
Silver Binding to Bacterial Glutaredoxins Observed by NMR

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Protein Expression and Purification
2.2. Chemical Shift Perturbation Studies
2.3. Size Exclusion Chromatography
2.4. Determination of τc
2.5. Protein NMR Data Collection and Resonance Assignment
2.6. Structure Calculations
2.7. Molecular Modeling of Ag Dimerized GRX
2.8. Mass Spectrometry
2.9. HEDS Assay
3. Results
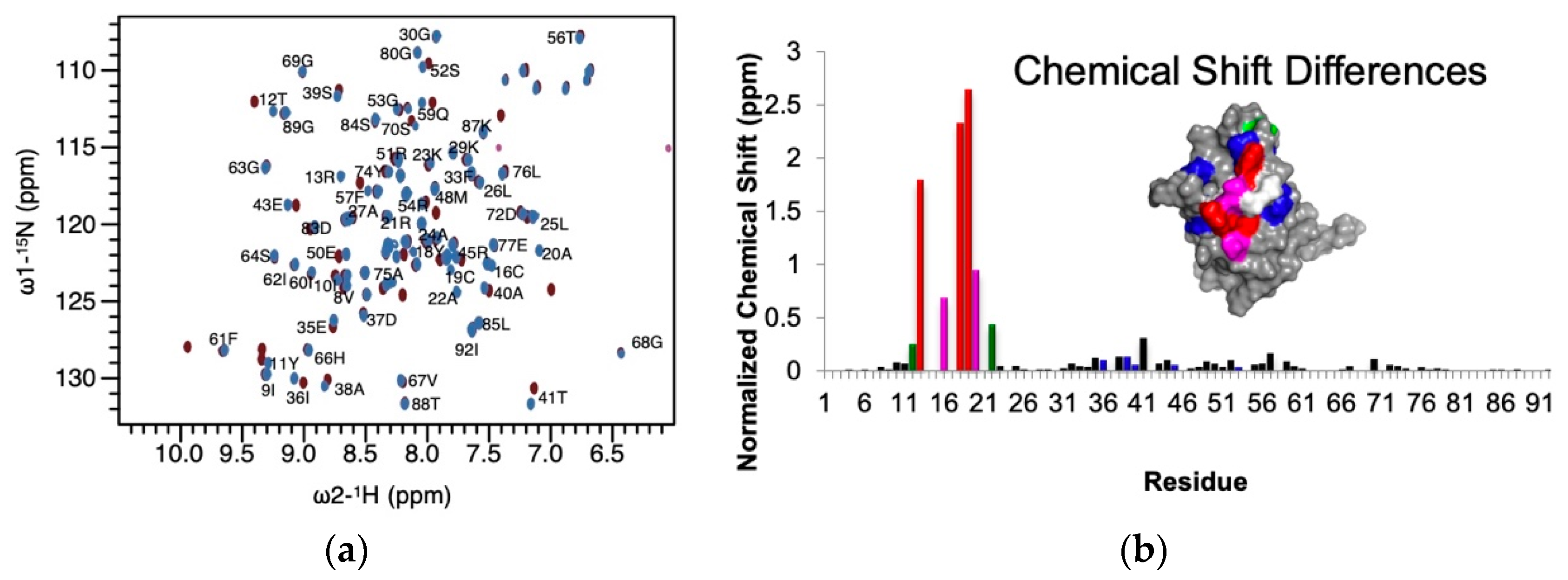
3.1. NMR Data Indicate That Multiple Metals Can Bind to the CPYC Active Site
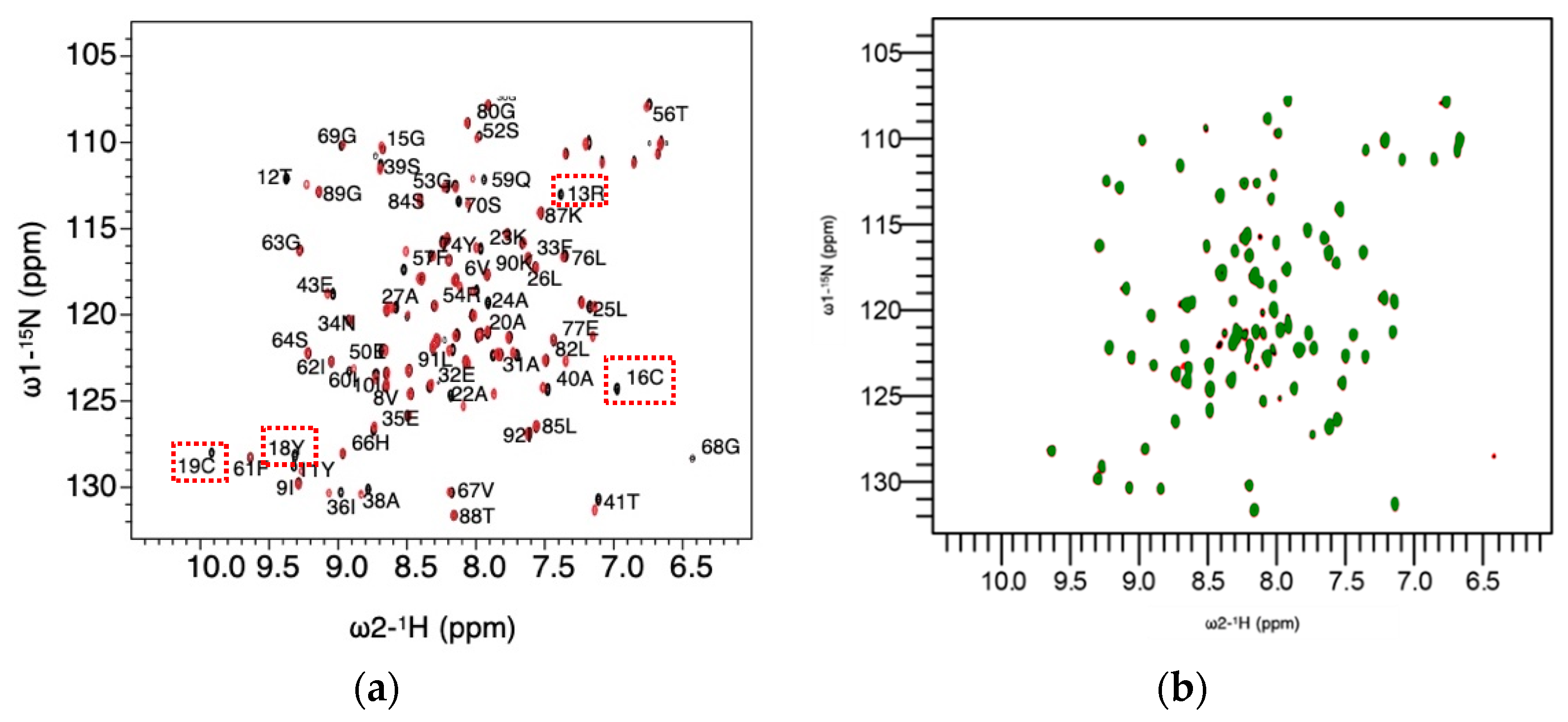
3.2. Cysteine Chemical Shifts Reveal Cysteine Bound to Silver
