
Culturable Diversity of Lichen-Associated Yeasts through Enrichment Strategies



Abstract
1. Introduction
2. Materials and Methods
2.1. Lichen Sample Collection and Molecular Identification
2.2. Liquid Enrichment Strategies
2.3. Evaluation of Terpenes as a Selective Liquid Enrichment Strategy
2.4. Evaluation of Nikkomycin Z as a Selective Inhibitor with Liquid Enrichment Strategy
2.5. Direct Plating Methods
2.6. Fungal Isolate Identification, Phylogenetic Placement Analyse, Visualization, and Storage
3. Results
3.1. Taxonomy of Lichen-Associated Yeasts from Liquid Enrichment Strategies
3.2. Evaluation of Liquid Enrichment Strategies for the Isolation of Lichen-Associated Yeasts
3.3. Evaluation of Terpenes and Nikkomycin Z for the Isolation of Lichen-Associated Yeasts
4. Discussion
4.1. Lichen-Associated Ascomycetous Yeast
4.2. Lichen-Associated Basidiomycetous Yeast
4.3. Methodological Observations and Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, J.M.; et al. Life with 6000 genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K.; Johri, B.N. Basidiomycetous yeasts: Current status. In Yeast Biotechnology: Diversity and Applications; Satyanarayana, T., Kunze, G., Eds.; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar] [CrossRef]
- Stefanini, I. Yeast-insect associations: It takes guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Petzold, D.E.; Mulhern, T. Vegetation distributions along lichen-dominated slopes of opposing aspect in the Eastern Canadian subarctic. Arctic 1987, 40, 221–224. [Google Scholar] [CrossRef]
- Millanes, A.M.; Diederich, P.; Wedin, M. Cyphobasidium gen. nov., a new lichen inhabiting lineage in the Cystobasidiomycetes (Pucciniomycotina, Basidiomycota, Fungi). Fungal Biol. 2016, 120, 1468–1477. [Google Scholar] [CrossRef]
- Spribille, T.; Tuovinen, V.; Res, P.; Vanderpool, D.; Wolinski, H.; Aime, M.C.; Schneider, K.; Stabentheiner, E.; Toome, H.M.; Thor, G.; et al. Basidiomycete yeasts in the cortex of ascomycete mac-rolichens. Science 2016, 353, 488–492. [Google Scholar] [CrossRef]
- Millanes, A.; Diederich, P.; Ekman, S.; Wedin, M. Phylogeny and character evolution in jelly fungi (Tremellomycetes, Basid-iomycota, Fungi). Mol. Phylogenet. Evol. 2011, 61, 12–28. [Google Scholar] [CrossRef]
- Spribille, T. Relative symbiont input and the lichen symbiotic outcome. Curr. Opin. Plant Biol. 2018, 44, 57–63. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Fell, J.W.; Teun, B.T.; Vincent, R.V. Chapter 7 Methods for isolation, phenotypic characterization and maintenance of yeasts. In The Yeasts, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: New York, NY, USA, 2011; pp. 87–110. [Google Scholar]
