
Murine Models of Obesity

 , , ,
, , ,  and
and
Abstract
1. Introduction
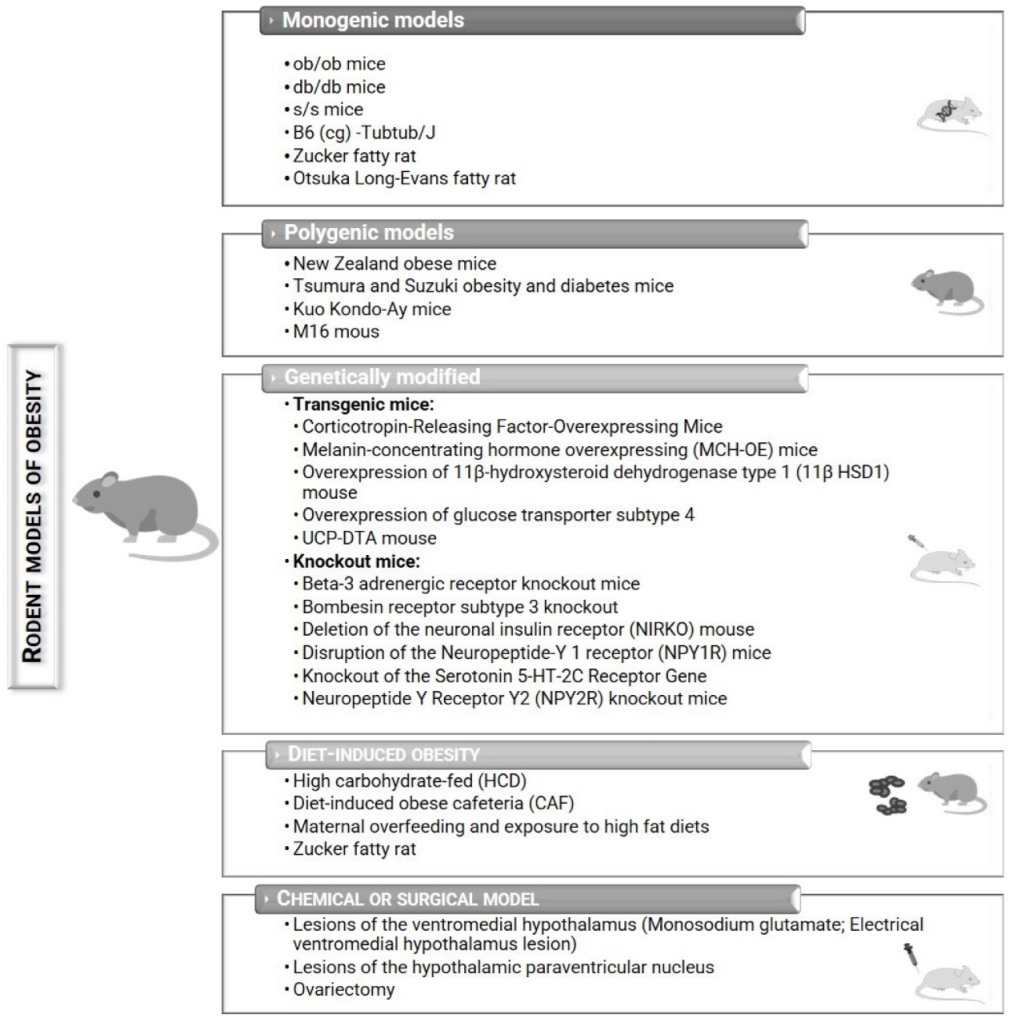
2. Rodent Models of Obesity
2.1. Monogenic Models
2.1.1. ob/ob Mice
2.1.2. db/db Mice
2.1.3. s/s Mice
2.1.4. B6 (cg)-Tubtub/J
2.1.5. Zucker Fatty Rat
2.1.6. Otsuka Long-Evans Tokushima Fatty Rat
2.2. Polygenic Models
2.2.1. New Zealand Obese Mice
2.2.2. Tsumura and Suzuki Obesity and Diabetes Mice
2.2.3. Kuo Kondo-Ay Mice
2.2.4. M16 Mice
2.3. Genetically Modified Mice
2.3.1. Transgenic Mice
Corticotropin-Releasing Factor Overexpressing Mice
Melanin-Concentrating Hormone Overexpressing (MCH-OE) Mice
Overexpression of 11β-Hydroxysteroid Dehydrogenase Type 1 (11β HSD1) Mice
Overexpression of Glucose Transporter Subtype 4
UCP-DTA Mice
2.3.2. Knockout Mice
Beta-3 Adrenergic Receptor Knockout Mice
Bombesin Receptor Subtype 3 Knockout Mice
Deletion of the Neuronal Insulin Receptor (NIRKO) Mice
Disruption of the Neuropeptide-Y 1 Receptor (NPY1R) Mice
Knockout of the Serotonin 5-HT-2C Receptor Gene
Neuropeptide Y Receptor Y2 (NPY2R) Knockout Mice
2.4. Chemical or Surgical Models of Obesity
2.4.1. Lesions of the Ventromedial Hypothalamus
Monosodium Glutamate (MSG)
Electrical VMH Lesion
2.4.2. Lesions of the Hypothalamic Paraventricular Nucleus
2.4.3. Lesions of the Arcuate Nucleus
2.4.4. Ovariectomy
2.5. Diet-Induced Obesity (DIO)
2.5.1. High-Fat Diet (HFD)/Exposure to High-Fat and Palatable Diets
2.5.2. High-Carbohydrate Diet (HCD)
2.5.3. Cafeteria Diet (CAF)
2.5.4. Maternal Overfeeding and Exposure to High-Fat Diets
2.5.5. Other Models
2.5.6. Animal Models and Clinical Setting
3. Potential Markers for Obesity Assessment in Rodents
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 10 January 2022).