3.3. The NMR Structure of Silver Metalated GRX Requires Remodeling of the CPYC Active Site to Accommodate the Metal Ion
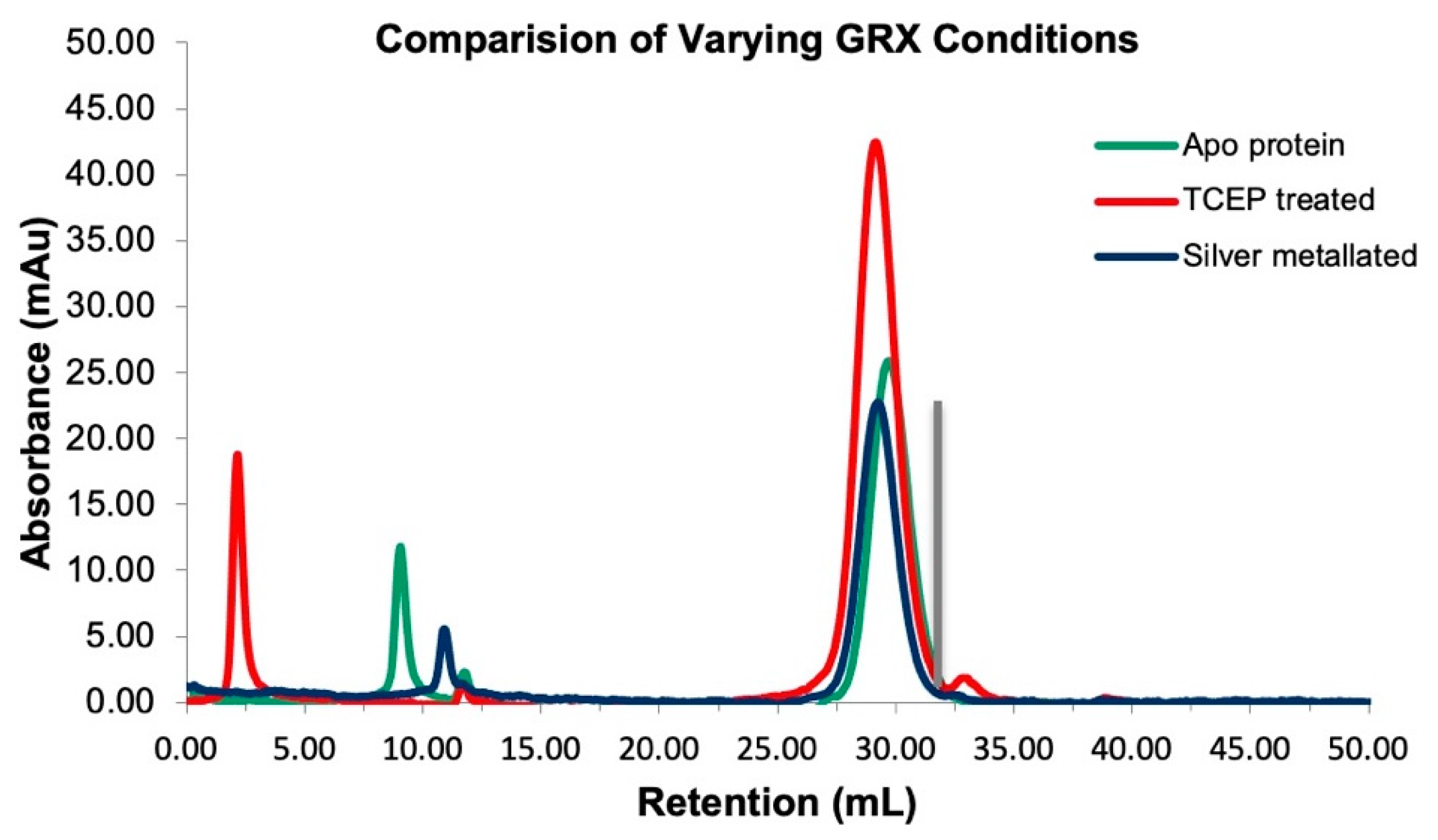
3.4. Biochemical Characterization of the Metalated Form Indicates the Protein Is a Dimer