- Gasulla, F.; Del Campo, E.M.; Casano, L.M.; Guéra, A. Advances in understanding of desiccation tolerance of lichens and li-chen-forming algae. Plants 2021, 10, 807. [Google Scholar] [CrossRef]
- Takahagi, T.; Endo, T.; Yamamoto, Y.; Sato, F. Lichen photobionts show tolerance against lichen acids produced by lichen mycobionts. Biosci. Biotechnol. Biochem. 2008, 72, 3122–3127. [Google Scholar] [CrossRef]
- Culberson, C.F. Chemical and Botanical Guide to Lichen Products; University of North Carolina Press: Chapel Hill, CA, USA, 1969; pp. 58–62. [Google Scholar]
- Kahriman, N.; Yazici, K.; Arslan, T.; Aslan, A.; Karaoglu, S.A.; Yayli, N. Chemical composition and antimicrobial activity of the essential oils from Evernia prunastri (L.) ach. and Evernia divaricata (L.) ach. Asian J. Chem. 2011, 23, 1937–1939. [Google Scholar]
- Thanh, V.N.; Smit, M.S.; Moleleki, N.; Fell, J.W. Rhodotorula cycloclastica sp. nov., Rodotorula retinophila sp. nov., and Rodotorula terpenoidalis sp. nov., three limonene-utilizing yeasts isolated from soil. FEMS Yeast Res. 2004, 4, 857–863. [Google Scholar]
- Chaffin, W.L.; Lopez-Ribot, J.L.; Casanova, M.; Gozalbo, D.; Martinez, J.P. Cell wall and secreted proteins of Candida albicans: Identification, function, and expression. Microbiol. Mol. Biol. Rev. 1998, 62, 130–180. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J. Fungal Cell Wall: Structure, Synthesis and Assembly; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Scorzetti, G.; Fell, J.W.; Fonseca, A.; Statzell-Tallman, A. Systematics of basidiomycetous yeasts: A comparison of large subunit D1/D2 and internal transcribed spacer rDNA regions. FEMS Yeast Res. 2002, 2, 495–517. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; Szöke, S.; Cardinali, G.; Eberhardt, U.; Stielow, B.; de Vries, M.; Verkleij, G.J.M.; Crous, P.W.; Boekhout, T.; et al. DN barcoding analysis of more than 9000 yeasts isolates contributes to quantitative thresholds for yeast species and genera delimitation. Stud. Mycol. 2016, 85, 91–105. [Google Scholar] [CrossRef]
- Osmundson, T.W.; Eyre, C.A.; Hayden, K.M.; Dhillon, J.; Garbelotto, M.M. Back to basics: An evaluation of NaOH and alternative rapid DNA extraction protocols for DNA barcoding, genotyping, and disease diagnostics from fungal and oomycete samples. Mol. Ecol. Resour. 2013, 13, 66–74. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of my-corrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, N., Gelfand, D., Sninsky, J., White, T., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Rehner, S.A.; Samuels, G.J. Molecular systematics of the Hypocreales: A teleomorph gene phylogeny and the status of their anamorphs. Can. J. Bot. 1995, 73, S816–S823. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Demarne, F.-E. ‘Rose-scented geranium’ a Pelargonium grown for the perfume industry. In Geranium and Pelargonium: The Genera Geranium and Pelargonium; Lis-Balchin, M., Ed.; Taylor & Francis: New York, NY, USA, 2002; pp. 193–211. [Google Scholar]