- Lau, D.C.; Douketis, J.D.; Morrison, K.M.; Hramiak, I.M.; Sharma, A.M.; Ur, E.; Obesity Canada Clinical Practice Guidelines Expert Panel. 2006 Canadian clinical practice guidelines on the management and prevention of obesity in adults and children [summary]. CMAJ 2007, 176, S1–S13. [Google Scholar] [CrossRef] [PubMed]
- Novelli, E.L.; Diniz, Y.S.; Galhardi, C.M.; Ebaid, G.M.; Rodrigues, H.G.; Mani, F.; Fernandes, A.A.; Cicogna, A.C.; Novelli Filho, J.L. Anthropometrical parameters and markers of obesity in rats. Lab. Anim. 2007, 41, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Kaila, B.; Raman, M. Obesity: A review of pathogenesis and management strategies. Can. J. Gastroenterol. 2008, 22, 61–68. [Google Scholar] [CrossRef]
- Mittwede, P.N.; Clemmer, J.S.; Bergin, P.F.; Xiang, L. Obesity and Critical Illness: Insights from Animal Models. Shock 2016, 45, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Kanasaki, K.; Koya, D. Biology of obesity: Lessons from animal models of obesity. J. Biomed. Biotechnol. 2011, 2011, 197636. [Google Scholar] [CrossRef] [PubMed]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; de Angelis, M.H.; Schurmann, A.; et al. Animal models of obesity and diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef]
- Grotto, D.; Camargo, I.F.; Kodaira, K.; Mazzei, L.G.; Castro, J.; Vieira, R.A.L.; Bergamaschi, C.C.; Lopes, L.C. Effect of mushrooms on obesity in animal models: Study protocol for a systematic review and meta-analysis. Syst. Rev. 2019, 8, 288. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R.M.; Elkhayat, E.S.; El Dine, R.S. Natural anti-obesity agents. Bull. Fac. Pharm. Cairo Univ. 2014, 52, 269–284. [Google Scholar] [CrossRef]
- Parasuraman, S.; Wen, L.E. Animal Model for Obesity-An Overview. Syst. Rev. Pharm. 2016, 6, 9–12. [Google Scholar] [CrossRef]
- Geiger, B.M.; Pothos, E.N. Chapter 1—Translating Animal Models of Obesity and Diabetes to the Clinic. In Handbook of Behavioral Neuroscience; Nomikos, G.G., Feltner, D.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 29, pp. 1–16. [Google Scholar]
- Barré-Sinoussi, F.; Montagutelli, X. Animal models are essential to biological research: Issues and perspectives. Future Sci. OA 2015, 1, FSO63. [Google Scholar] [CrossRef]
- Barrett, P.; Mercer, J.G.; Morgan, P.J. Preclinical models for obesity research. Dis. Models Mech. 2016, 9, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Lutz, T.A.; Woods, S.C. Overview of animal models of obesity. Curr. Protoc. Pharm. 2012, 58, 5–61. [Google Scholar] [CrossRef] [PubMed]
- Preguica, I.; Alves, A.; Nunes, S.; Fernandes, R.; Gomes, P.; Viana, S.D.; Reis, F. Diet-induced rodent models of obesity-related metabolic disorders-A guide to a translational perspective. Obes. Rev. 2020, 21, e13081. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.; Hambly, C.; Mitchell, S.; Krol, E. Animal models of obesity. Obes. Rev. 2007, 8 (Suppl. 1), 55–61. [Google Scholar] [CrossRef]
- Suleiman, J.B.; Mohamed, M.; Bakar, A.B.A. A systematic review on different models of inducing obesity in animals: Advantages and limitations. J. Adv. Vet. Anim. Res. 2020, 7, 103–114. [Google Scholar] [CrossRef]
- Speakman, J.; Hambly, C.; Mitchell, S.; Krol, E. The contribution of animal models to the study of obesity. Lab. Anim. 2008, 42, 413–432. [Google Scholar] [CrossRef]
- Crujeiras, A.B.; Carreira, M.C.; Cabia, B.; Andrade, S.; Amil, M.; Casanueva, F.F. Leptin resistance in obesity: An epigenetic landscape. Life Sci. 2015, 140, 57–63. [Google Scholar] [CrossRef]
- Liu, J.; Lee, J.; Salazar Hernandez, M.A.; Mazitschek, R.; Ozcan, U. Treatment of obesity with celastrol. Cell 2015, 161, 999–1011. [Google Scholar] [CrossRef]
- Smith, J.T.; Acohido, B.V.; Clifton, D.K.; Steiner, R.A. KiSS-1 neurones are direct targets for leptin in the ob/ob mouse. J. Neuroendocr. 2006, 18, 298–303. [Google Scholar] [CrossRef]
- Ferguson, D.; Blenden, M.; Hutson, I.; Du, Y.; Harris, C.A. Mouse Embryonic Fibroblasts Protect ob/ob Mice From Obesity and Metabolic Complications. Endocrinology 2018, 159, 3275–3286. [Google Scholar] [CrossRef]
- van der Spek, R.; Kreier, F.; Fliers, E.; Kalsbeek, A. Chapter 11—Circadian rhythms in white adipose tissue. In Progress in Brain Research; Kalsbeek, A., Merrow, M., Roenneberg, T., Foster, R.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 199, pp. 183–201. [Google Scholar]
- Bao, R.; Meng, Y.; Zhang, H.; Yang, C.; Li, W.; Zhang, C.; Zhang, J.; Sun, R.; Li, Z.; Jiang, W.; et al. Elaiophylin reduces body weight and lowers glucose levels in obese mice by activating AMPK. Cell Death Dis. 2021, 12, 972. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.; Zhang, Y.; Zhao, F.; Chen, K.; Wu, T.; Liu, Q.; Huang, S.; Zhang, A.; Jia, Z. Celastrol inhibits intestinal lipid absorption by reprofiling the gut microbiota to attenuate high-fat diet-induced obesity. iScience 2021, 24, 102077. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, L.; Li, X.; Wu, Y.; Yin, F.; Liu, J. Trilobatin ameliorates insulin resistance through IRS-AKT-GLUT4 signaling pathway in C2C12 myotubes and ob/ob mice. Chin. Med. 2020, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Simperova, A.; Al-Nakkash, L.; Faust, J.J.; Sweazea, K.L. Genistein supplementation prevents weight gain but promotes oxidative stress and inflammation in the vasculature of female obese ob/ob mice. Nutr. Res. 2016, 36, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Wargent, E.T.; Zaibi, M.S.; Silvestri, C.; Hislop, D.C.; Stocker, C.J.; Stott, C.G.; Guy, G.W.; Duncan, M.; Di Marzo, V.; Cawthorne, M.A. The cannabinoid Delta(9)-tetrahydrocannabivarin (THCV) ameliorates insulin sensitivity in two mouse models of obesity. Nutr. Diabetes 2013, 3, e68. [Google Scholar] [CrossRef]
