
Effects of Deadwood on Macroinvertebrate Assemblages in Three Sand-Type Lowland Streams



Abstract
1. Introduction
- Deadwood (DW) serves as a food source for MI, increases habitat and species diversity, and individual density [6]. Compared to sand, wood and detritus promote area-specific assemblages by MI in general and by aquatic insects specifically.
- The MI assemblages of detritus are higher in wood sections compared to the control sections. In contrast wood has no effect on MI assemblages of the sand of wood sections, so the sand of wood sections is not significantly different from that of control sections.
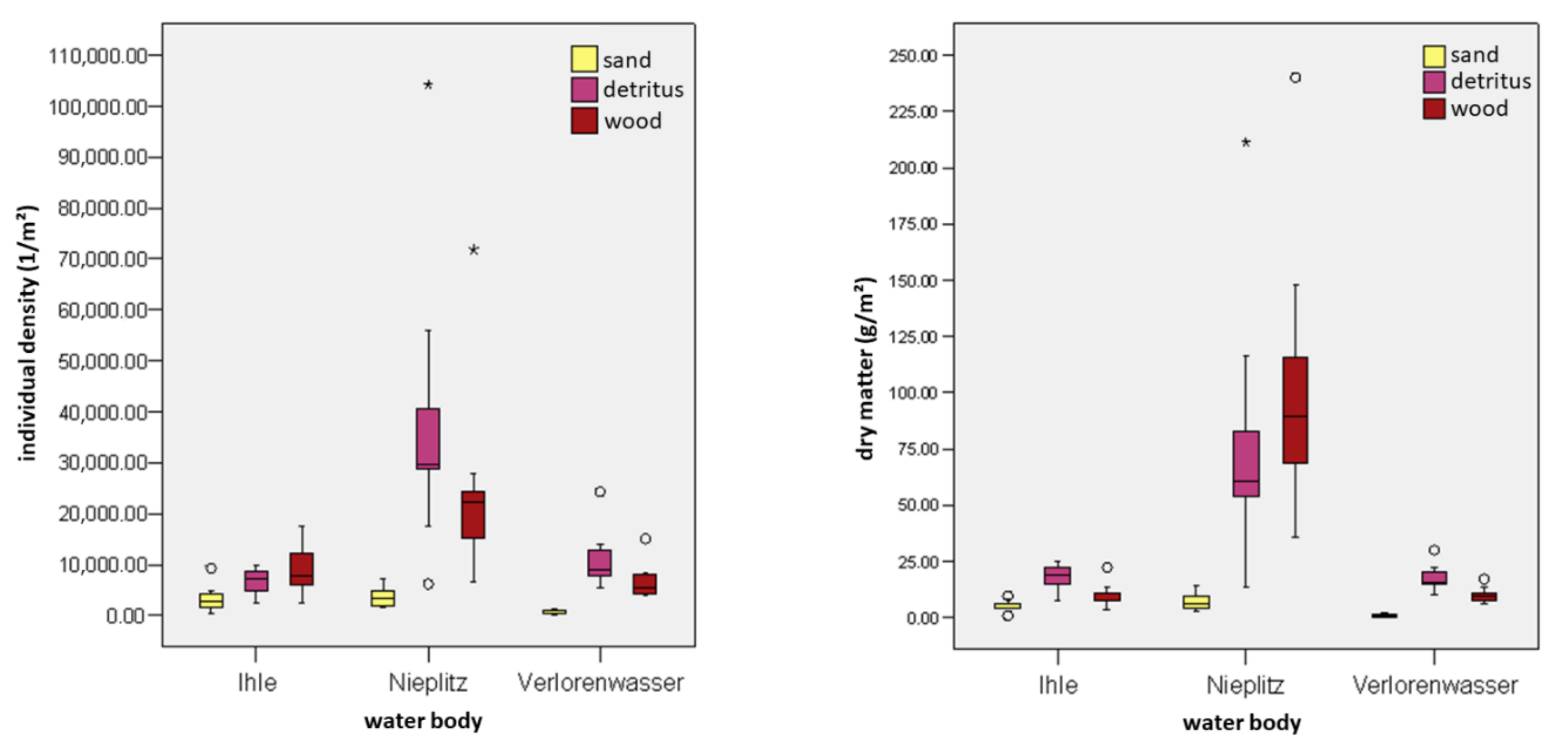
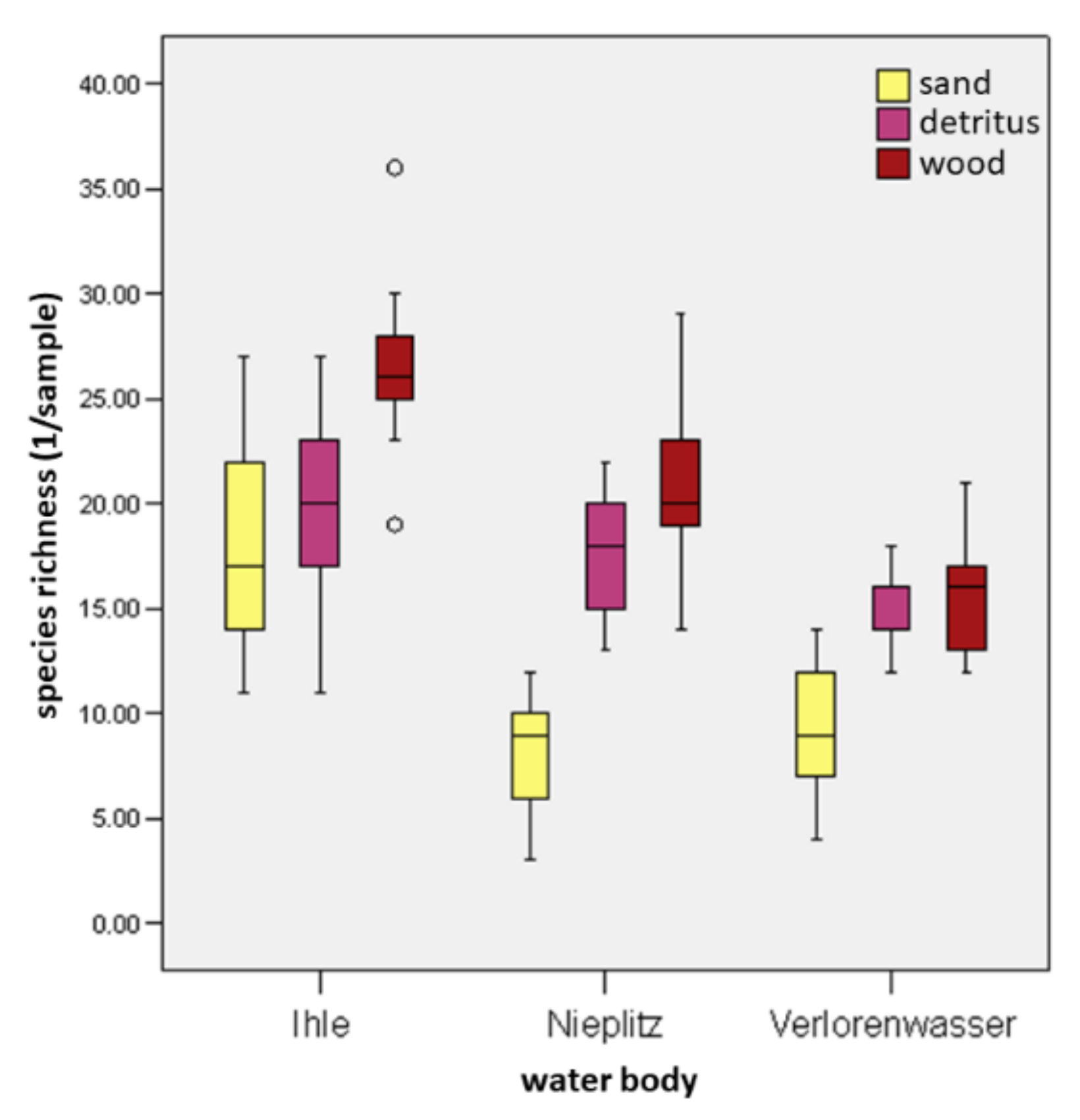
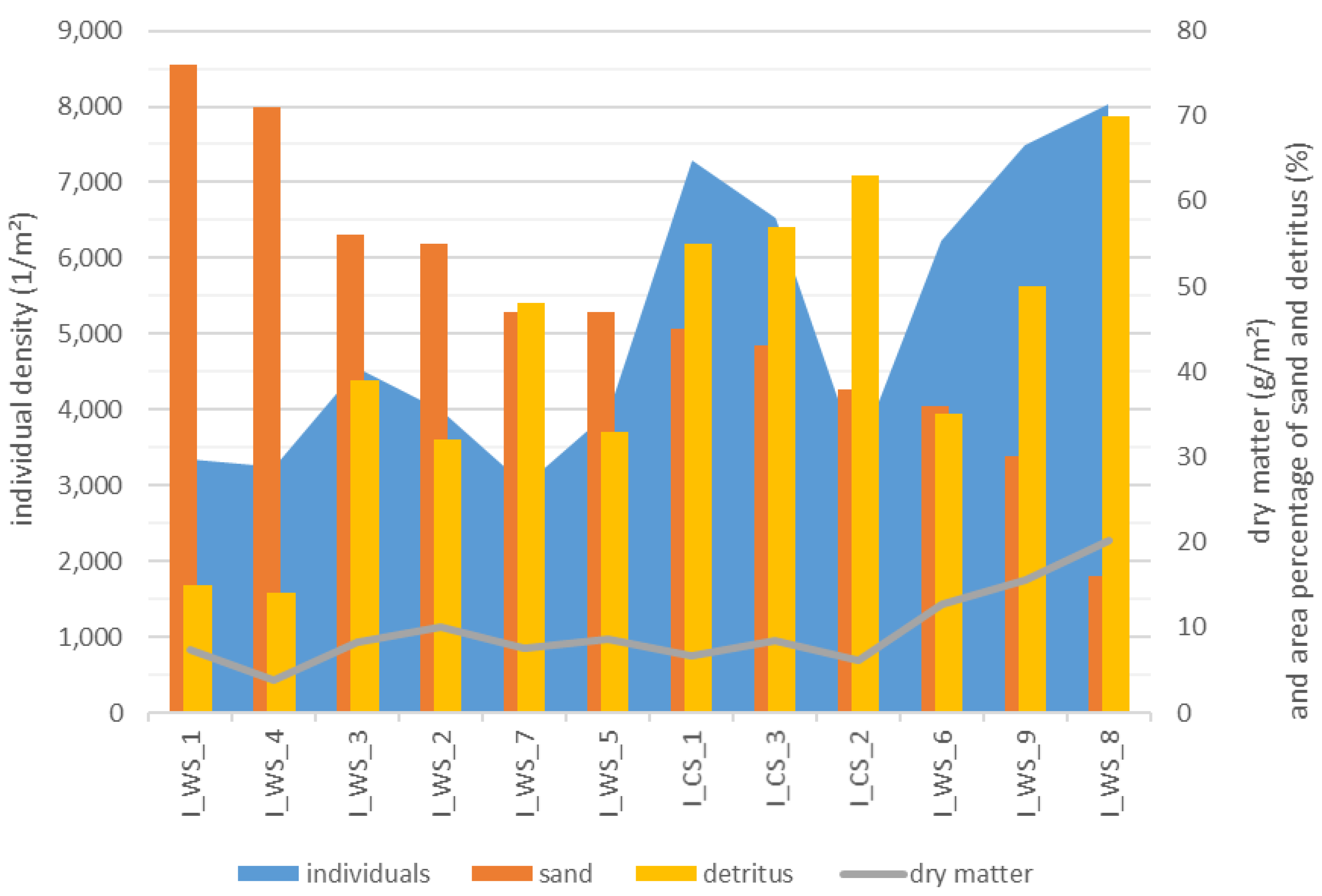
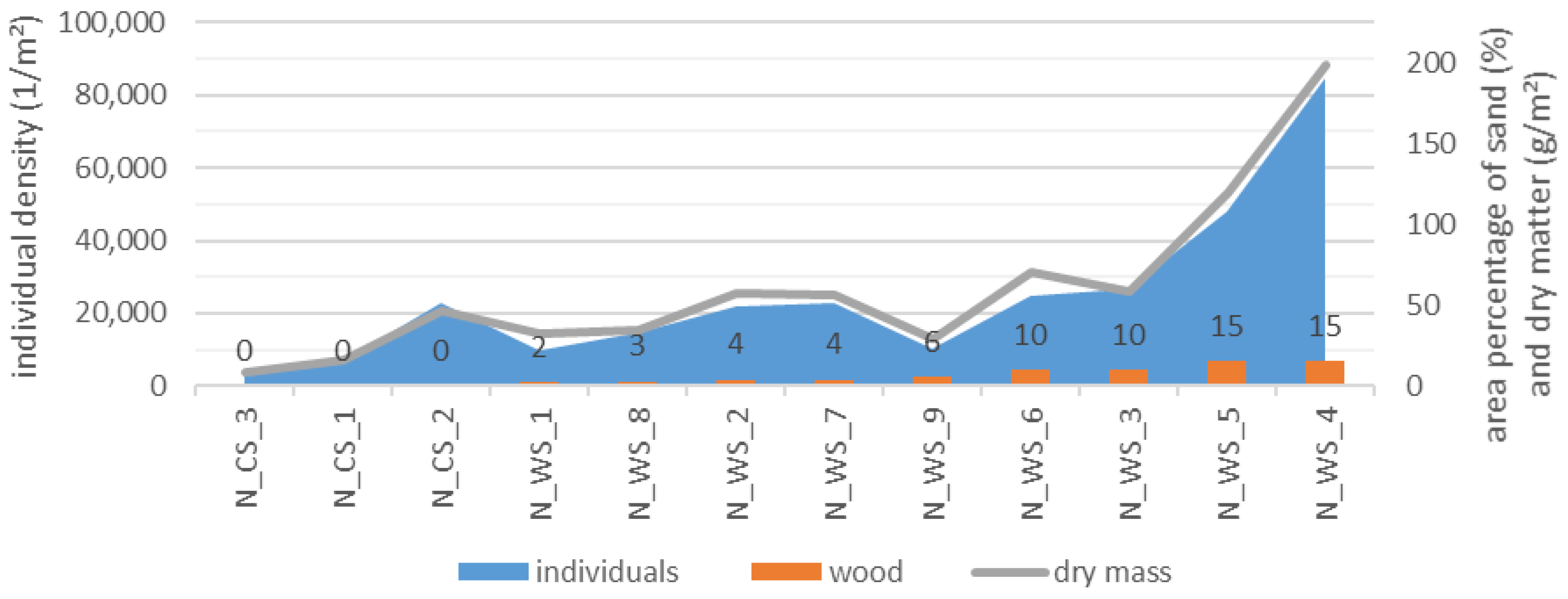
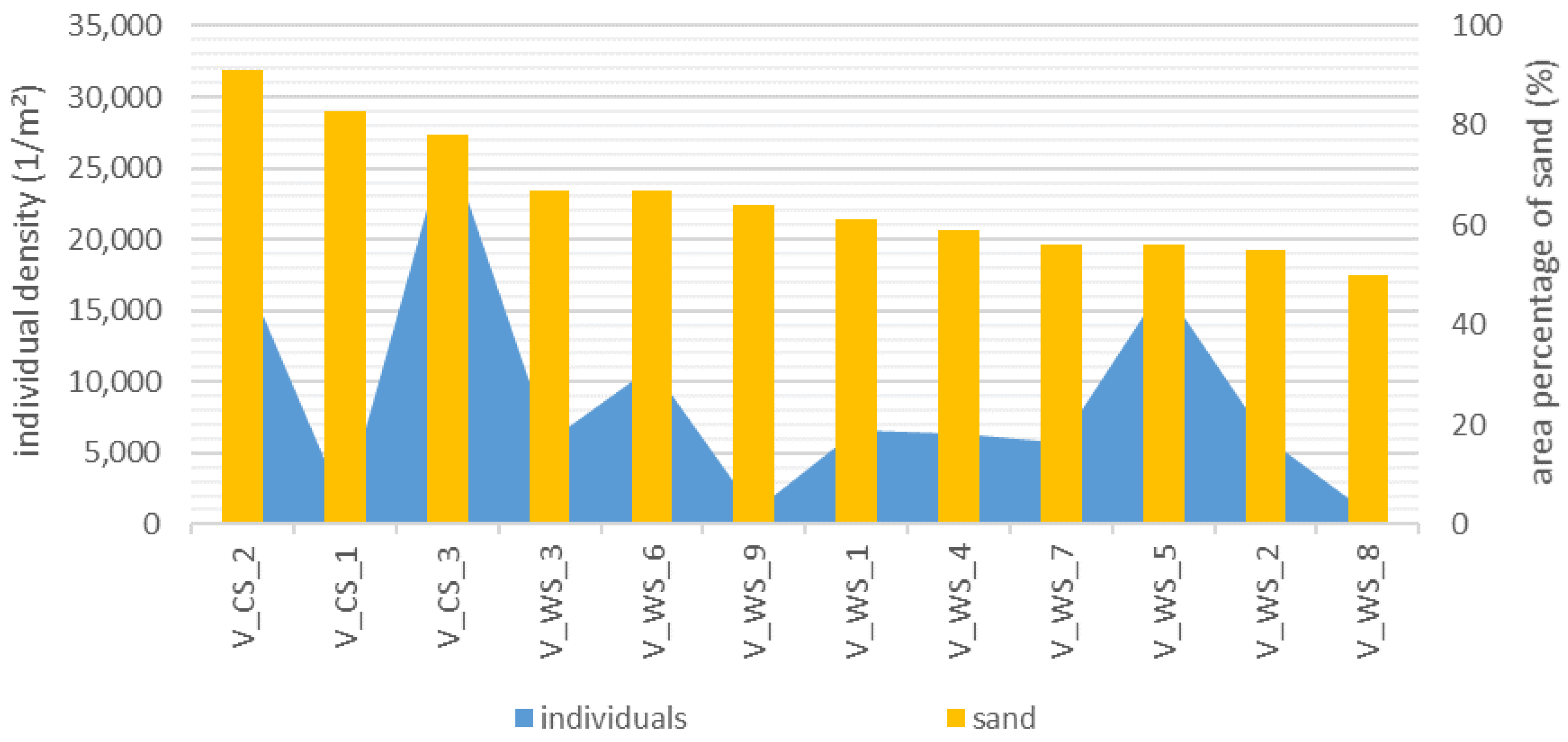
- The percentage of substrates per unit area directly influences the individual density and the dry matter of a water body section. Each section of wood and detritus promotes the total MI assemblages in a stream (including all substrates), while sand inhibits species richness and density of individuals.
- Wood promotes MI assemblages in a water segment (including detritus, sand and gravel) in contrast to a section of water without wood (control section).