- Tariq, V.; Devlin, P. Sensitivity of fungi to Nikkomycin Z. Fungal Genet. Biol. 1996, 20, 4–11. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The barcode of life data system (www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Gouy, M.; Tannier, E.; Comte, N.; Parsons, D.P. Seaview version 5: A multiplatform software for multiple sequence alignment, molecular phylogenetic analyses, and tree reconciliation. In Multiple Sequence Alignment; Methods in Molecular Biology; Katoh, K., Ed.; Humana: New York, NY, USA, 2021; pp. 241–260. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Zhang, H.; Gao, S.; Lercher, M.J.; Chen, W.-H.; Hu, S. Evolview v2: An online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 2016, 44, W236–W241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wei, X.L.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and distribution of lichen-associated fungi in the Ny-Ålesund Region (Svalbard, High Arctic) as revealed by 454 pyrosequencing. Sci. Rep. 2015, 5, 14850. [Google Scholar] [CrossRef] [PubMed]
- Banchi, E.; Stankovic, D.; Fernández-Mendoza, F.; Gionechetti, F.; Pallavicini, A.; Muggia, L. ITS2 metabarcoding analysis complements lichen mycobiome diversity data. Mycol. Prog. 2018, 17, 1049–1066. [Google Scholar] [CrossRef]
- Aime, M.C.; Miller, A.; Aoki, T.; Bensch, K.; Cai, L.; Crous, P.W.; Hawksworth, D.L.; Hyde, K.; Kirk, P.; Lücking, R.; et al. How to publish a new fungal species, or name, version 3.0. IMA Fungus 2021, 12, 11. [Google Scholar] [CrossRef]
- Biosca, E.G.; Flores, R.; Santander, R.D.; Díez-Gil, J.L.; Barreno, E. Innovative approaches using lichen enriched media to improve isolation and capturability of lichen associated bacteria. PLoS ONE 2016, 11, e0160328. [Google Scholar] [CrossRef]
- Muggia, L.; Grube, M. Fungal diversity in lichens: From extremotolerance to interactions with algae. Life 2018, 8, 15. [Google Scholar] [CrossRef]
- Fernámdez-Mendoza, F.; Fleischhacker, A.; Kopun, T.; Grube, M.; Muggia, L. ITS1 metabarcoding highlights low specificity of lichen mycobiomes at a local scale. Mol. Ecol. 2017, 26, 4811–4830. [Google Scholar] [CrossRef] [PubMed]
- Romão, D.; Staley, C.; Ferreira, F.; Rodrigues, R.; Sabino, R.; Veríssimo, C.; Wang, P.; Sadowsky, M.; Brandão, J. Next-generation sequencing and culture-based techniques offer complementary insights into fungi and prokaryotes in beach sands. Mar. Pollut. Bull. 2017, 119, 351–358. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S.; Govindarajulu, M.B.; Rajamani, T.; Tripathi, M.; Joshi, Y. Endolichenic fungi in lichens of Champawat district, Uttarakhand, northern India. Mycol. Progress 2017, 16, 205–211. [Google Scholar] [CrossRef]
