; Yamaguchi, A.; Yamashita, H.; Enomoto, S.; Miyazono, K. Identification of type I and type II serine/threonine kinase receptors for growth/differentiation factor-5. J. Biol. Chem. 1996, 271, 21345–21352. [Google Scholar] [CrossRef]
- Dituri, F.; Cossu, C.; Mancarella, S.; Giannelli, G. The Interactivity between TGFbeta and BMP Signaling in Organogenesis, Fibrosis, and Cancer. Cells 2019, 8, 1130. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Shen, H.; Sun, Y.; Li, P.; Burczynski, F.; Namaka, M.; Gong, Y. Bone morphogenetic protein 4 mediates bile duct ligation induced liver fibrosis through activation of Smad1 and ERK1/2 in rat hepatic stellate cells. J. Cell. Physiol. 2006, 207, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.W.; Hsia, Y.; Yang, W.Y.; Lin, Y.I.; Li, C.C.; Tsai, T.F.; Chang, K.W.; Shieh, G.S.; Tsai, S.F.; Wang, H.D.; et al. Identification of the common regulators for hepatocellular carcinoma induced by hepatitis B virus X antigen in a mouse model. Carcinogenesis 2012, 33, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Arndt, S.; Wacker, E.; Dorn, C.; Koch, A.; Saugspier, M.; Thasler, W.E.; Hartmann, A.; Bosserhoff, A.K.; Hellerbrand, C. Enhanced expression of BMP6 inhibits hepatic fibrosis in non-alcoholic fatty liver disease. Gut 2015, 64, 973–981. [Google Scholar] [CrossRef]
- Desroches-Castan, A.; Tillet, E.; Ricard, N.; Ouarne, M.; Mallet, C.; Belmudes, L.; Coute, Y.; Boillot, O.; Scoazec, J.Y.; Bailly, S.; et al. Bone Morphogenetic Protein 9 Is a Paracrine Factor Controlling Liver Sinusoidal Endothelial Cell Fenestration and Protecting Against Hepatic Fibrosis. Hepatology 2019, 70, 1392–1408. [Google Scholar] [CrossRef]
- Li, P.; Li, Y.; Zhu, L.; Yang, Z.; He, J.; Wang, L.; Shang, Q.; Pan, H.; Wang, H.; Ma, X.; et al. Targeting secreted cytokine BMP9 gates the attenuation of hepatic fibrosis. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 709–720. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, X.; Wang, H.; Yan, W.; Zhang, Q.; Chang, X. Bone morphogenetic protein 2 inhibits hepatocellular carcinoma growth and migration through downregulation of the PI3K/AKT pathway. Tumour Biol. 2014, 35, 5189–5198. [Google Scholar] [CrossRef]
- Yuan, Y.M.; Ma, N.; Zhang, E.B.; Chen, T.W.; Jiang, H.; Yin, F.F.; Wang, J.J.; Zhang, F.K.; Ni, Q.Z.; Wang, X.; et al. BMP10 suppresses hepatocellular carcinoma progression via PTPRS-STAT3 axis. Oncogene 2019, 38, 7281–7293. [Google Scholar] [CrossRef]
- Herrera, B.; Garcia-Alvaro, M.; Cruz, S.; Walsh, P.; Fernandez, M.; Roncero, C.; Fabregat, I.; Sanchez, A.; Inman, G.J. BMP9 is a proliferative and survival factor for human hepatocellular carcinoma cells. PLoS ONE 2013, 8, e69535. [Google Scholar] [CrossRef]
- Chen, H.; Nio, K.; Tang, H.; Yamashita, T.; Okada, H.; Li, Y.; Doan, P.T.B.; Li, R.; Lv, J.; Sakai, Y.; et al. BMP9-ID1 Signaling Activates HIF-1alpha and VEGFA Expression to Promote Tumor Angiogenesis in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2022, 23, 1475. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).