2. Material and Methods
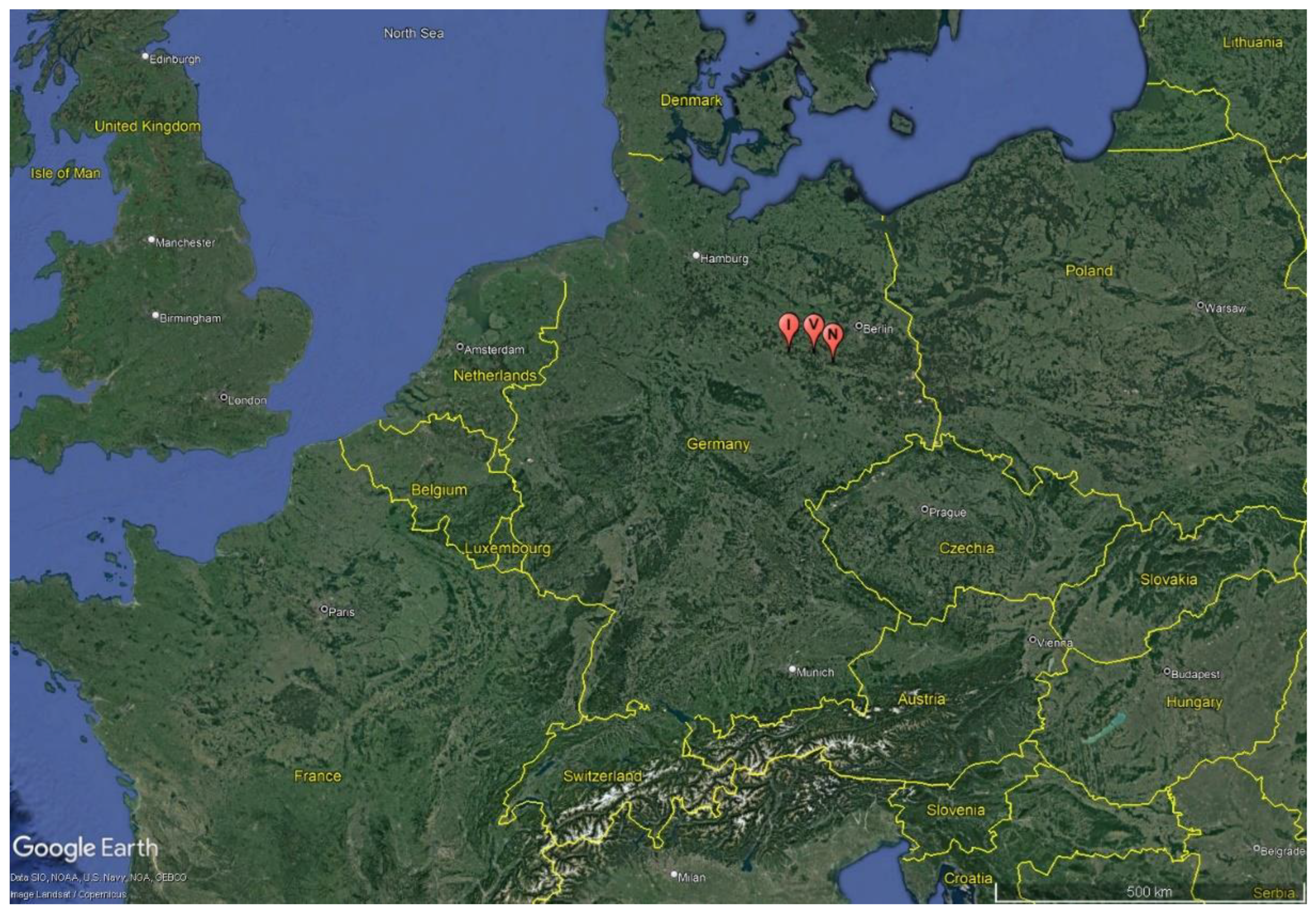
2.1. Project Area
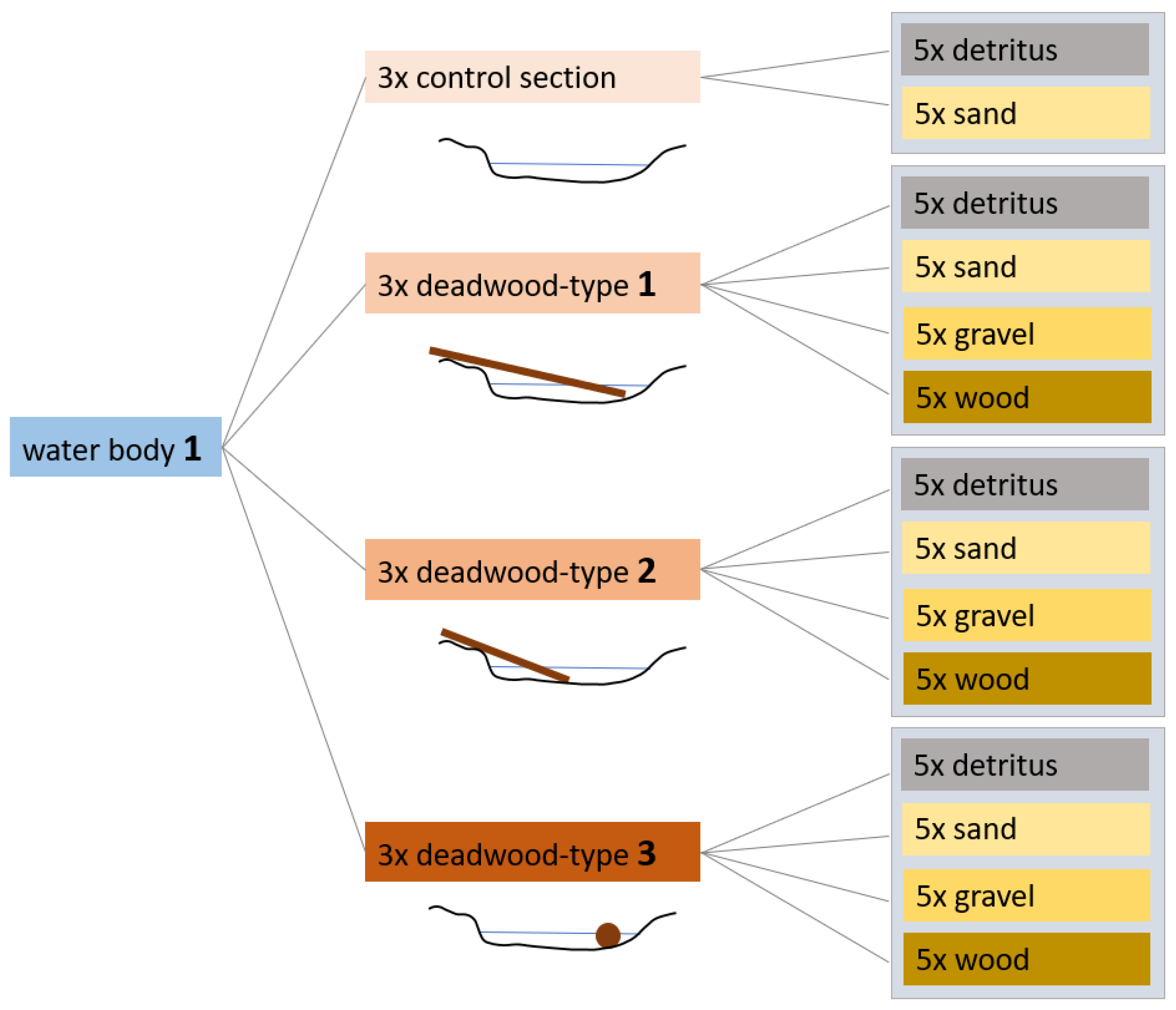
2.2. Methods
2.2.1. Sampling and Taxonomies
2.2.2. Measurement of Dry Matter
2.2.3. Statistics
3. Results
3.1. Stream Bottom
3.2. MI Colonization in the Streams
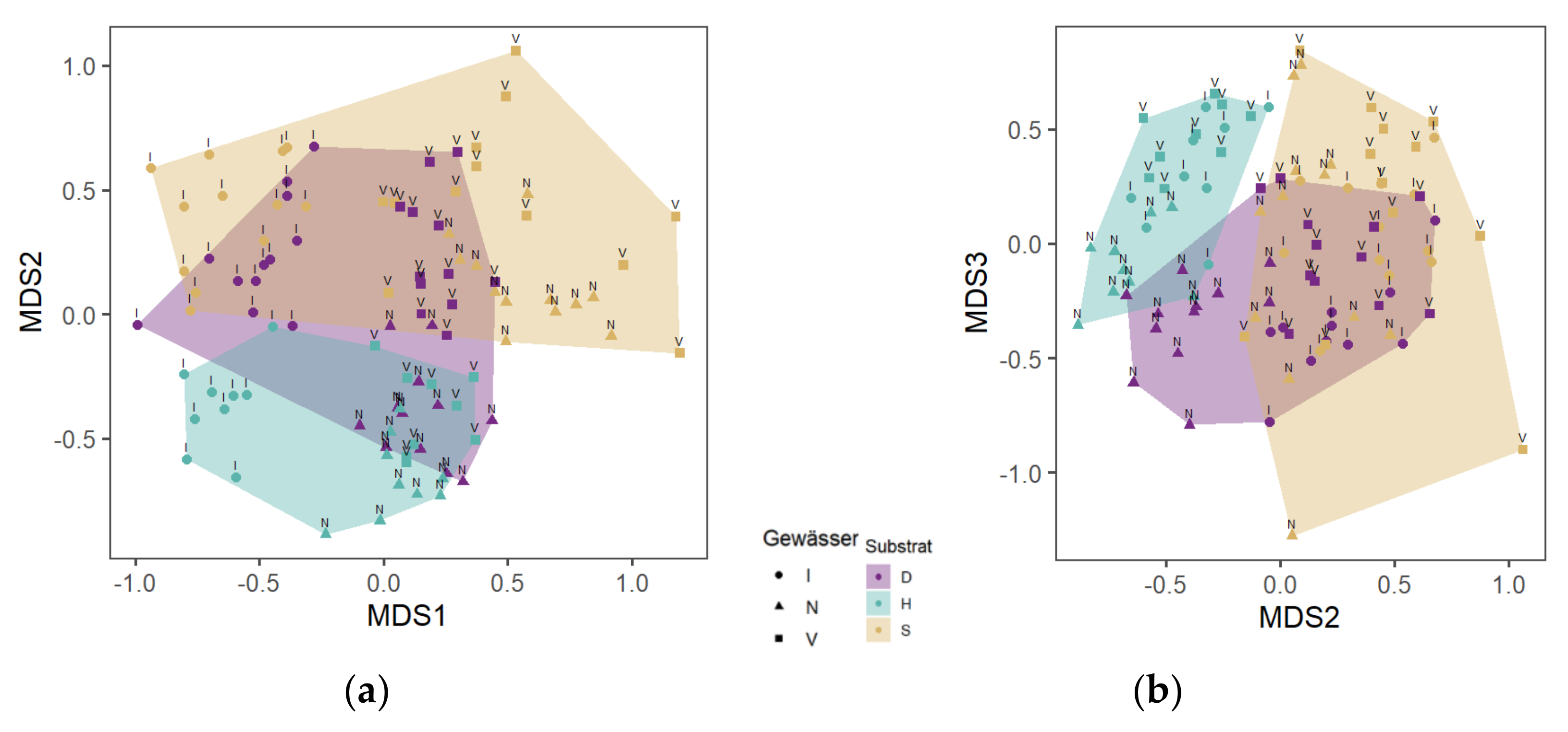
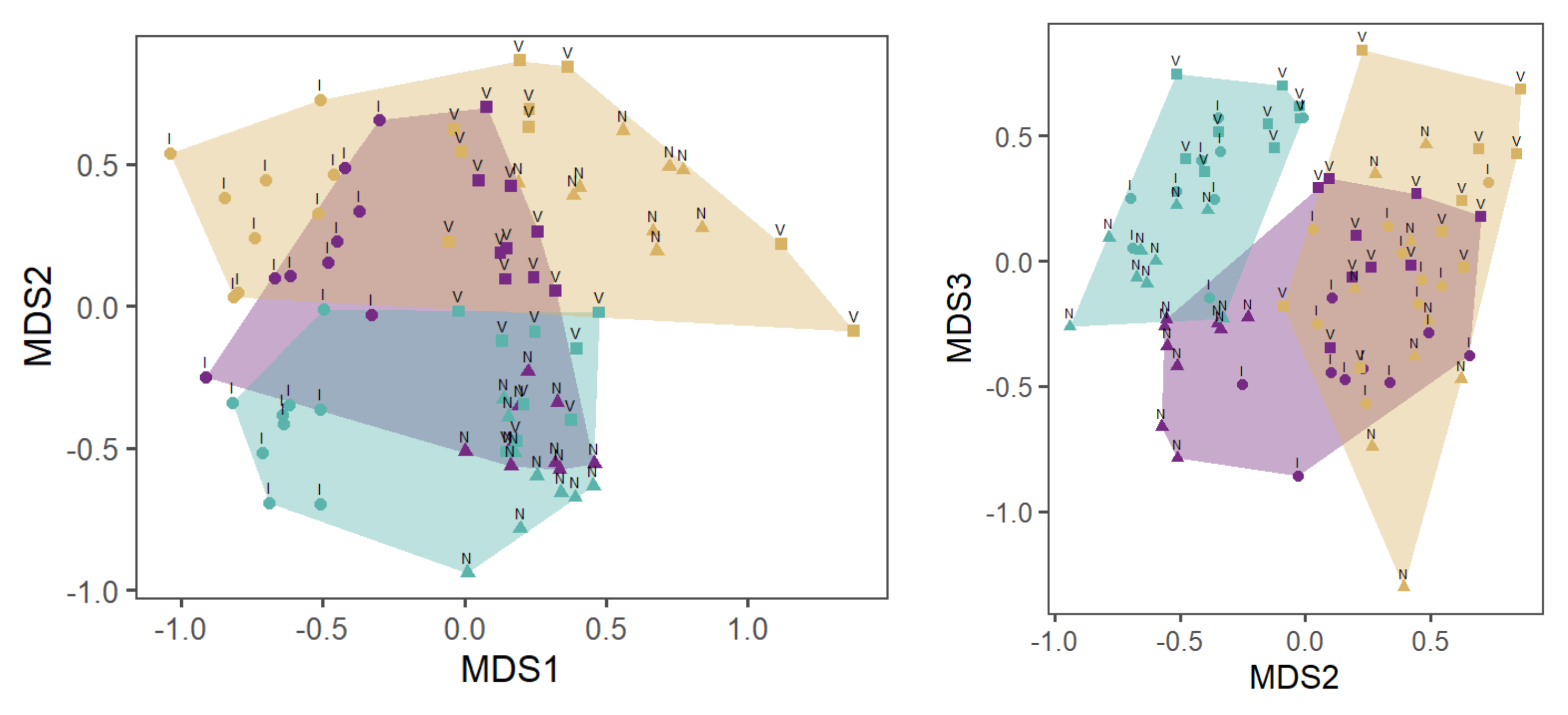
3.3. Species Composition of MI in Dependence on Substrates
3.3.1. Wood and Control Sections
3.3.2. Wood Sections
3.4. Colonization of MI and Insects in Dependence on Substrates
3.5. Relationship between the MI Population and the Area Percentage of Substrates
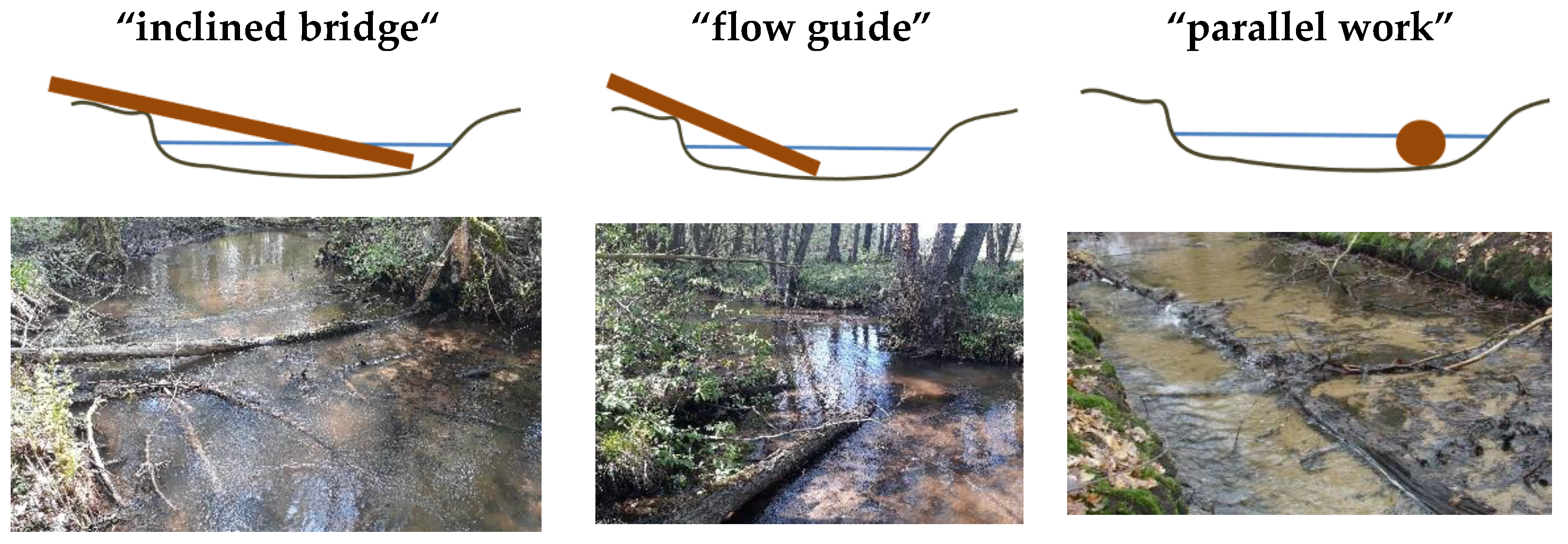
3.6. Wood Structure Types
4. Discussion
4.1. Influence of Deadwood and Restoration on the MI-Fauna
4.2. Effect of Substrates on MI Populations
4.3. Substrates in WS and CS
4.4. Comparison of Stream Sections in WS and CS
4.5. Wood Texture Types
5. Conclusions and Recommendations for Improvement Measures
- Make surrounding land use more MI friendly: Riparian strips should be identified and secured so that deadwood can be brought in a natural way. This input can promote detritus formation and MI colonization, particularly in sections that have little or no deadwood. Care should be taken to ensure that the formation of colonization-promoting substrates such as detritus and wood is favored but sand is not completely covered.
