
Bioluminescence Resonance Energy Transfer (BRET) Allows Monitoring the Barnase-Barstar Complex In Vivo


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Strains and Culture Conditions
2.3. Genetic Manipulations
2.4. Construction of Plasmids for Expression in E. coli Cells
2.5. Expression of the Genetic Constructs in E. coli
2.6. Registration of Fluorescence and Bioluminescence Spectra
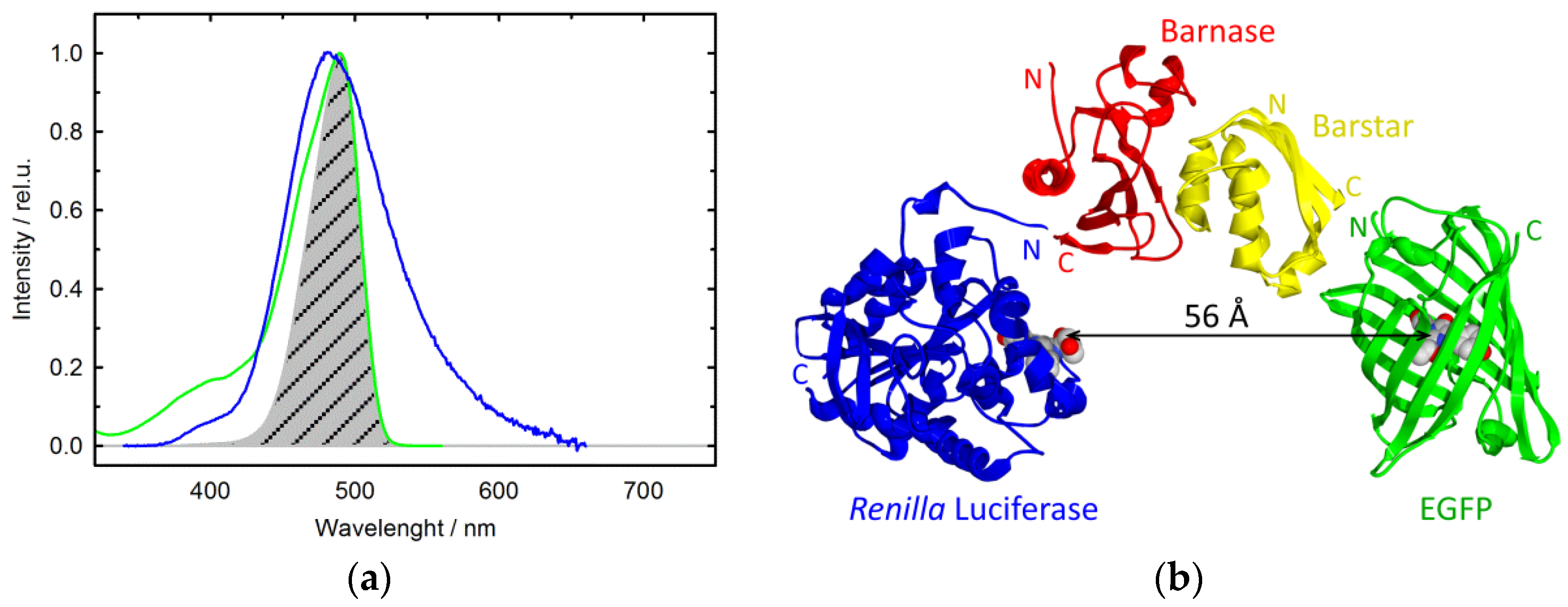
3. Results
Generation and Identification of Fusion Proteins Barnase–RLuc and Barstar–EGFP
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haddock, S.H.D.; Moline, M.A.; Case, J.F. Bioluminescence in the Sea. Annu. Rev. Mar. Sci. 2010, 2, 443–493. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, O. Bioluminescence in the sea: Photoprotein systems. Symp. Soc. Exp. Biol. 1985, 39, 352–371. [Google Scholar]
- Ward, W.; Cormier, M. An energy transfer protein in coelenterate bioluminescence. Characterization of the Renilla green-fluorescent protein. J. Biol. Chem. 1979, 254, 781–788. [Google Scholar] [CrossRef]
- Gorokhovatsky, A.Y.; Marchenkov, V.V.; Rudenko, N.V.; Ivashina, T.V.; Ksenzenko, V.N.; Burkhardt, N.; Semisotnov, G.V.; Vinokurov, L.M.; Alakhov, Y.B. Fusion of Aequorea victoria GFP and aequorin provides their Ca2+-induced interaction that results in red shift of GFP absorption and efficient bioluminescence energy transfer. Biochem. Biophys. Res. Commun. 2004, 320, 703–711. [Google Scholar] [CrossRef]
- Wu, P.; Brand, L. Resonance Energy Transfer: Methods and Applications. Anal. Biochem. 1994, 218, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Morise, H.; Shimomura, O.; Johnson, F.H.; Winant, J. Intermolecular energy transfer in the bioluminescent system of Aequorea. Biochemistry 1974, 13, 2656–2662. [Google Scholar] [CrossRef]
- De, A.; Jasani, A.; Arora, R.; Gambhir, S.S. Evolution of BRET Biosensors from Live Cell to Tissue-Scale In vivo Imaging. Front. Endocrinol. 2013, 4, 131. [Google Scholar] [CrossRef] [Green Version]
- Dale, N.C.; Johnstone, E.K.M.; White, C.W.; Pfleger, K.D.G. NanoBRET: The Bright Future of Proximity-Based Assays. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef]
- Yeh, H.W.; Ai, H.-W. Development and Applications of Bioluminescent and Chemiluminescent Reporters and Biosensors. Annu. Rev. Anal. Chem. 2019, 12, 129–150. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, O.; Johnson, F.H. Chemical nature of the light emitter in bioluminescence of aequorin. Tetrahedron Lett. 1973, 14, 2963–2966. [Google Scholar] [CrossRef]