- Sun, S.-S.; Wang, K.; Ma, K.; Bao, L.; Liu, H.-W. An insoluble polysaccharide from the sclerotium of Poria cocos improves hyperglycemia, hyperlipidemia and hepatic steatosis in ob/ob mice via modulation of gut microbiota. Chin. J. Nat. Med. 2019, 17, 3–14. [Google Scholar] [CrossRef]
- Chung, M.Y.; Park, H.J.; Manautou, J.E.; Koo, S.I.; Bruno, R.S. Green tea extract protects against nonalcoholic steatohepatitis in ob/ob mice by decreasing oxidative and nitrative stress responses induced by proinflammatory enzymes. J. Nutr. Biochem. 2012, 23, 361–367. [Google Scholar] [CrossRef]
- Zheng, J.; Manabe, Y.; Sugawara, T. Siphonaxanthin, a carotenoid from green algae Codium cylindricum, protects Ob/Ob mice fed on a high-fat diet against lipotoxicity by ameliorating somatic stresses and restoring anti-oxidative capacity. Nutr. Res. 2020, 77, 29–42. [Google Scholar] [CrossRef]
- Kitada, M.; Ogura, Y.; Koya, D. Rodent models of diabetic nephropathy: Their utility and limitations. Int. J. Nephrol. Renov. Dis. 2016, 9, 279–290. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Mills, D.A.; Murphy, K.; Noratto, G. Effect of barley supplementation on the fecal microbiota, caecal biochemistry, and key biomarkers of obesity and inflammation in obese db/db mice. Eur. J. Nutr. 2018, 57, 2513–2528. [Google Scholar] [CrossRef]
- Guo, Y.; Fu, W.; Xin, Y.; Bai, J.; Peng, H.; Fu, L.; Liu, J.; Li, L.; Ma, Y.; Jiang, H. Antidiabetic and Antiobesity Effects of Artemether in db/db Mice. BioMed Res. Int. 2018, 2018, 8639523. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Koh, E.; Kim, D.; Lee, N.; Cho, S.M.; Lee, Y.J.; Cho, I.H.; Yang, H.J. Dendropanax trifidus Sap-Mediated Suppression of Obese Mouse Body Weight and the Metabolic Changes Related with Estrogen Receptor Alpha and AMPK-ACC Pathways in Muscle Cells. Nutrients 2022, 14, 1098. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Fang, K.; Dan, X.; Gu, M. Crocin ameliorates hepatic steatosis through activation of AMPK signaling in db/db mice. Lipids Health Dis. 2019, 18, 11. [Google Scholar] [CrossRef]
- Bates, S.H.; Stearns, W.H.; Dundon, T.A.; Schubert, M.; Tso, A.W.; Wang, Y.; Banks, A.S.; Lavery, H.J.; Haq, A.K.; Maratos-Flier, E.; et al. STAT3 signalling is required for leptin regulation of energy balance but not reproduction. Nature 2003, 421, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Saadat, N.; IglayReger, H.B.; Myers, M.G., Jr.; Bodary, P.; Gupta, S.V. Differences in metabolomic profiles of male db/db and s/s, leptin receptor mutant mice. Physiol. Genom. 2012, 44, 374–381. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Coleman, D.L.; Eicher, E.M. Fat (fat) and Tubby (tub): Two Autosomal Recessive Mutations Causing Obesity Syndromes in the Mouse. J. Hered. 1990, 81, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Kleyn, P.W.; Fan, W.; Kovats, S.G.; Lee, J.J.; Pulido, J.C.; Wu, Y.; Berkemeier, L.R.; Misumi, D.J.; Holmgren, L.; Charlat, O.; et al. Identification and Characterization of the Mouse Obesity Gene tubby: A Member of a Novel Gene Family. Cell 1996, 85, 281–290. [Google Scholar] [CrossRef]
- Hernández-Granados, M.J.; Ramírez-Emiliano, J.; Elena Franco-Robles, E. Rodent Models of Obesity and Diabetes. In Experimental Animal Models of Human Diseases—An Effective Therapeutic Strategy; Bartholomew, I., Ed.; IntechOpen: London, UK, 2018. [Google Scholar]
- Yorek, M.A. Alternatives to the Streptozotocin-Diabetic Rodent. Int. Rev. Neurobiol. 2016, 127, 89–112. [Google Scholar] [CrossRef]
- Zucker, L.M.; Zucker, T.F. Fatty, A new mutation in the rat. J. Hered. 1961, 52, 275–278. [Google Scholar] [CrossRef]
- Owens, D.R. Chapter 23—Spontaneous, Surgically and Chemically Induced Models of Disease. In The Laboratory Rat, 2nd ed.; Suckow, M.A., Weisbroth, S.H., Franklin, C.L., Eds.; Academic Press: Burlington, VT, USA, 2006; pp. 711–732. [Google Scholar]
- Agil, A.; Navarro-Alarcon, M.; Ali, F.A.Z.; Albrakati, A.; Salagre, D.; Campoy, C.; Elmahallawy, E.K. Melatonin Enhances the Mitochondrial Functionality of Brown Adipose Tissue in Obese-Diabetic Rats. Antioxidants 2021, 10, 1482. [Google Scholar] [CrossRef]
- Kato, Y.; Sakoh, M.; Nagai, T.; Yoshida, A.; Ishida, H.; Inoue, N.; Yanagita, T.; Nagao, K. Ozonated Olive Oil Alleviates Hepatic Steatosis in Obese Zucker (fa/fa) Rats. J. Oleo Sci. 2022; ahead of print. [Google Scholar] [CrossRef]
- Szabadfi, K.; Pinter, E.; Reglodi, D.; Gabriel, R. Chapter One—Neuropeptides, Trophic Factors, and Other Substances Providing Morphofunctional and Metabolic Protection in Experimental Models of Diabetic Retinopathy. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 311, pp. 1–121. [Google Scholar]
- Cho, I.J.; Oh, D.H.; Yoo, J.; Hwang, Y.C.; Ahn, K.J.; Chung, H.Y.; Jeong, S.W.; Moon, J.Y.; Lee, S.H.; Lim, S.J.; et al. Allopurinol ameliorates high fructose diet induced hepatic steatosis in diabetic rats through modulation of lipid metabolism, inflammation, and ER stress pathway. Sci. Rep. 2021, 11, 9894. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, M. Chapter 8—Models to Investigate Cardiac Metabolism. In The Scientist’s Guide to Cardiac Metabolism; Schwarzer, M., Doenst, T., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 103–122. [Google Scholar]
- Shin, Y.; Lee, D.; Ahn, J.; Lee, M.; Shin, S.S.; Yoon, M. The herbal extract ALS-L1023 from Melissa officinalis reduces weight gain, elevated glucose levels and β-cell loss in Otsuka Long-Evans Tokushima fatty rats. J. Ethnopharmacol. 2021, 264, 113360. [Google Scholar] [CrossRef] [PubMed]
- Wanezaki, S.; Saito, S.; Inoue, N.; Tachibana, N.; Shirouchi, B.; Sato, M.; Yanagita, T.; Nagao, K. Soy β-Conglycinin Peptide Attenuates Obesity and Lipid Abnormalities in Obese Model OLETF Rats. J. Oleo Sci. 2020, 69, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Moran, T.H. Actions of CCK in the controls of food intake and body weight: Lessons from the CCK-A receptor deficient OLETF rat. Neuropeptides 2002, 36, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.-Y.; Lin, C.-H.; Huang, T.-H.; Chuang, S.-Y. In Vivo Rodent Models of Type 2 Diabetes and Their Usefulness for Evaluating Flavonoid Bioactivity. Nutrients 2019, 11, 530. [Google Scholar] [CrossRef]