- Create diverse water body structures: Settlement by MI is dependent on the available surface area of wood and less on volume. Thus, when present in sufficient quantities, branches (0.05 to 0.1 m in diameter) and coarse organic material, such as large logs, have the potential as additional microhabitats.
- Considering the surface of deadwood: More individuals would be found under the bark of older deadwood because they are protected from drifting. When putting in deadwood, care should be taken to ensure that deadwood with bark is among.
- Use of deadwood also in Heavily Modified Water Bodies: The effect of introduced deadwood in unnatural streams compared to near-natural streams with natural deadwood is similar, the species richness did not differ significantly from each other.
- Further development of monitoring: In order to identify problems and to check effectiveness, monitoring and further research are of great importance for the promotion of colonization by MI, insects and sensitive species by means of deadwood.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed]
- Hornberg, C.; Niekisch, M.; Calliess, C.; Kemfert, C.; Lucht, W.; Messari-Becker, L.; Rotter, V.S. For a Surface-Effective Insect Protection. Opinion; Scientific Advisory Board for Biodiversity and Genetic Resources at the Federal Ministry of Food and Agriculture, Ed.; German Advisory Council on the Environment (SRU): Berlin, Germany, 2016; Available online: https://www.umweltrat.de/SharedDocs/Downloads/DE/04_Stellungnamen/2016_2020/2018_10_AS_Insektenschutz.pdf?__blob=publicationFile&v=19 (accessed on 4 August 2020).