- Xu, Y.; Piston, D.W.; Johnson, C.H. A bioluminescence resonance energy transfer (BRET) system: Application to interacting circadian clock proteins. Proc. Natl. Acad. Sci. 1999, 96, 151–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audet, M.; Lagacé, M.; Silversides, D.W.; Bouvier, M. Protein-protein interactions monitored in cells from transgenic mice using bioluminescence resonance energy transfer. FASEB J. 2010, 24, 2829–2838. [Google Scholar] [CrossRef] [PubMed]
- Buckle, A.; Schreiber, G.; Fersht, A.R. Protein-protein recognition: Crystal structural analysis of a barnase-barstar complex at 2.0-.ANG. resolution. Biochemistry 1994, 33, 8878–8889. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, G.; Fersht, A.R. Interaction of barnase with its polypeptide inhibitor barstar studied by protein engineering. Biochemistry 1993, 32, 5145–5150. [Google Scholar] [CrossRef]
- Shilova, O.; Kotelnikova, P.; Proshkina, G.; Shramova, E.; Deyev, S. Barnase-Barstar Pair: Contemporary Application in Cancer Research and Nanotechnology. Molecules 2021, 26, 6785. [Google Scholar] [CrossRef]
- Brown, N.E.; Blumer, J.B.; Hepler, J.R. Bioluminescence resonance energy transfer to detect protein-protein interactions in live cells. In Protein-Protein Interactions; Humana Press: New York, NY, USA, 2015; pp. 457–465. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.W.; Samanta, A.; Díaz, S.A.; Buckhout-White, S.; Walper, S.A.; Goldman, E.R.; Medintz, I.L. Dendrimeric DNA Nanostructures as Scaffolds for Efficient Bidirectional BRET–FRET Cascades. Adv. Opt. Mater. 2017, 5, 1700181. [Google Scholar] [CrossRef]
- Loening, A.M.; Fenn, T.D.; Wu, A.M.; Gambhir, S.S. Consensus guided mutagenesis of Renilla luciferase yields enhanced stability and light output. Protein Eng. Des. Sel. 2006, 19, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Issad, T.; Boute, N.; Boubekeur, S.; Lacasa, D. Interaction of PTPB with the insulin receptor precursor during its biosynthesis in the endoplasmic reticulum. Biochimie 2005, 87, 111–116. [Google Scholar] [CrossRef]
- Kamal, M.; Marquez, M.; Vauthier, V.; Leloire, A.; Froguel, P.; Jockers, R.; Couturier, C. Improved donor/acceptor BRET couples for monitoring β-arrestin recruitment to G protein-coupled receptors. Biotechnol. J. 2009, 4, 1337–1344. [Google Scholar] [CrossRef] [Green Version]
- Gomes, I.; Filipovska, J.; Jordan, B.A.; Devi, L.A. Oligomerization of opioid receptors. Methods 2002, 27, 358–365. [Google Scholar] [CrossRef]
- Gorokhovatsky, A.Y.; Rudenko, N.V.; Marchenkov, V.V.; Skosyrev, V.S.; A Arzhanov, M.; Burkhardt, N.; Zakharov, M.V.; Semisotnov, G.; Vinokurov, L.M.; Alakhov, Y.B. Homogeneous assay for biotin based on Aequorea victoria bioluminescence resonance energy transfer system. Anal. Biochem. 2003, 313, 68–75. [Google Scholar] [CrossRef]