- Wang, Y.W.; Sun, G.D.; Sun, J.; Liu, S.J.; Wang, J.; Xu, X.H.; Miao, L.N. Spontaneous type 2 diabetic rodent models. J. Diabetes Res. 2013, 2013, 401723. [Google Scholar] [CrossRef]
- Joost, H.G. The genetic basis of obesity and type 2 diabetes: Lessons from the new zealand obese mouse, a polygenic model of the metabolic syndrome. Results Probl. Cell Differ. 2010, 52, 1–11. [Google Scholar] [CrossRef]
- Kluge, R.; Scherneck, S.; Schürmann, A.; Joost, H.G. Pathophysiology and genetics of obesity and diabetes in the New Zealand obese mouse: A model of the human metabolic syndrome. Methods Mol. Biol. 2012, 933, 59–73. [Google Scholar] [CrossRef]
- Jurgens, H.S.; Schurmann, A.; Kluge, R.; Ortmann, S.; Klaus, S.; Joost, H.G.; Tschop, M.H. Hyperphagia, lower body temperature, and reduced running wheel activity precede development of morbid obesity in New Zealand obese mice. Physiol. Genom. 2006, 25, 234–241. [Google Scholar] [CrossRef]
- Korovila, I.; Höhn, A.; Jung, T.; Grune, T.; Ott, C. Reduced Liver Autophagy in High-Fat Diet Induced Liver Steatosis in New Zealand Obese Mice. Antioxidants 2021, 10, 501. [Google Scholar] [CrossRef]
- Fajardo, R.J.; Karim, L.; Calley, V.I.; Bouxsein, M.L. A review of rodent models of type 2 diabetic skeletal fragility. J. Bone Min. Res. 2014, 29, 1025–1040. [Google Scholar] [CrossRef] [PubMed]
- Noge, I.; Kagawa, Y.; Maeda, T. A new diabetic mouse model derived from the ddY strain. Biol. Pharm. Bull. 2010, 33, 988–992. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rodrigues, R. A Comprehensive Review: The Use of Animal Models in Diabetes Research. J. Anal. Pharm. Res. 2016, 3, 00071. [Google Scholar] [CrossRef]
- Hussein, G.M.; Matsuda, H.; Nakamura, S.; Akiyama, T.; Tamura, K.; Yoshikawa, M. Protective and ameliorative effects of mate (Ilex paraguariensis) on metabolic syndrome in TSOD mice. Phytomed. Int. J. Phytother. Phytopharm. 2011, 19, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Nishitsuji, K.; Watanabe, S.; Xiao, J.; Nagatomo, R.; Ogawa, H.; Tsunematsu, T.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; et al. Effect of coffee or coffee components on gut microbiome and short-chain fatty acids in a mouse model of metabolic syndrome. Sci. Rep. 2018, 8, 16173. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Nishida, T.; Baba, H.; Hatta, H.; Imura, J.; Sutoh, M.; Toyohara, S.; Hokao, R.; Watanabe, S.; Ogawa, H.; et al. Histopathological characteristics of glutamine synthetase-positive hepatic tumor lesions in a mouse model of spontaneous metabolic syndrome (TSOD mouse). Mol. Clin. Oncol. 2016, 5, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Akase, T.; Shimada, T.; Harasawa, Y.; Akase, T.; Ikeya, Y.; Nagai, E.; Iizuka, S.; Nakagami, G.; Iizaka, S.; Sanada, H.; et al. Preventive Effects of Salacia reticulata on Obesity and Metabolic Disorders in TSOD Mice. Evid.-Based Complement. Altern. Med. 2011, 2011, 484590. [Google Scholar] [CrossRef]
- Kudo, T.; Toda, T.; Ushiki, T.; Ohi, K.; Ikarashi, N.; Ochiai, W.; Sugiyama, K. Differences in the pharmacokinetics of Cyp3a substrates in TSOD and streptozotocin-induced diabetic mice. Xenobiotica 2010, 40, 282–290. [Google Scholar] [CrossRef]
- Miki, S.; Inokuma, K.I.; Takashima, M.; Nishida, M.; Sasaki, Y.; Ushijima, M.; Suzuki, J.I.; Morihara, N. Aged garlic extract suppresses the increase of plasma glycated albumin level and enhances the AMP-activated protein kinase in adipose tissue in TSOD mice. Mol. Nutr. Food Res. 2017, 61, 1600797. [Google Scholar] [CrossRef]
- Othman, M.B.; Sakamoto, K. Effect of inactivated Bifidobacterium longum intake on obese diabetes model mice (TSOD). Food Res. Int. 2020, 129, 108792. [Google Scholar] [CrossRef]
- Shida, K.; Ohsawa, M.; Takahashi, S.; Ota, H.; Tamura, T.; Kusama, N.; Nakasone, M.; Yamazaki, H.; Sobue, K. Peripheral neuropathy in the pre-diabetic state of the type 2 diabetes mouse model (TSOD mice) involves TRPV1 expression in dorsal root ganglions. IBRO Neurosci. Rep. 2022, 12, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Tanaka, T.; Tanaka, M. Potential Cancer Chemopreventive Activity of Protocatechuic Acid. J. Exp. Clin. Med. 2011, 3, 27–33. [Google Scholar] [CrossRef]
- Nishitsuji, K.; Xiao, J.; Nagatomo, R.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; Tsuneyama, K. Analysis of the gut microbiome and plasma short-chain fatty acid profiles in a spontaneous mouse model of metabolic syndrome. Sci. Rep. 2017, 7, 15876. [Google Scholar] [CrossRef] [PubMed]
- Scroyen, I.; Hemmeryckx, B.; Lijnen, H.R. From mice to men--mouse models in obesity research: What can we learn? Thromb. Haemost. 2013, 110, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-S.; Kim, J. Application of Animal Models in Diabetic Cardiomyopathy. DMJ 2021, 45, 129–145. [Google Scholar] [CrossRef]
- Allan, M.F.; Eisen, E.J.; Pomp, D. The M16 mouse: An outbred animal model of early onset polygenic obesity and diabesity. Obes. Res. 2004, 12, 1397–1407. [Google Scholar] [CrossRef]
- Huijbers, I.J. Generating Genetically Modified Mice: A Decision Guide. In Site-Specific Recombinases, 2017/08/18 ed.; Eroshenko, N., Ed.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2017; Volume 1642, pp. 1–19. [Google Scholar]
- Stengel, A.; Goebel, M.; Million, M.; Stenzel-Poore, M.P.; Kobelt, P.; Monnikes, H.; Tache, Y.; Wang, L. Corticotropin-releasing factor-overexpressing mice exhibit reduced neuronal activation in the arcuate nucleus and food intake in response to fasting. Endocrinology 2009, 150, 153–160. [Google Scholar] [CrossRef]