- Liess, M.; Liebmann, L.; Vormeier, P.; Weisner, O.; Altenburger, R.; Borchardt, D.; Brack, W.; Chatzinotas, A.; Escher, B.; Foit, K.; et al. Pesticides are the dominant stressors for vulnerable insects in lowland streams. Water Res. 2021, 201, 117262. [Google Scholar] [CrossRef] [PubMed]
- Seidel, M. Naturnaher Einsatz von Holz zur Entwicklung von Fließgewässern im Norddeutschen Tiefland. Schriftenreihe des Instituts für Wasserwirtschaft und Ökotechnologie; Ettmer, B., Lüderitz, V., Eds.; Shaker Verlag GmbH: Aachen, Germany, 2018. [Google Scholar] [CrossRef]
- UBA. Umweltbundesamt (2019): Gewässertyp des Jahres 2018—Sandiger Tieflandbach. 2019. Available online: https://www.umweltbundesamt.de/themen/wasser/gewaessertyp-des-jahres/gewaessertyp-des-jahres-2018-sandiger-tieflandbach#kurzbeschreibung (accessed on 10 November 2020).
- Röthele, I.; Mertens, M.; Küry, D. Erhebung Biomasse Makrozoobenthos 2014 Violenbach, Orisbach, Ergolz, Frenke, Hintere Frenke, Vordere Frenke. 2014. Available online: https://www.baselland.ch/politik-und-behorden/direktionen/volkswirtschafts-und-gesundheitsdirektion/jagd-und-fischerei/fischereiwesen/fischfauna-untersuchungsberichte/downloads/erhebung-biomasse-makrozoobenthos_2014.pdf/@@download/file/erhebung-biomasse-makrozoobenthos_2014.pdf (accessed on 10 May 2020).