- Selbmann, L.; Grube, M.; Onofri, S.; Isola, D.; Zucconi, L. Antarctic epilithic lichens as niches for black meristematic fungi. Biology 2013, 2, 784–797. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S.; Zalar, P.; Urzì, C.; de Leo, F.; Yurlova, N.A.; Sterflinger, K. Relationships of dothideaceous black yeasts and meristematic fungi based on 5.8S and ITS2 rDNA sequence comparison. Stud. Mycol. 1999, 43, 31–37. [Google Scholar]
- Kachalkin, A.V.; Glushakova, A.M.; Pankratov, T.A. Yeast population of the Kindo Peninsula lichens. Microbiology 2017, 86, 786–792. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z. The genera of fungi—G 4: Camarosporium and Dothiora. IMA Fungus 2017, 8, 131–152. [Google Scholar] [CrossRef] [PubMed]
- Clauzade, G.; Diederich, P.P.; Roux, C. Nelikenigintaj fungoj likenlogaj–Ilustrita determinlibro. Bull. Soc. Linn. Provence 1989, 1, 1–142. [Google Scholar]
- Diederish, P.; Lawrey, J.D.; Damien, E. The 2018 classification and checklist of lichenicolous fungi, with 2000 non-lichenized, obligately lichenicolous taxa. Bryologist 2018, 121, 340–425. [Google Scholar] [CrossRef]
- Ekanayaka, A.H.; Hyde, K.D.; Gentekaki, E.; McKenzie, E.H.C.; Zhao, Q.; Bulgakov, T.S.; Camporesi, E. Preliminary classification of Leotiomycetes. Mycosphere 2019, 10, 310–489. [Google Scholar] [CrossRef]
- Frank, J.L. Mycosymbioces mycenophila. Index Fungorum. 2014. Available online: https://www.gbif.org/species/9746710 (accessed on 1 November 2022).
- Edwards, A.; Leech, T.; Senior, I. A gall-inducing infection of Lepista spp. In Norfolk by Mycosymbioces mycenophila—Fist record for Britain. Field Mycol. 2020, 21, 119–123. [Google Scholar] [CrossRef]
- Tanney, J.B.; Quijada, L. Comments on the occurrence of yeast-like morphologies in the Leotiomycetes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005141. [Google Scholar] [CrossRef]
- Černajová, I.; Škaloud, P. The first survey of Cystobasidiomycetes yeasts in the lichen genus Cladonia; with the description of Lichenozyma pisutiana gen. nov., sp. nov. Fungal Biol. 2019, 123, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Cometto, A.; Leavitt, S.D.; Millanes, A.M.; Wedin, M.; Grube, M.; Muggia, L. The yeast lichenosphere: High diversity of ba-sidiomycetes from the lichens Tephromela atra and Rhizoplaca melanophthalma. Fungal Biol. 2022, 126, 587–608. [Google Scholar] [CrossRef] [PubMed]
- Aime, M.C.; Toome, M.; McLaughlin, D.J. 10 Pucciniomycotina. In Systematics and Evolution; McLaughlin, D., Spatafora, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 7A. [Google Scholar] [CrossRef]