- Wang, L.; Goebel-Stengel, M.; Yuan, P.Q.; Stengel, A.; Tache, Y. Corticotropin-releasing factor overexpression in mice abrogates sex differences in body weight, visceral fat, and food intake response to a fast and alters levels of feeding regulatory hormones. Biol. Sex Differ. 2017, 8, 2. [Google Scholar] [CrossRef]
- Kang, W.; Tong, T.; Park, T. Corticotropin releasing factor-overexpressing mouse is a model of chronic stress-induced muscle atrophy. PLoS ONE 2020, 15, e0229048. [Google Scholar] [CrossRef]
- Ross, J.A.; Alexis, R.; Reyes, B.A.S.; Risbrough, V.; Van Bockstaele, E.J. Localization of amyloid beta peptides to locus coeruleus and medial prefrontal cortex in corticotropin releasing factor overexpressing male and female mice. Brain Struct. Funct. 2019, 224, 2385–2405. [Google Scholar] [CrossRef]
- Ludwig, D.S.; Tritos, N.A.; Mastaitis, J.W.; Kulkarni, R.; Kokkotou, E.; Elmquist, J.; Lowell, B.; Flier, J.S.; Maratos-Flier, E. Melanin-concentrating hormone overexpression in transgenic mice leads to obesity and insulin resistance. J. Clin. Investig. 2001, 107, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Pissios, P.; Ozcan, U.; Kokkotou, E.; Okada, T.; Liew, C.W.; Liu, S.; Peters, J.N.; Dahlgren, G.; Karamchandani, J.; Kudva, Y.C.; et al. Melanin concentrating hormone is a novel regulator of islet function and growth. Diabetes 2007, 56, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Pereira-da-Silva, M.; De Souza, C.T.; Gasparetti, A.L.; Saad, M.J.; Velloso, L.A. Melanin-concentrating hormone induces insulin resistance through a mechanism independent of body weight gain. J. Endocrinol. 2005, 186, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Tschop, M.; Heiman, M.L. Overview of rodent models for obesity research. Curr. Protoc. Neurosci. 2002, 17, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Lord, M.N.; Subramanian, K.; Kanoski, S.E.; Noble, E.E. Melanin-concentrating hormone and food intake control: Sites of action, peptide interactions, and appetition. Peptides 2021, 137, 170476. [Google Scholar] [CrossRef]
- Li, C.; Xia, J.; Zhu, W.; Xin, L.; An, C.; Yang, S.; Li, K. Systemic overexpression of the 11betaHSD1 promotes endoplasmic reticulum stress in multiple tissues and the development of metabolic syndrome in mice. Mol. Med. Rep. 2017, 16, 7738–7744. [Google Scholar] [CrossRef][Green Version]
- Arch, J.R. Lessons in obesity from transgenic animals. J. Endocrinol. Investig. 2002, 25, 867–875. [Google Scholar] [CrossRef]
- Dammann, C.; Stapelfeld, C.; Maser, E. Expression and activity of the cortisol-activating enzyme 11β-hydroxysteroid dehydrogenase type 1 is tissue and species-specific. Chem.-Biol. Interact. 2019, 303, 57–61. [Google Scholar] [CrossRef]
- Atkinson, M.D.; Atkinson, E. Sambucus nigra L. J. Ecol. 2002, 90, 895–923. [Google Scholar] [CrossRef]
- Tsao, T.-S.; Katz, E.B.; Pommer, D.; Charron, M.J. Amelioration of insulin resistance but not hyperinsulinemia in obese mice overexpressing GLUT4 selectively in skeletal muscle. Metab. Clin. Exp. 2000, 49, 340–346. [Google Scholar] [CrossRef]
- Shepherd, P.R.; Gnudi, L.; Tozzo, E.; Yang, H.; Leach, F.; Kahn, B.B. Adipose cell hyperplasia and enhanced glucose disposal in transgenic mice overexpressing GLUT4 selectively in adipose tissue. J. Biol. Chem. 1993, 268, 22243–22246. [Google Scholar] [CrossRef]
- Townsend, K.; Tseng, Y.H. Brown adipose tissue: Recent insights into development, metabolic function and therapeutic potential. Adipocyte 2012, 1, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Lowell, B.B.; Hamann, A.; Lawitts, J.A.; Himms-Hagen, J.; Boyer, B.B.; Kozak, L.P.; Flier, J.S. Development of obesity in transgenic mice after genetic ablation of brown adipose tissue. Nature 1993, 366, 740–742. [Google Scholar] [CrossRef]
- Klaus, S.; Munzberg, H.; Truloff, C.; Heldmaier, G. Physiology of transgenic mice with brown fat ablation: Obesity is due to lowered body temperature. Am. J. Physiol. 1998, 274, R287–R293. [Google Scholar] [CrossRef] [PubMed]
- Hamann, A.; Flier, J.S.; Lowell, B.B. Decreased brown fat markedly enhances susceptibility to diet-induced obesity, diabetes, and hyperlipidemia. Endocrinology 1996, 137, 21–29. [Google Scholar] [CrossRef]
- Callahan, Z.J.; Oxendine, M.J.; Schaeffer, P.J. Intramuscular triglyceride content precedes impaired glucose metabolism without evidence for mitochondrial dysfunction during early development of a diabetic phenotype. Appl. Physiol. Nutr. Metab. 2017, 42, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.Y. Utility and importance of gene knockout animals for nutritional and metabolic research. J. Nutr. 1998, 128, 2052–2057. [Google Scholar] [CrossRef][Green Version]
- Sakamoto, Y.; Oniki, K.; Kumagae, N.; Morita, K.; Otake, K.; Ogata, Y.; Saruwatari, J. Beta-3-adrenergic Receptor rs4994 Polymorphism Is a Potential Biomarker for the Development of Nonalcoholic Fatty Liver Disease in Overweight/Obese Individuals. Dis. Markers 2019, 2019, 4065327. [Google Scholar] [CrossRef]
- Schena, G.; Caplan, M.J. Everything You Always Wanted to Know about β3-AR * (* But Were Afraid to Ask). Cells 2019, 8, 357. [Google Scholar] [CrossRef]
- Xiao, C.; Liu, N.; Province, H.; Piñol, R.A.; Gavrilova, O.; Reitman, M.L. BRS3 in both MC4R- and SIM1-expressing neurons regulates energy homeostasis in mice. Mol. Metab. 2020, 36, 100969. [Google Scholar] [CrossRef]
- Fruhbeck, G.; Gomez-Ambrosi, J. Control of body weight: A physiologic and transgenic perspective. Diabetologia 2003, 46, 143–172. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, N.; Moreno, P.; Jensen, R.T. Bombesin receptor subtype 3 as a potential target for obesity and diabetes. Expert Opin. Targets 2015, 19, 1153–1170. [Google Scholar] [CrossRef] [PubMed]