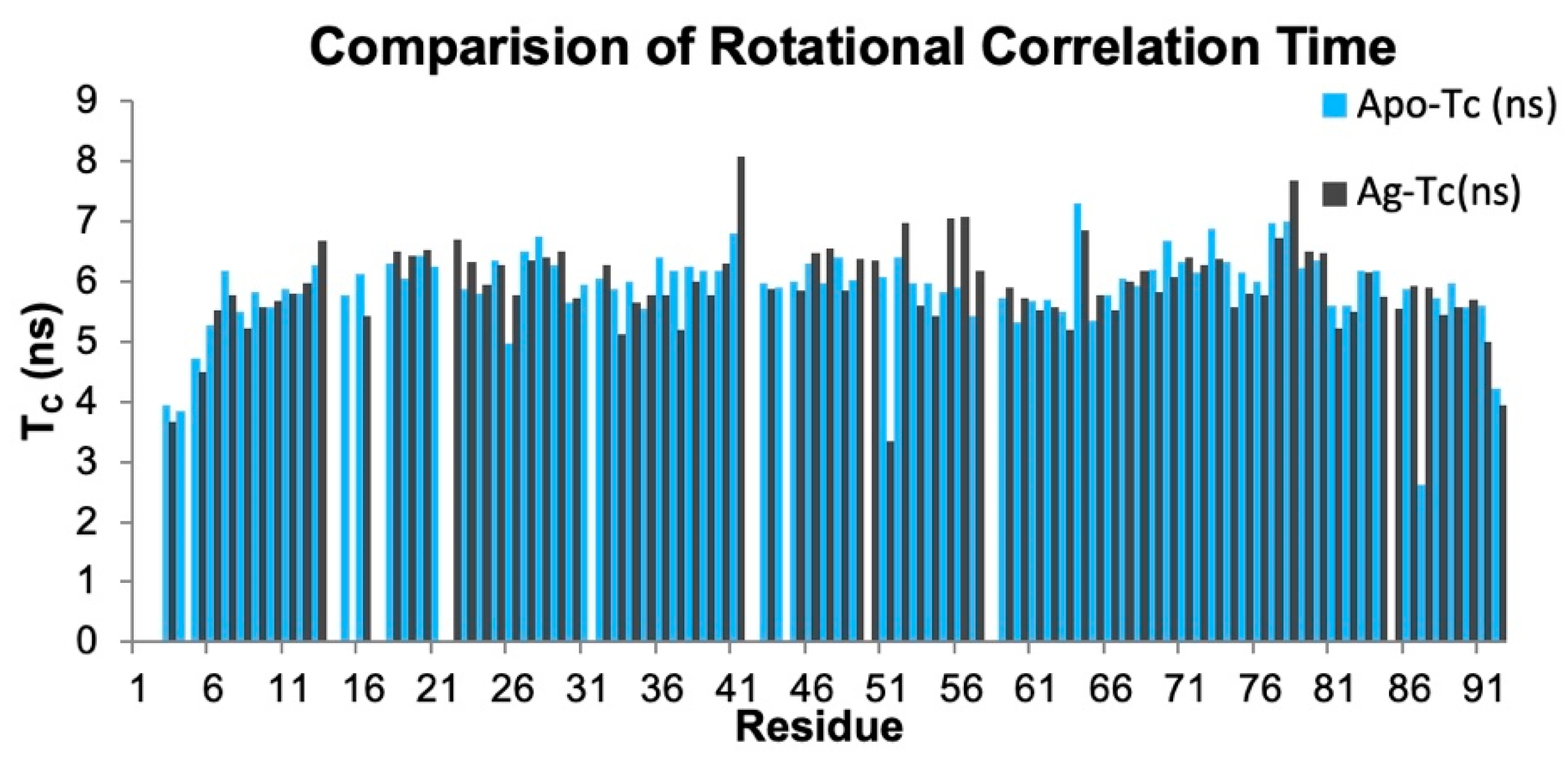
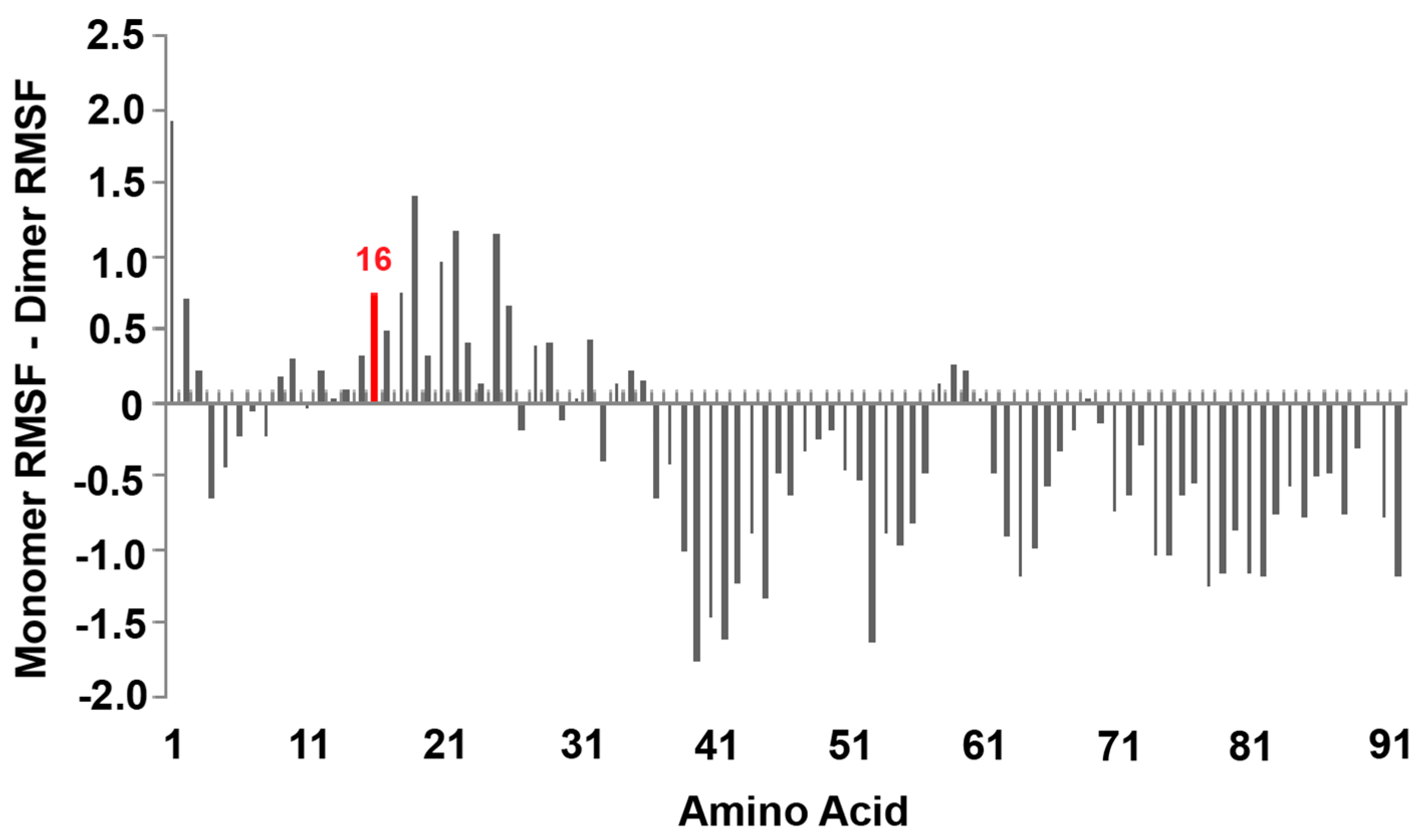
3.5. The Silver Metalated Form of the Protein Experiences an Increase in Flexibility
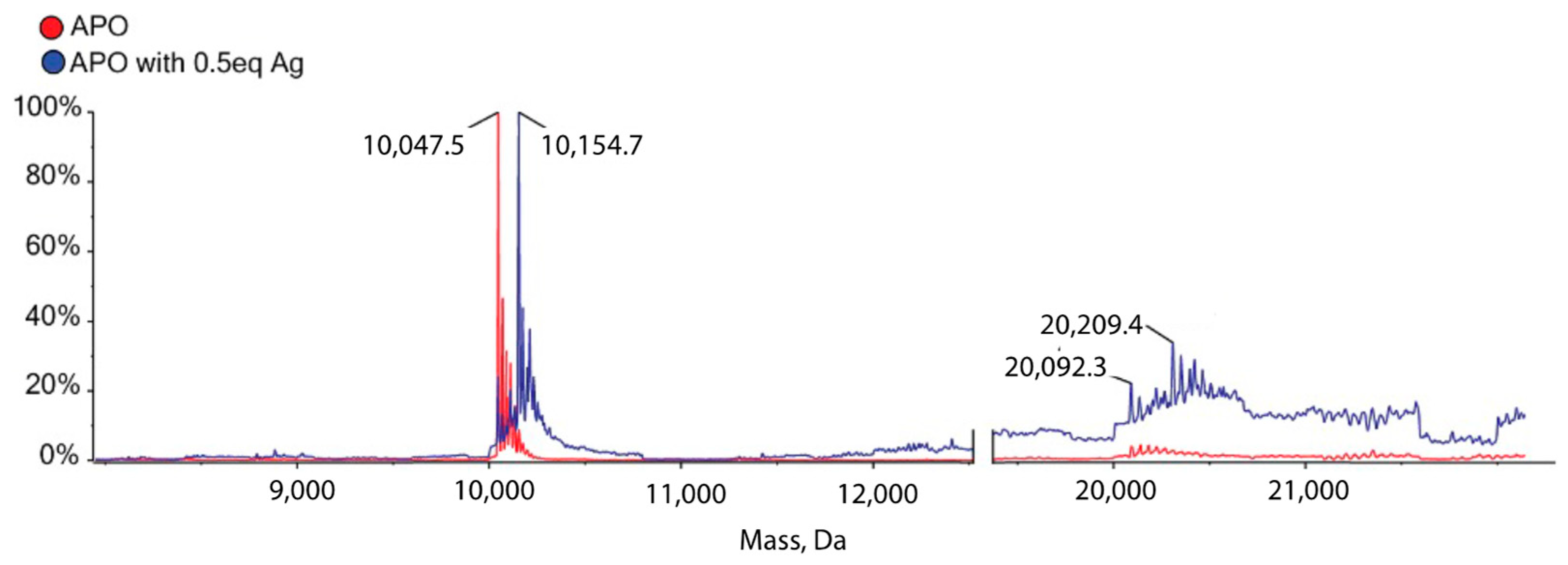