- Petch, T. Note on the biology of the genus Septobasidium. Ann. Bot. 1911, 25, 842. [Google Scholar] [CrossRef]
- Oberwinkler, F. Yeasts in Pucciniomycotina. Myco. Prog. 2017, 16, 831–856. [Google Scholar] [CrossRef]
- Bai, L.; Chong, A.; Zhu, Y.; Wang, X.; Wang, M.; Li, J.; Zhang, Z.; Zhao, X. First isolation and identification of cold adaptive yeast Curvibasidium rogersii from Usnea lichen and genome-based studies of its biological properties. Acta Microbiol. Sin. 2022, 62, 567–578. [Google Scholar]
- Wang, Q.; Li, J.; Yang, J.; Zou, Y.; Zhao, X.-Q. Diversity of endophytic bacterial and fungal microbiota associated with the medicinal lichen Usnea longissimi at high altitudes. Front. Microbiol. 2022, 13, 958917. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, Y.; Wang, X.; Wei, X.; Wei, J. Lichen-associated fungal community in Hypogymnia hypotrypa (Parmeliaceae, Ascomycota) affected by geographic distribution and altitude. Front. Microbiol. 2016, 7, 1231. [Google Scholar] [CrossRef]
- Dimitrova, S.; Pavlova, K.; Lukanov, L.; Korotkova, E.; Petrova, E.; Zagorchev, P.; Kuncheva, M. Production of metabolites with antioxidant and emulsifying properties by Antarctic strain Sporobolomyces salmonicolor AL1. Appl. Biochem. Biotechnol. 2013, 169, 301–311. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; Dayo-Owoyemi, I.; Nobre, F.S.; Pagnocca, F.C.; Chaud, L.C.S.; Pessoa, A.; Felipe, M.G.A.; Sette, L.D. Taxonomic assessment and enzymes production by yeasts isolated from marine and terrestrial Antarctic samples. Extremophiles 2013, 17, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Middelhoven, W.J.; Koorevaar, M.; Schuur, G.W. Degradation of benzene compounds by yeasts in acidic soils. Plant Soil 1992, 145, 37–43. [Google Scholar] [CrossRef]
- Isola, D.; Scano, A.; Orrù, G.; Prenafeta-Boldú, F.X. Hydrocarbon-contaminated sites; is there something more than Exophiala xenobiotica? New insights into black fungal diversity using the long cold incubation method. J. Fungi 2021, 7, 817. [Google Scholar] [CrossRef] [PubMed]
- Deacon, J. Fungal Biology; Blackwell: Malden, MA, USA, 2006; p. 371. [Google Scholar]
- Muggia, L.; Fleischhacker, A.; Kopun, T.; Grube, M. Extremotolerant fungi from alpine rock lichens and their phylogenetic relationships. Fungal Divers. 2016, 76, 119–142. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
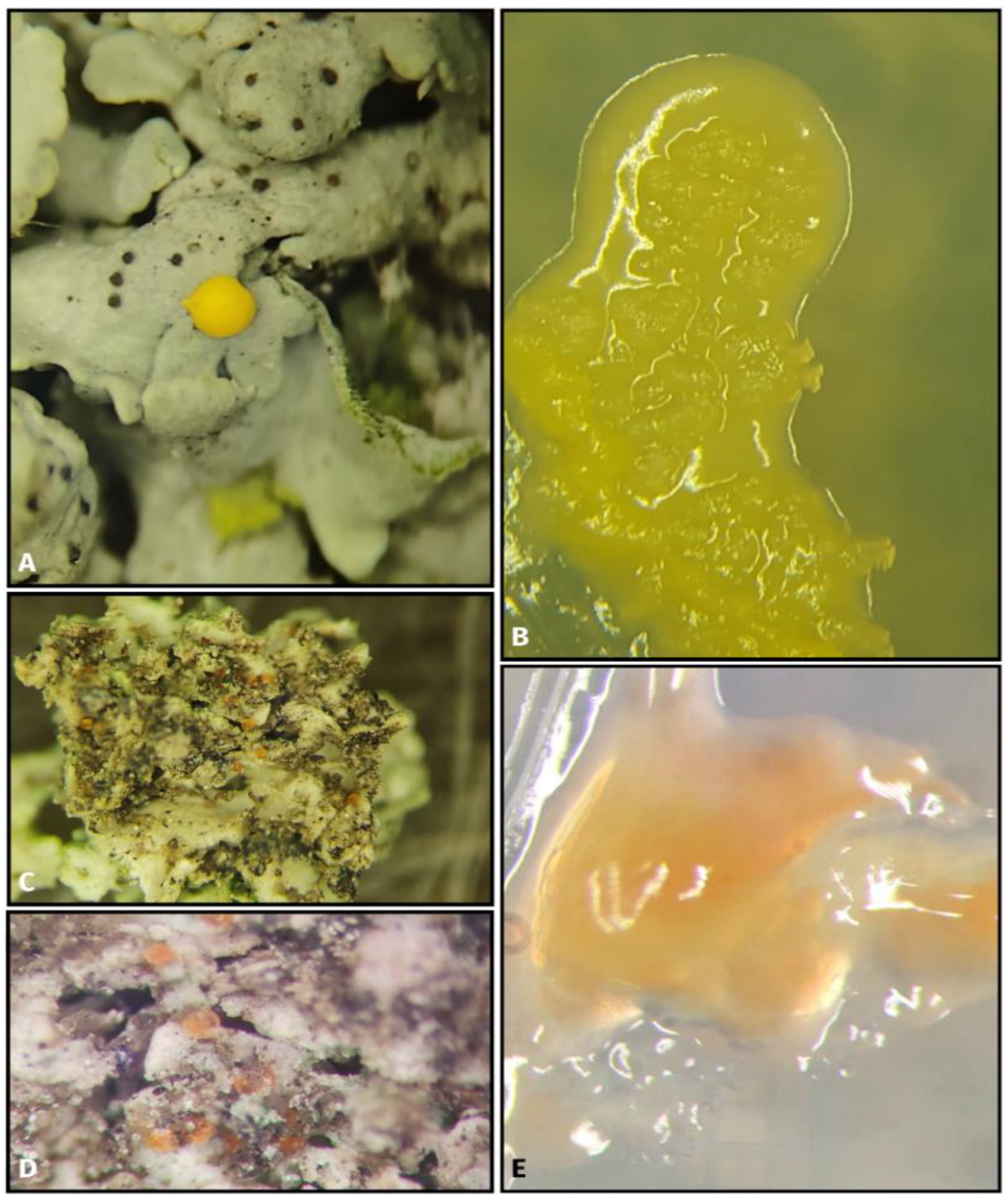{kind=link}
{kind=link}
{kind=link}
{kind=link}
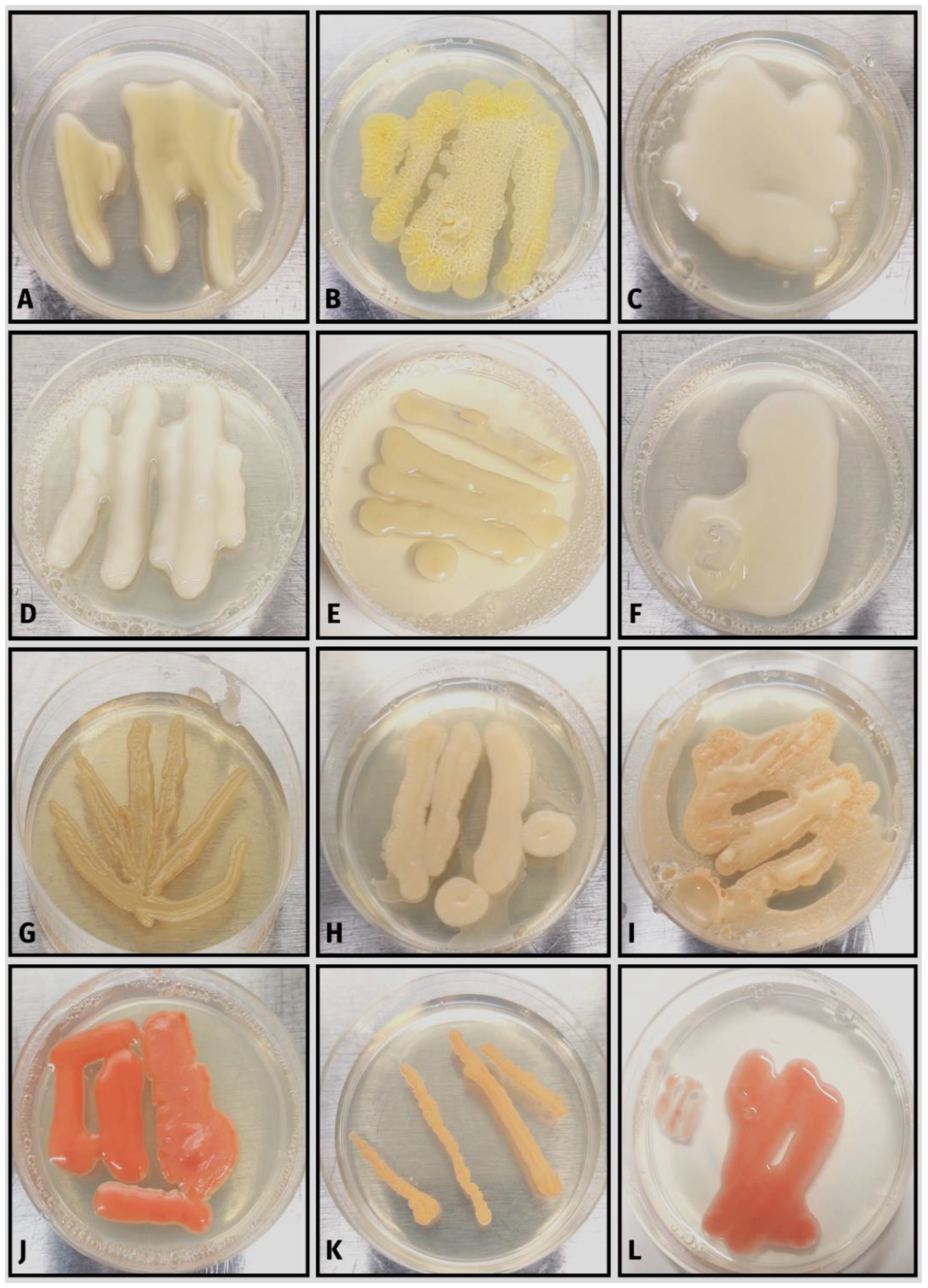{kind=link}
| Lichen Identification | Isolation Technique 1 | Substrate ID | Location 2 | Substrate Type | PUL Acc. Number | NCBI Match/Morphology 3 |
|---|---|---|---|---|---|---|
| Aspicilia sp. | LES/terpene | DBRRPAJu05 | PA | Rock | F29321 | HM060755 |