- Kleinridders, A.; Cai, W.; Cappellucci, L.; Ghazarian, A.; Collins, W.R.; Vienberg, S.G.; Pothos, E.N.; Kahn, C.R. Insulin resistance in brain alters dopamine turnover and causes behavioral disorders. Proc. Natl. Acad. Sci. USA 2015, 112, 3463–3468. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Gautam, D.; Surjo, D.; Ueki, K.; Baudler, S.; Schubert, D.; Kondo, T.; Alber, J.; Galldiks, N.; Kustermann, E.; et al. Role for neuronal insulin resistance in neurodegenerative diseases. Proc. Natl. Acad. Sci. USA 2004, 101, 3100–3105. [Google Scholar] [CrossRef]
- Bertocchi, I.; Oberto, A.; Longo, A.; Palanza, P.; Eva, C. Conditional inactivation of Npy1r gene in mice induces sex-related differences of metabolic and behavioral functions. Horm. Behav. 2020, 125, 104824. [Google Scholar] [CrossRef] [PubMed]
- Paterlini, S.; Panelli, R.; Gioiosa, L.; Parmigiani, S.; Franceschini, P.; Bertocchi, I.; Oberto, A.; Bartolomucci, A.; Eva, C.; Palanza, P. Conditional Inactivation of Limbic Neuropeptide Y-1 Receptors Increases Vulnerability to Diet-Induced Obesity in Male Mice. Int. J. Mol. Sci. 2021, 22, 8745. [Google Scholar] [CrossRef]
- Burke, L.K.; Doslikova, B.; D’Agostino, G.; Garfield, A.S.; Farooq, G.; Burdakov, D.; Low, M.J.; Rubinstein, M.; Evans, M.L.; Billups, B.; et al. 5-HT obesity medication efficacy via POMC activation is maintained during aging. Endocrinology 2014, 155, 3732–3738. [Google Scholar] [CrossRef]
- D’Agostino, G.; Lyons, D.; Cristiano, C.; Lettieri, M.; Olarte-Sanchez, C.; Burke, L.K.; Greenwald-Yarnell, M.; Cansell, C.; Doslikova, B.; Georgescu, T.; et al. Nucleus of the Solitary Tract Serotonin 5-HT2C Receptors Modulate Food Intake. Cell Metab. 2018, 28, 619–630.e5. [Google Scholar] [CrossRef]
- Tecott, L.H.; Sun, L.M.; Akana, S.F.; Strack, A.M.; Lowenstein, D.H.; Dallman, M.F.; Julius, D. Eating disorder and epilepsy in mice lacking 5-HT2c serotonin receptors. Nature 1995, 374, 542–546. [Google Scholar] [CrossRef]
- Sargent, B.J.; Henderson, A.J. Targeting 5-HT receptors for the treatment of obesity. Curr. Opin. Pharm. 2011, 11, 52–58. [Google Scholar] [CrossRef]
- Canal, C.E.; Olaghere da Silva, U.B.; Gresch, P.J.; Watt, E.E.; Sanders-Bush, E.; Airey, D.C. The serotonin 2C receptor potently modulates the head-twitch response in mice induced by a phenethylamine hallucinogen. Psychopharmacology 2010, 209, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Nebuka, M.; Ohmura, Y.; Izawa, S.; Bouchekioua, Y.; Nishitani, N.; Yoshida, T.; Yoshioka, M. Behavioral characteristics of 5-HT2C receptor knockout mice: Locomotor activity, anxiety-, and fear memory-related behaviors. Behav. Brain Res. 2020, 379, 112394. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Lee, N.J.; Ip, C.K.; Enriquez, R.; Tasan, R.; Zhang, L.; Herzog, H. NPY derived from AGRP neurons controls feeding via Y1 and energy expenditure and food foraging behaviour via Y2 signalling. Mol. Metab. 2022, 59, 101455. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.A.; Martin, N.M.; Bloom, S.R. Hypothalamic regulation of food intake and clinical therapeutic applications. Arq. Bras. Endocrinol. Metabol. 2009, 53, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Roth, C.L.; D’Ambrosio, G.; Elfers, C. Activation of nuclear factor kappa B pathway and reduction of hypothalamic oxytocin following hypothalamic lesions. J. Syst. Integr. Neurosci. 2016, 2, 79–84. [Google Scholar] [CrossRef][Green Version]
- Gaur, A.; Pal, G.K.; Pal, P. Role of Ventromedial Hypothalamus in Sucrose-Induced Obesity on Metabolic Parameters. Ann. Neurosci. 2021, 28, 39–46. [Google Scholar] [CrossRef]
- Kiba, T.; Tanaka, K.; Numata, K.; Hoshino, M.; Misugi, K.; Inoue, S. Ventromedial hypothalamic lesion-induced vagal hyperactivity stimulates rat pancreatic cell proliferation. Gastroenterology 1996, 110, 885–893. [Google Scholar] [CrossRef]
- Kinyua, A.W.; Yang, D.J.; Chang, I.; Kim, K.W. Steroidogenic Factor 1 in the Ventromedial Nucleus of the Hypothalamus Regulates Age-Dependent Obesity. PLoS ONE 2016, 11, e0162352. [Google Scholar] [CrossRef]
- Von Diemen, V.; Trindade, E.N.; Trindade, M.R. Experimental model to induce obesity in rats. Acta Cir. Bras. 2006, 21, 425–429. [Google Scholar] [CrossRef]
- Miśkowiak, B.; Partyka, M. Effects of neonatal treatment with MSG (monosodium glutamate) on hypothalamo-pituitary-thyroid axis in adult male rats. Histol. Histopathol. 1993, 8, 731–734. [Google Scholar]
- Saito, M.; Minokoshi, Y.; Shimazu, T. Brown adipose tissue after ventromedial hypothalamic lesions in rats. Am. J. Physiol. 1985, 248, E20–E25. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Dallman, M.F. Hypothalamic obesity: Multiple routes mediated by loss of function in medial cell groups. Endocrinology 1999, 140, 4081–4088. [Google Scholar] [CrossRef]
- Fukushima, M.; Tokunaga, K.; Lupien, J.; Kemnitz, J.W.; Bray, G.A. Dynamic and static phases of obesity following lesions in PVN and VMH. Am. J. Physiol. 1987, 253, R523–R529. [Google Scholar] [CrossRef] [PubMed]
- Sims, J.S.; Lorden, J.F. Effect of paraventricular nucleus lesions on body weight, food intake and insulin levels. Behav. Brain Res. 1986, 22, 265–281. [Google Scholar] [CrossRef]
- An, J.J.; Liao, G.Y.; Kinney, C.E.; Sahibzada, N.; Xu, B. Discrete BDNF Neurons in the Paraventricular Hypothalamus Control Feeding and Energy Expenditure. Cell Metab. 2015, 22, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, L.; Lian, H.; Zhang, Y.; Tong, S.; Wang, Z. Dopamine receptor 2 downregulation and brain-derived neurotrophic factor upregulation in the paraventricular nucleus are correlated with brown adipose tissue thermogenesis in rats with bilateral substantia nigra lesions. J. Chem. Neuroanat. 2021, 117, 102016. [Google Scholar] [CrossRef]