- Neuhaus, V.; Mende, M. Engineered Large Wood Structures in Stream Restoration Projects in Switzerland: Practice-Based Experiences. Water 2021, 13, 2520. [Google Scholar] [CrossRef]
- Google. n.d. Locations of Ihle, Nieplitz and Verlorenwasser. Data SIO, NOAA, U.S. Navy, NGA, GEBCA. Image Landsat/Copernicus. Available online: https://earth.google.com/web/search/ihle/@49.42246021,8.12620422,-57.14636488a,1828973.53736222d,35y,-0.32186261h,23.21376508t,0r/data=CigiJgokCZXaZMslE0VAEYEcRHJFEEVAGQC0GiHTVQRAIauPEd9hMQRA (accessed on 26 December 2022).
- UBA. Umweltbundesamt. Fließgewässertyp des Jahres. GIS-Karte. 2016. Available online: https://gis.uba.de/maps/resources/apps/gdj/index.html?lang=de (accessed on 24 November 2020).
- Pottgiesser, T.; Sommerhäuser, M. Aktualisierung der Steckbriefe der Bundesdeutschen Fließgewässertypen (Teil A) und Ergänzung der Steckbriefe der Deutschen Fließgewässertypen um Typspezifische Referenzbedingungen und Bewertungsverfahren aller Qualitätselemente (Teil B). Essen. 2008. Available online: http://gewaesser-bewertung.de/files/steckbriefe_fliessgewaessertypen_april2008.pdf (accessed on 30 April 2020).
- Gerhard, M.; Reich, M. Totholz in Fließgewässern: Empfehlungen zur Gewässerentwicklung; Gemeinnützige Fortbildungsgesellschaft für Wasserwirtschaft und Landschaftsentwicklung (GFG) mbH: Mainz, Germany; WBW Fortbildungsgesellschaft für Gewässerentwicklung mbH: Karlsruhe, Germany, 2001; pp. 12–19. Available online: https://wbw-fortbildung.de/sites/default/files/2021-10/Totholz%20in%20Flie%C3%9Fgew%C3%A4ssern%20-%20Empfehlung%20zur%20Gew%C3%A4sserentwicklung.pdf (accessed on 4 August 2020).
- Seidel, M.; Brunke, M. Impulsgebende Maßnahmen in Tieflandbächen Schleswig-Holsteins—Wirkung und mögliche Einbauvarianten für eine naturnähere Gestaltung. Wasser Wirtsch. 2015, 12, 55–60. [Google Scholar] [CrossRef]
- Bellmann, H. Der Kosmos Libellenführer: Die Arten Mitteleuropas Sicher Bestimmen; Franckh-Kosmos Verlags-GmbH &Co. KG: Stuttgart, Germany, 2007. [Google Scholar]
- Eiseler, B. Bildbestimmungshilfen für die Eintagsfliegenlarven der deutschen Mittelgebirge und des Tieflandes. In Lauterbornia. Internationale Zeitschrift für Faunistik und Floristik der Binnengewässer Europas; Heft 53; Mauch, E., Ed.; Kessler Verlagsdruckerei: Dinkelscherben, Germany, 2005. [Google Scholar]
- Eiseler, B. Bestimmungshilfen—Makrozoobenthos (1). Arbeitsblatt 14. In Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen (Ed.): Taxonomie für die Praxis; LANUV: Recklinghausen, Germany, 2010. [Google Scholar]
- Eiseler, B.; Hess, M. Bestimmungshilfen—Makrozoobenthos (2). Arbeitsblatt 20. In Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen (Ed.): Taxonomie für die Praxis; LANUV: Recklinghausen, Germany, 2013. [Google Scholar]
- Glöer, P. Süßwassermollusken. Ein Bestimmungsschlüssel für die Bundesrepublik Deutschland (14. Auflage); Deutscher Jugendbund für Naturbeobachtung, Ed.; SENSER-DRUCK: Augsburg, Germany, 2015. [Google Scholar]
- Waringer, J.; Graf, W. Atlas der Österreichischen Köcherfliegenlarven unter Einschluß der Angrenzenden Gebiete; Facultas Universitätsverlag: Vienna, Austria, 1997. [Google Scholar]
- Jiang, X.; Song, Z.; Xiong, J.; Xie, Z. Can excluding non-insect taxa from stream macroinvertebrate surveys enhance the sensitivity of taxonomic distinctness indices to human disturbance? Ecol. Indic. 2014, 41, 175–182. [Google Scholar] [CrossRef]
- Chapman, M.G.; Underwood, A.J. Ecological patterns in multivariate assemblages: Information and interpretation of negative values in ANOSIM tests. Mar. Ecol. Prog. Ser. 1999, 180, 257–265. [Google Scholar]
- Poledniková, Z.; Galia, T. Ecosystem Services of Large Wood: Mapping the Research Gap. Water 2021, 13, 2594. [Google Scholar] [CrossRef]
- Roni, P.; Beechie, T.; Pess, G.; Hanson, K. Wood placement in river restoration: Fact, fiction, and future direction. Can. J. Fish. Aquat. Sci. 2015, 72, 466–478. [Google Scholar] [CrossRef]
- Whiteway, S.L.; Biron, P.M.; Zimmermann, A.; Venter, O.; Grant, J.W. Do in-stream restoration structures enhance salmonid abundance? A meta-analysis. Can. J. Fish. Aquat. Sci. 2010, 67, 831–841. [Google Scholar] [CrossRef]
- Lo, H.W.; Smith, M.; Klaar, M.; Woulds, C. Potential secondary effects of in-stream wood structures installed for natural flood management: A conceptual model. WIREs Water 2021, 8, e1546. [Google Scholar] [CrossRef]
- Mutz, M. Influences of Woody Debris on Flow Patterns and Channel Morphology in a Low Energy, Sand-Bed Stream Reach. Int. Rev. Hydrobiol. 2000, 1, 107–121. [Google Scholar] [CrossRef]
- Seidel, M.; Li, F.; Langheinrich, U.; Gersberg, R.M.; Lüderitz, V. Self-dynamics as a driver for restoration success in a lowland stream reach. Limnologica 2021, 88, 125873. [Google Scholar] [CrossRef]
- Seidel, M.; Li, F.; Winking, C.; Sommerhäuser, M.; Lüderitz, V. Should We Sample More than Required by the European Water Framework Directive? Case study: Emscher Catchment. CLEAN—Soil Air Water 2022, 50, 2000391. [Google Scholar] [CrossRef]
- Lüderitz, V.; Jüpner, R.; Müller, S.; Feld, C.K. Renaturalization of streams and rivers—The special importance of integrated ecological methods in measurement of success. An example from Saxony-Anhalt (Germany). Limnologica 2004, 34, 249–263. [Google Scholar] [CrossRef]
- Unselt, C.; Haack, S.; Dreger, F.; Gebauer, D.; Brunk, I. Pflege- und Entwicklungsplan Naturpark Hoher Fläming. Landesamt Brandenburg—Landesanstalt für Großschutzgebiete (Ed.). Available online: https://www.hoher-flaeming-naturpark.de/fileadmin/user_upload/PDF/Hoher-Flaeming/PEP_Hauptstudie_Naturpark_Hoher_Flaeming_2006_bearbeitet.pdf (accessed on 25 November 2020).