| Buellia sp. | LES | DBRBNC22-C | NC | Branch | F29267 | morphology |
| LES | DBRBNC22-E | NC | Branch | F29269 | KX512852 | |
| Candelaria concolor | LES | DBRBY01 | IN | Branch | F29314 | morphology |
| NZ | DBRNZ-1 | IN | Branch | F29327 | DQ986791 | |
| NZ | DBRNZ-7 | IN | Branch | F29330 | DQ986791 | |
| Canoparmelia caroliniana | LES | DBRBNC05-C | NC | Branch | F29240 | morphology |
| LES | DBRBNC08 | NC | Branch | F29243 | morphology | |
| LES | DBRBNC09 | NC | Branch | F29244 | AY584634 | |
| LES | DBRDNC10-B | NC | Bark | F29246 | AY584634 | |
| LES | DBRBNC31 | NC | Branch | F29279 | AY584634 | |
| Cladonia sp. | LES | DBRGC01 | PA | Ground | F29312 | DQ534464 |
| LES | DBRGRD01 | PA | Ground | F29313 | MT553308 | |
| Lecanora sp. | LES | DBRBNC05-B | NC | Branch | F29239 | morphology |
| LES | DBRBNC11-A | NC | Branch | F29247 | morphology | |
| LES | DBRBNC13-A | NC | Branch | F29250 | KJ766583 | |
| LES | DBRBNC16-B | NC | Branch | F29256 | OL663912 | |
| LES | DBRBNC20-A | NC | Branch | F29262 | OL663912 | |
| LES | DBRBNC20-B | NC | Branch | F29263 | OL663870 | |
| Lepraria harrisiana | LES | DBRDNC06 | NC | Bark | F29241 | JQ070289 |
| Lepraria sp. | LES | DBRDNC07 | NC | Bark | F29242 | morphology |
| LES | DBRDNC12 | NC | Bark | F29249 | morphology | |
| LES | DBRDNC29 | NC | Bark | F29277 | morphology | |
| Parmotrema sp. | LES | DBRBNC03 | NC | Branch | F29236 | DQ912339 |
| LES | DBRDNC04 | NC | Bark | F29237 | morphology | |
| LES | DBRBNC05-A | NC | Branch | F29238 | morphology | |
| LES | DBRBNC11-B | NC | Branch | F29248 | MG233922 | |
| LES | DBRBNC14 | NC | Branch | F29253 | morphology | |
| LES | DBRBNC16-A | NC | Branch | F29255 | morphology | |
| LES | DBRBNC17 | NC | Branch | F29257 | morphology | |
| LES | DBRBNC18-B | NC | Branch | F29259 | morphology | |
| LES | DBRBNC21 | NC | Branch | F29264 | morphology | |
| LES | DBRBNC22-D | NC | Branch | F29268 | morphology | |
| Physcia sp. | LES | DBRBIN01-1 | IN | Branch | F29220 | morphology |
| LES | DBRBIN01-2 | IN | Branch | F29221 | morphology | |
| LES | DBRBIN02-1 | IN | Branch | F29222 | morphology | |
| LES | DBRBIN02-2 | IN | Branch | F29223 | morphology | |
| LES | DBRBIN03-1 | IN | Branch | F29224 | morphology | |
| LES | DBRBNC13-B | NC | Branch | F29251 | AY860575 | |
| LES | DBRBNC13-C | NC | Branch | F29252 | JQ301596 | |
| NZ | DBRNZ-2 | IN | Branch | F29328 | JQ301596 | |
| NZ | DBRNZ-4 | IN | Branch | F29329 | JQ301596 | |
| NZ | DBRNZ-8 | IN | Branch | F29331 | JQ301596 | |
| NZ | DBRNZ-9 | IN | Branch | F29332 | JQ301596 | |
| NZ | DBRNZ-10 | IN | Branch | F29333 | JQ301596 | |
| Porpidia crustulata | LES/terpene | DBRRPAJu01 | PA | Rock | F29317 | AY532943 |
| LES/terpene | DBRRPAJu02 | PA | Rock | F29318 | AY532943 | |