- Franca, L.M.; Freitas, L.N.; Chagas, V.T.; Coelho, C.F.; Barroso, W.A.; Costa, G.C.; Silva, L.A.; Debbas, V.; Laurindo, F.R.; Paes, A.M. Mechanisms underlying hypertriglyceridemia in rats with monosodium L-glutamate-induced obesity: Evidence of XBP-1/PDI/MTP axis activation. Biochem. Biophys. Res. Commun. 2014, 443, 725–730. [Google Scholar] [CrossRef]
- Gloy, V.; Langhans, W.; Hillebrand, J.J.; Geary, N.; Asarian, L. Ovariectomy and overeating palatable, energy-dense food increase subcutaneous adipose tissue more than intra-abdominal adipose tissue in rats. Biol. Sex Differ. 2011, 2, 6. [Google Scholar] [CrossRef]
- Wang, W.; Yang, Q.; Zhou, C.; Jiang, H.; Sun, Y.; Wang, H.; Luo, X.; Wang, Z.; Zhang, J.; Wang, K.; et al. Transcriptomic changes in the hypothalamus of ovariectomized mice: Data from RNA-seq analysis. Ann. Anat.-Anat. Anz. 2022, 241, 151886. [Google Scholar] [CrossRef]
- Romero-Picó, A.; Novelle, M.G.; Al-Massadi, O.; Beiroa, D.; Tojo, M.; Heras, V.; Ruiz-Pino, F.; Senra, A.; López, M.; Blouet, C.; et al. Kappa-Opioid Receptor Blockade Ameliorates Obesity Caused by Estrogen Withdrawal via Promotion of Energy Expenditure through mTOR Pathway. Int. J. Mol. Sci. 2022, 23, 3118. [Google Scholar] [CrossRef]
- de Moura e Dias, M.; dos Reis, S.A.; da Conceição, L.L.; Sediyama, C.M.N.O.; Pereira, S.S.; de Oliveira, L.L.; Gouveia Peluzio, M.C.; Martinez, J.A.; Milagro, F.I. Diet-induced obesity in animal models: Points to consider and influence on metabolic markers. Diabetol. Metab. Syndr. 2021, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Panchal, S.K.; Brown, L. Rodent models for metabolic syndrome research. J. Biomed. Biotechnol. 2011, 2011, 351982. [Google Scholar] [CrossRef] [PubMed]
- Glastras, S.J.; Chen, H.; Teh, R.; McGrath, R.T.; Chen, J.; Pollock, C.A.; Wong, M.G.; Saad, S. Mouse Models of Diabetes, Obesity and Related Kidney Disease. PLoS ONE 2016, 11, e0162131. [Google Scholar] [CrossRef]
- Yang, Y.; Smith, D.L., Jr.; Keating, K.D.; Allison, D.B.; Nagy, T.R. Variations in body weight, food intake and body composition after long-term high-fat diet feeding in C57BL/6J mice. Obesity 2014, 22, 2147–2155. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.; Chandrasekera, P.C.; Barnard, N.D. You are what you eat, or are you? The challenges of translating high-fat-fed rodents to human obesity and diabetes. Nutr. Diabetes 2014, 4, e135. [Google Scholar] [CrossRef]
- Eng, J.M.; Estall, J.L. Diet-Induced Models of Non-Alcoholic Fatty Liver Disease: Food for Thought on Sugar, Fat, and Cholesterol. Cells 2021, 10, 1805. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Chen, D.; Zhang, K.; Yu, B. A high-amylopectin diet caused hepatic steatosis associated with more lipogenic enzymes and increased serum insulin concentration. Br. J. Nutr. 2011, 106, 1470–1475. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Fairus, A.; Ima-Nirwana, S. Animal models of metabolic syndrome: A review. Nutr. Metab. 2016, 13, 65. [Google Scholar] [CrossRef]
- Bastias-Perez, M.; Serra, D.; Herrero, L. Dietary Options for Rodents in the Study of Obesity. Nutrients 2020, 12, 3234. [Google Scholar] [CrossRef]
- Virtuoso, A.; Tveden-Nyborg, P.; Schou-Pedersen, A.M.V.; Lykkesfeldt, J.; Müller, H.K.; Elfving, B.; Sørensen, D.B. A Long-Term Energy-Rich Diet Increases Prefrontal BDNF in Sprague-Dawley Rats. Nutrients 2022, 14, 126. [Google Scholar] [CrossRef]
- Matuszewska, J.; Zalewski, T.; Klimaszyk, A.; Ziarniak, K.; Jurga, S.; Chmurzynska, A.; Sliwowska, J.H. Mothers’ cafeteria diet induced sex-specific changes in fat content, metabolic profiles, and inflammation outcomes in rat offspring. Sci. Rep. 2021, 11, 18573. [Google Scholar] [CrossRef] [PubMed]
- Baccouche, B.; Benlarbi, M.; Barber, A.J.; Chaouacha-Chekir, R.B. Short-Term Administration of Astaxanthin Attenuates Retinal Changes in Diet-Induced Diabetic Psammomys obesus. Curr. Eye Res. 2018, 43, 1177–1189. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, R.; Walder, K.R.; Hagemeyer, C.E.; Kanwar, J.R. Psammomys obesus: A Natural Diet-Controlled Model for Diabetes and Cardiovascular Diseases. Curr. Atheroscler. Rep. 2018, 20, 46. [Google Scholar] [CrossRef]
- Shafrir, E.; Ziv, E.; Kalman, R. Nutritionally induced diabetes in desert rodents as models of type 2 diabetes: Acomys cahirinus (spiny mice) and Psammomys obesus (desert gerbil). ILAR J. 2006, 47, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Chaouad, B.; Moudilou, E.N.; Ghoul, A.; Zerrouk, F.; Moulahoum, A.; Othmani-Mecif, K.; Cherifi, M.E.H.; Exbrayat, J.M.; Benazzoug, Y. Hyperhomocysteinemia and myocardial remodeling in the sand rat, Psammomys obesus. Acta Histochem. 2019, 121, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.X.; Harn, H.I.; Ou, K.L.; Lei, M.; Chuong, C.M. Comparative regenerative biology of spiny (Acomys cahirinus) and laboratory (Mus musculus) mouse skin. Exp. Dermatol. 2019, 28, 442–449. [Google Scholar] [CrossRef]
- Maden, M.; Brant, J.O.; Rubiano, A.; Sandoval, A.G.W.; Simmons, C.; Mitchell, R.; Collin-Hooper, H.; Jacobson, J.; Omairi, S.; Patel, K. Perfect chronic skeletal muscle regeneration in adult spiny mice, Acomys cahirinus. Sci. Rep. 2018, 8, 8920. [Google Scholar] [CrossRef]
- Simkin, J.; Gawriluk, T.R.; Gensel, J.C.; Seifert, A.W. Macrophages are necessary for epimorphic regeneration in African spiny mice. eLife 2017, 6, e24623. [Google Scholar] [CrossRef]
- Wong, W.; Kim, A.; Monaghan, J.R.; Seifert, A.W.; Maden, M.; Crane, J.D. Spiny mice (Acomys) exhibit attenuated hallmarks of aging and rapid cell turnover after UV exposure in the skin epidermis. PLoS ONE 2020, 15, e0241617. [Google Scholar] [CrossRef]
- Choudhary, A.; Ibdah, J.A. Animal models in today’s translational medicine world. Mo. Med. 2013, 110, 220–222. [Google Scholar]