- Hartley, R.W. Barnase and barstar: Expression of its cloned inhibitor permits expression of a cloned ribonuclease. J. Mol. Biol. 1988, 202, 913–915. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.J.; Russell, D.D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; ISBN 0-87969-577-3. [Google Scholar]
- Kholod, N.; Mustelin, T. Novel Vectors for Co-Expression of Two Proteins in E. coli. BioTechniques 2001, 31, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer: Boston, MA, USA, 2006. [Google Scholar]
- Patterson, G.; Knobel, S.; Sharif, W.; Kain, S.; Piston, D. Use of the green fluorescent protein and its mutants in quantitative fluorescence microscopy. Biophys. J. 1997, 73, 2782–2790. [Google Scholar] [CrossRef] [Green Version]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Plasmids | Expression Product | Cell Optical Density After Growth (A590) |
|---|---|---|
| p11cjoe/barstar-EGFP | barstar–EGFP fusion | 2.7 |
| p11cjoe/RLuc | Renilla luciferase | 3.0 |
| p28/barnase-RLuc | barnase–luciferase fusion | 1.1 |
| p28/barnase-RLuc + p11cjoe/barstar-EGFP (co-expression) | barnase–luciferase and barstar–EGFP fusions | 4.5 |
| Parameter | Name | Value |
|---|---|---|
| J(λ) | the overlapping integral (cm−1mol−1nm4) | 1.329 × 1015 |
| R0 | Forster radius (Å) | 32.2 |
| k2 | chromophore orientation | 0.67 |
| n | refraction factor | 1.4 |
| QD | quantum yield of donor | 0.053 |
| QA | quantum yield of acceptor | 0.6 |
| E | BRET efficiency | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchenkov, V.; Ivashina, T.; Marchenko, N.; Ksenzenko, V.; Semisotnov, G. Bioluminescence Resonance Energy Transfer (BRET) Allows Monitoring the Barnase-Barstar Complex In Vivo. Biophysica 2022, 2, 72-78. https://doi.org/10.3390/biophysica2010007
Marchenkov V, Ivashina T, Marchenko N, Ksenzenko V, Semisotnov G. Bioluminescence Resonance Energy Transfer (BRET) Allows Monitoring the Barnase-Barstar Complex In Vivo. Biophysica. 2022; 2(1):72-78. https://doi.org/10.3390/biophysica2010007
Chicago/Turabian StyleMarchenkov, Victor, Tanya Ivashina, Natalia Marchenko, Vladimir Ksenzenko, and Gennady Semisotnov. 2022. "Bioluminescence Resonance Energy Transfer (BRET) Allows Monitoring the Barnase-Barstar Complex In Vivo" Biophysica 2, no. 1: 72-78. https://doi.org/10.3390/biophysica2010007
APA StyleMarchenkov, V., Ivashina, T., Marchenko, N., Ksenzenko, V., & Semisotnov, G. (2022). Bioluminescence Resonance Energy Transfer (BRET) Allows Monitoring the Barnase-Barstar Complex In Vivo. Biophysica, 2(1), 72-78. https://doi.org/10.3390/biophysica2010007