| Porpidia sp. | LES | DBRRPAAp01 | PA | Rock | F29229 | AY532967 |
| LES | DBRRPAAp02 | PA | Rock | F29230 | AY532973 | |
| LES/terpene | DBRRPAJu03 | PA | Rock | F29319 | morphology | |
| LES/terpene | DBRRPAJu06 | PA | Rock | F29322 | AY532973 | |
| LES/terpene | DBRRPAJu09 | PA | Rock | F29325 | AY532943 | |
| Punctelia sp. | LES | DBRBNC02 | NC | Branch | F29235 | AY584636 |
| Punctelia ulophylla | LES | DBRDNC10-A | NC | Bark | F29245 | GU994641 |
| Rhizocarpon sp. | LES/terpene | DBRRPAJu04 | PA | Rock | F29320 | KJ766652 |
| LES/terpene | DBRRPAJu07 | PA | Rock | F29323 | KJ766652 | |
| LES/terpene | DBRRPAJu08 | PA | Rock | F29324 | KJ766652 | |
| Rhizoplaca sp. | LES/terpene | DBRRPAJu10 | PA | Rock | F29326 | MN756833 |
| Trapelia sp. | LES | DBRRPAAp04 | PA | Rock | F29232 | KJ766667 |
| Umbilicaria sp. | LES | DBRPNC27 | NC | Pinecone | F29274 | HM161589 |
| Usnea sp. | LES | DBRBNC18-A | NC | Branch | F29258 | MH310889 |
| LES | DBRBNC19-A | NC | Branch | F29260 | morphology | |
| LES | DBRBNC19-B | NC | Branch | F29261 | morphology | |
| LES | DBRBNC22-B | NC | Branch | F29266 | morphology | |
| LES | DBRBNC22-F | NC | Branch | F29270 | morphology | |
| LES | DBRBNC23 | NC | Branch | F29271 | morphology | |
| LES | DBRBNC24-A | NC | Branch | F29272 | KY033353 | |
| LES | DBRBNC24-B | NC | Branch | F29273 | morphology | |
| LES | DBRBNC28-A | NC | Branch | F29275 | morphology | |
| LES | DBRBNC28-B | NC | Branch | F29276 | morphology | |
| LES | DBRBNC32 | NC | Branch | F29280 | morphology | |
| Verrucaria dolosa | LES | DBRRIN03-1 | IN | Rock | F29228 | JX848554 |
| Verrucaria sp. | LES | DBRRIN01-1 | IN | Rock | F29225 | FJ664859 |
| LES | DBRRIN01-2 | IN | Rock | F29226 | morphology | |
| Xanthoparmelia sp. | LES | DBRBG01 | PA | Rock | F29315 | morphology |
| LES | DBRBG02 | IN | Rock | F29316 | morphology | |
| LES | DBRRPAAp03 | PA | Rock | F29231 | KJ766676 |
| Phylum | Class | Order | Family | Genus |
|---|---|---|---|---|
| LES * | ||||
| Ascomycota | Dothideomycetes | Dothideales | Dothioraceae | Dothiora |
| Plowrightia | ||||
| Scleroconidioma | ||||
| Aureobasidium | ||||
| Eurotiomycetes | Chaetothyriales | Herpotrichiellaceae | Exophiala | |
| Phialophora | ||||
| Leotiomycetes | Helotiales | Calloriaceae | Tricellula | |
| Leotiales | Unknown | |||
| Leotiomycetes incertae sedis | Unknown | |||
| Sordariomycetes | Chaiochaetales | Coniochaetaceae | Coniochaeta | |
| Glomerellales | Plectosphaerellaceae | Gibellulopsis | ||
| Plectosphaerella | ||||
| Hypocreales | Clavicipitaceae | Metapochonia | ||
| Nectriaceae | Thyronectria | |||
| Sordariales | Cephalothecaceae | Phialemonium | ||
| Saccharomycetes | Saccharomycetales | Debaryomycetaceae | Debaryomyces | |
| Saccharomycetales | Saccharomycetaceae | Lachancea | ||