- Loos, R.J.F.; Yeo, G.S.H. The genetics of obesity: From discovery to biology. Nat. Rev. Genet. 2022, 23, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Sinasac, D.S.; Riordan, J.D.; Spiezio, S.H.; Yandell, B.S.; Croniger, C.M.; Nadeau, J.H. Genetic control of obesity, glucose homeostasis, dyslipidemia and fatty liver in a mouse model of diet-induced metabolic syndrome. Int. J. Obes. 2016, 40, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Flores-Dorantes, M.T.; Díaz-López, Y.E.; Gutiérrez-Aguilar, R. Environment and Gene Association With Obesity and Their Impact on Neurodegenerative and Neurodevelopmental Diseases. Front. Neurosci. 2020, 14, 863. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Pound, P.; Ritskes-Hoitinga, M. Is it possible to overcome issues of external validity in preclinical animal research? Why most animal models are bound to fail. J. Transl. Med. 2018, 16, 304. [Google Scholar] [CrossRef] [PubMed]
- Bernardis, L.L. Prediction of carcass fat, water and lean body mass from Lee’s "nutritive ratio" in rats with hypothalamic obesity. Experientia 1970, 26, 789–790. [Google Scholar] [CrossRef]
- Gargiulo, S.; Gramanzini, M.; Megna, R.; Greco, A.; Albanese, S.; Manfredi, C.; Brunetti, A. Evaluation of growth patterns and body composition in C57Bl/6J mice using dual energy X-ray absorptiometry. BioMed Res. Int. 2014, 2014, 253067. [Google Scholar] [CrossRef]
- Nixon, J.P.; Zhang, M.; Wang, C.; Kuskowski, M.A.; Novak, C.M.; Levine, J.A.; Billington, C.J.; Kotz, C.M. Evaluation of a quantitative magnetic resonance imaging system for whole body composition analysis in rodents. Obesity 2010, 18, 1652–1659. [Google Scholar] [CrossRef]
- Francisqueti, F.V.; Nascimento, A.F.; Minatel, I.O.; Dias, M.C.; Luvizotto, R.A.M.; Berchieri-Ronchi, C.; Ferreira, A.L.A.; Correa, C.R. Metabolic syndrome and inflammation in adipose tissue occur at different times in animals submitted to a high-sugar/fat diet. J. Nutr. Sci. 2017, 6, e41. [Google Scholar] [CrossRef]
- Leopoldo, A.S.; Lima-Leopoldo, A.P.; Nascimento, A.F.; Luvizotto, R.A.; Sugizaki, M.M.; Campos, D.H.; da Silva, D.C.; Padovani, C.R.; Cicogna, A.C. Classification of different degrees of adiposity in sedentary rats. Braz. J. Med. Biol. Res. 2016, 49, e5028. [Google Scholar] [CrossRef]
- Arantes-Rodrigues, R.; Henriques, A.; Pires, M.J.; Colaco, B.; Calado, A.M.; Rema, P.; Colaco, A.; Fernandes, T.; De la Cruz, P.L.; Lopes, C.; et al. High doses of olive leaf extract induce liver changes in mice. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2011, 49, 1989–1997. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Begum, S.; Oh, D.S.; Wee, A.J.; Yun, B.S.; Sung, C.K. Ameliorating Effect of Mycoleptodonoides aitchisonii on High-fat Diet-induced Obese Mice. Prev. Nutr. Food Sci. 2014, 19, 69–74. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lee, M.O. Determination of the surface area of the white rat with its application to the expression of metabolic results. Am. J. Physiol.-Leg. Content 1929, 89, 24–33. [Google Scholar] [CrossRef]
- Simson, E.L.; Gold, R.M. The lee obesity index vindicated? Physiol. Behav. 1982, 29, 371–376. [Google Scholar] [CrossRef]
- Ayala, J.E.; Samuel, V.T.; Morton, G.J.; Obici, S.; Croniger, C.M.; Shulman, G.I.; Wasserman, D.H.; McGuinness, O.P.; Consortium, N.I.H.M.M.P.C. Standard operating procedures for describing and performing metabolic tests of glucose homeostasis in mice. Dis. Models Mech. 2010, 3, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Benede-Ubieto, R.; Estevez-Vazquez, O.; Ramadori, P.; Cubero, F.J.; Nevzorova, Y.A. Guidelines and Considerations for Metabolic Tolerance Tests in Mice. Diabetes Metab. Syndr. Obes. 2020, 13, 439–450. [Google Scholar] [CrossRef]
- Bowe, J.E.; Franklin, Z.J.; Hauge-Evans, A.C.; King, A.J.; Persaud, S.J.; Jones, P.M. Metabolic phenotyping guidelines: Assessing glucose homeostasis in rodent models. J. Endocrinol. 2014, 222, G13–G25. [Google Scholar] [CrossRef]
- Jensen, T.L.; Kiersgaard, M.K.; Sorensen, D.B.; Mikkelsen, L.F. Fasting of mice: A review. Lab. Anim. 2013, 47, 225–240. [Google Scholar] [CrossRef]
- Nagy, C.; Einwallner, E. Study of In Vivo Glucose Metabolism in High-fat Diet-fed Mice Using Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT). J. Vis. Exp. 2018, 7, 56672. [Google Scholar] [CrossRef]
- Parasuraman, S.; Raveendran, R. Measurement of invasive blood pressure in rats. J. Pharm. Pharm. 2012, 3, 172–177. [Google Scholar] [CrossRef]
- Plehm, R.; Barbosa, M.E.; Bader, M. Animal models for hypertension/blood pressure recording. In Cardiovascular Disease, 2006/11/07 ed.; Wang, Q., Ed.; Methods in Molecular Medicine; Humana Press: Totowa, NJ, USA, 2006; Volume 129, pp. 115–126. [Google Scholar]
- Sikaris, K.A. The clinical biochemistry of obesity. Clin. Biochem. Rev. 2004, 25, 165–181. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Models | Advantages | Disadvantages |
|---|---|---|
| Monogenic |
|
|
| Polygenic |
|
|
| Genetically modified |
|
|
| Diet-induced |
|
|
| Chemical or surgical |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, T.; Castro-Ribeiro, C.; Lemos, S.; Ferreira, T.; Nascimento-Gonçalves, E.; Rosa, E.; Oliveira, P.A.; Antunes, L.M. Murine Models of Obesity. Obesities 2022, 2, 127-147. https://doi.org/10.3390/obesities2020012
Martins T, Castro-Ribeiro C, Lemos S, Ferreira T, Nascimento-Gonçalves E, Rosa E, Oliveira PA, Antunes LM. Murine Models of Obesity. Obesities. 2022; 2(2):127-147. https://doi.org/10.3390/obesities2020012
Chicago/Turabian StyleMartins, Tânia, Catarina Castro-Ribeiro, Sílvia Lemos, Tiago Ferreira, Elisabete Nascimento-Gonçalves, Eduardo Rosa, Paula Alexandra Oliveira, and Luís Miguel Antunes. 2022. "Murine Models of Obesity" Obesities 2, no. 2: 127-147. https://doi.org/10.3390/obesities2020012
APA StyleMartins, T., Castro-Ribeiro, C., Lemos, S., Ferreira, T., Nascimento-Gonçalves, E., Rosa, E., Oliveira, P. A., & Antunes, L. M. (2022). Murine Models of Obesity. Obesities, 2(2), 127-147. https://doi.org/10.3390/obesities2020012