- Dahm, V.; Kupilas, B.; Rolauffs, P.; Hering, D. Strategien zur Optimierung von Fließgewässer-Renaturierungsmaßnahmen und ihrer Erfolgskontrolle. Umweltbundesamt (Ed.). Texte 43/2014. Dessau-Roßlau. Available online: https://www.umweltbundesamt.de/sites/default/files/medien/378/publikationen/texte_43_2014_strategien_zur_optimierung_von_fliessgewaesser-renaturierung_0.pdf (accessed on 24 October 2020).
- Benke, A.C.; Van Arsdall, T.C.; Gillespie, D.M.; Parrish, F.K. Invertebrate Productivity in a Subtropical Blackwater River: The Importance of Habitat and Life History. Ecol. Monogr. 1984, 54, 25–63. [Google Scholar] [CrossRef]
- Speth, S.; Böttger, K. Die sustratspezifische Verteilung der Ephemeroptera, Plecoptera und Trichoptera (Insecta) in einem sandigen Bach des norddeutschen Tieflnads (Osterau, Schleswig-Holstein). Limnologica 1993, 23, 369–380. [Google Scholar]
- Hoffmann, A.; Hering, D. Wood-Associated Macroinvertebrate Fauna in Central European Streams. Int. Rev. Hydrobiol. 2000, 85, 25–48. [Google Scholar] [CrossRef]
- Pilotto, F.; Harvey, G.L.; Wharton, G.; Pusch, M.T. Simple large wood structures promote hydromorphological heterogeneity and benthic macroinvertebrate diversity in low-gradient rivers. Aquat. Sci. 2016, 78, 755–766. [Google Scholar] [CrossRef]
- Boiché, A.; Mondy, C.; Jacquemin, G.; Thiebaut, G. Découverte d’Isoptena serricornis (Pictet, 1841) en France [Plecoptera, Chloroperlidae]/First record of Isoptena serricornis (Pictet, 1841) in France [Plecoptera, Chloroperlidae]. Ephemera 2009, 10, 1–6. [Google Scholar]
- Brandt, J. Ephemera danica im Alstereinzugsgebiet. Institut für Hygiene und Umwelt—Hamburger Landesinstitut für Lebensmittelsicherheit Gesundheitsschutz und Umweltuntersuchungen. 2011. Available online: https://www.hamburg.de/contentblob/2996626/cd3f7d6818384ae51eca59f1170a7873/data/ephemera-danica.pdf (accessed on 2 November 2020).
- Putri, A.M.S.; Suryanti, S.; Widyorini, N. Hubungan Tekstur Sedimen Dengan Kandungan Bahan Organik Dan Kelimpahan Makrozoobenthos Di Muara Sungai Banjir Kanal Timur Semarang The Relation of Sediment Texture to Organic Matter and Macrozoobenthos Abundance in the Estuarine of Banjir Kanal Timur River. Saintek Perikan. Indones. J. Fish. Sci. Technol. 2017, 12, 75–80. [Google Scholar] [CrossRef]
- Noack, M.; Podraza, P.; Lechner, A. Untersuchungen von Mikrohabitaten und deren Bedeutung für das Makrozoobenthos in renaturierten Gewässerabschnitten von Sorpe und Röhr. In Korrespondenz Wasserwirtschaft; Osnabrück University: Osnabrück, Austria, 2017. [Google Scholar]
- Lester, R.E.; Wright, W.; Jones-Lennon, M.; Rayment, P. Large versus small wood in streams: The effect of wood dimension on macroinvertebrate communities. Fundam. Appl. Limnol. 2009, 174, 339–351. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Ihle | Nieplitz | Verlorenwasser |
|---|---|---|---|
| detritus (WS) | 37 (15–70) | 28 (26–43) | 34 (26–43) |
| sand (WS) | 48 (16–76) | 65 (16–76) | 60 (50–67) |
| wood (WS) | 9 (5–15) | 7 (2–15) | 7 (3–13) |
| gravel (WS) | 6 (0–14); with n = 6: 8 (6–14) | 0 | 0 |
| detritus (CS) | 58 (55–63) | 25 (23–29) | 16 (9–23) |
| sand (CS) | 42 (38–45) | 75 (71–76) | 84 (78–91) |
| Ihle | Nieplitz | Verlorenwasser | |
|---|---|---|---|
| Total number of taxa (species richness) | 85 (without G, 2.06 m2) 89 (with G, 2.44 m2) | 65 (2.06 m2) | 57 (2.06 m2) |
| number of families | 46 (without G) 50 (with G) | 37 | 35 |
| number of orders | 14 (without and with G) | 13 | 13 |
| taxa of insects | 58 (without G) 61 (with G) | 47 | 38 |
| taxa of EPT | 36 (without G) 38 (with G) | 33 | 23 |
| individual density per m2 | 6215 (with G) | 20,492 | 5457 |
| dry matter in g/m2 | 11.05 (with G) | 56.86 | 8.90 |
| Individuals | Dry Matter | ||
|---|---|---|---|
| Ihle | sand | −0.71 (p < 0.05) | −0.59 (p < 0.05) |
| detritus | 0.59 (p < 0.05) | ns | |
| Nieplitz | wood | 0.73 (p < 0.05) | 0.72 (p < 0.05) |
| Verlorenwasser | sand | −0.58 (p = 0.05) | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zigann, J.M.; Gersberg, R.M.; Lüderitz, V. Effects of Deadwood on Macroinvertebrate Assemblages in Three Sand-Type Lowland Streams. Ecologies 2023, 4, 88-105. https://doi.org/10.3390/ecologies4010008
Zigann JM, Gersberg RM, Lüderitz V. Effects of Deadwood on Macroinvertebrate Assemblages in Three Sand-Type Lowland Streams. Ecologies. 2023; 4(1):88-105. https://doi.org/10.3390/ecologies4010008
Chicago/Turabian StyleZigann, Julia Marie, Richard M. Gersberg, and Volker Lüderitz. 2023. "Effects of Deadwood on Macroinvertebrate Assemblages in Three Sand-Type Lowland Streams" Ecologies 4, no. 1: 88-105. https://doi.org/10.3390/ecologies4010008
APA StyleZigann, J. M., Gersberg, R. M., & Lüderitz, V. (2023). Effects of Deadwood on Macroinvertebrate Assemblages in Three Sand-Type Lowland Streams. Ecologies, 4(1), 88-105. https://doi.org/10.3390/ecologies4010008