| Sordariomycetes incertae sedis | Thyridiaceae | Thyridium | ||
| Basidiomycota | Tremellomycetes | Cystofilobasidiales | Cystofilobasidiaceae | Cystofilobasidium |
| Filobasidiales | Filobasidiaceae | Heterocephalacria | ||
| Tremellales | Bulleraceae | Bullera | ||
| Genolevuria | ||||
| Pseudotremella | ||||
| Bulleribasidiaceae | Vishniacozyma | |||
| Carcinomycetaceae | Carcinomyces | |||
| Cryptococcaceae | Kwoniella | |||
| Rhynchogastremataceae | Papiliotrema | |||
| Sirobasidiaceae | Fibulobasidium | |||
| Tremellaceae | Tremella | |||
| Trimorphomyceteceae | Carlosrosaea | |||
| Saitozyma | ||||
| Sugitazyma | ||||
| Cystobasidiomycetes | Unknown | |||
| Microbotryomycetes | Kriegeriales | Kriegeriaceae | Libkindia | |
| Leucosporidiales | Leucosporidiaceae | Leucosporidium | ||
| Microbotryomycetes incertae sedis | Chrysozymaceae | Oberwinklerozyma | ||
| Trigonosporomyces | ||||
| Yunzhangia | ||||
| Colacogloea | ||||
| Curvibasidium | ||||
| Sporidiobolales | Sporidiobolaceae | Rhodotorula | ||
| Sporobolomyces | ||||
| Pucciniomycetes | Septobasidiales | Septobasidiaceae | Septobasidium | |
| Direct Plating | ||||
| Ascomycota | Dothideomycetes | Dothideales | Saccotheciaceae | Aureobasidium |
| Saccharomycetes | Saccharomycetales | Phaffomycetaceae | Barnettozyma | |
| Taphrinomycetes | Taphrinales | Taphrinaceae | Taphrina | |
| Basidiomycota | Tremellomycetes | Filobasidiales | Filobasidiaceae | Filobasidium |
| Tremellales | Bulleribasidiaceae | Diozegia | ||
| Hannaella | ||||
| Rhynchogastremataceae | Papiliotrema | |||
| Tremellaceae | Tremella | |||
| Cystobasidiomycetes | Cystobasidiomycetes incertae sedis | Symmetrosporaceae | Symmetrospora | |
| Erythrobasidiales | Erythrobasidiaceae | Erythrobasidium | ||
| Exobasidiomycetes | Microstromatales | Microstromatales incertae sedis | Jaminaea | |
| Malasseziomycetes | Malasseziales | Malesseziaceae | Malassezia | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raudabaugh, D.B.; Aime, M.C. Culturable Diversity of Lichen-Associated Yeasts through Enrichment Strategies. Ecologies 2023, 4, 152-170. https://doi.org/10.3390/ecologies4010012
Raudabaugh DB, Aime MC. Culturable Diversity of Lichen-Associated Yeasts through Enrichment Strategies. Ecologies. 2023; 4(1):152-170. https://doi.org/10.3390/ecologies4010012
Chicago/Turabian StyleRaudabaugh, Daniel B., and M. Catherine Aime. 2023. "Culturable Diversity of Lichen-Associated Yeasts through Enrichment Strategies" Ecologies 4, no. 1: 152-170. https://doi.org/10.3390/ecologies4010012
APA StyleRaudabaugh, D. B., & Aime, M. C. (2023). Culturable Diversity of Lichen-Associated Yeasts through Enrichment Strategies. Ecologies, 4(1), 152-170. https://doi.org/10.3390/ecologies4010012