3.6. Mass-Spectrometry Reveals That the Protein Forms Metal-Mediated Dimers
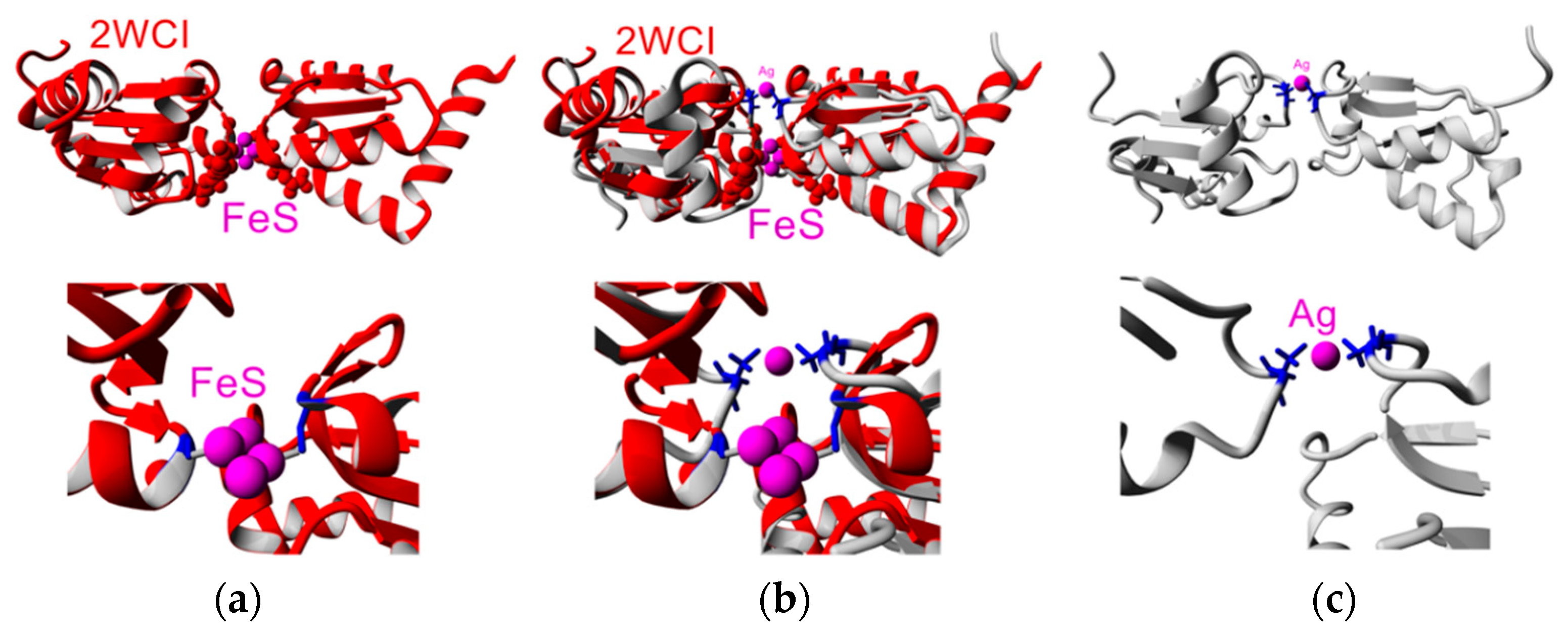
3.7. Gadolinium Reveals Dimerization Interface
3.8. Molecular Dynamics Indicates Possible Dimer Structure
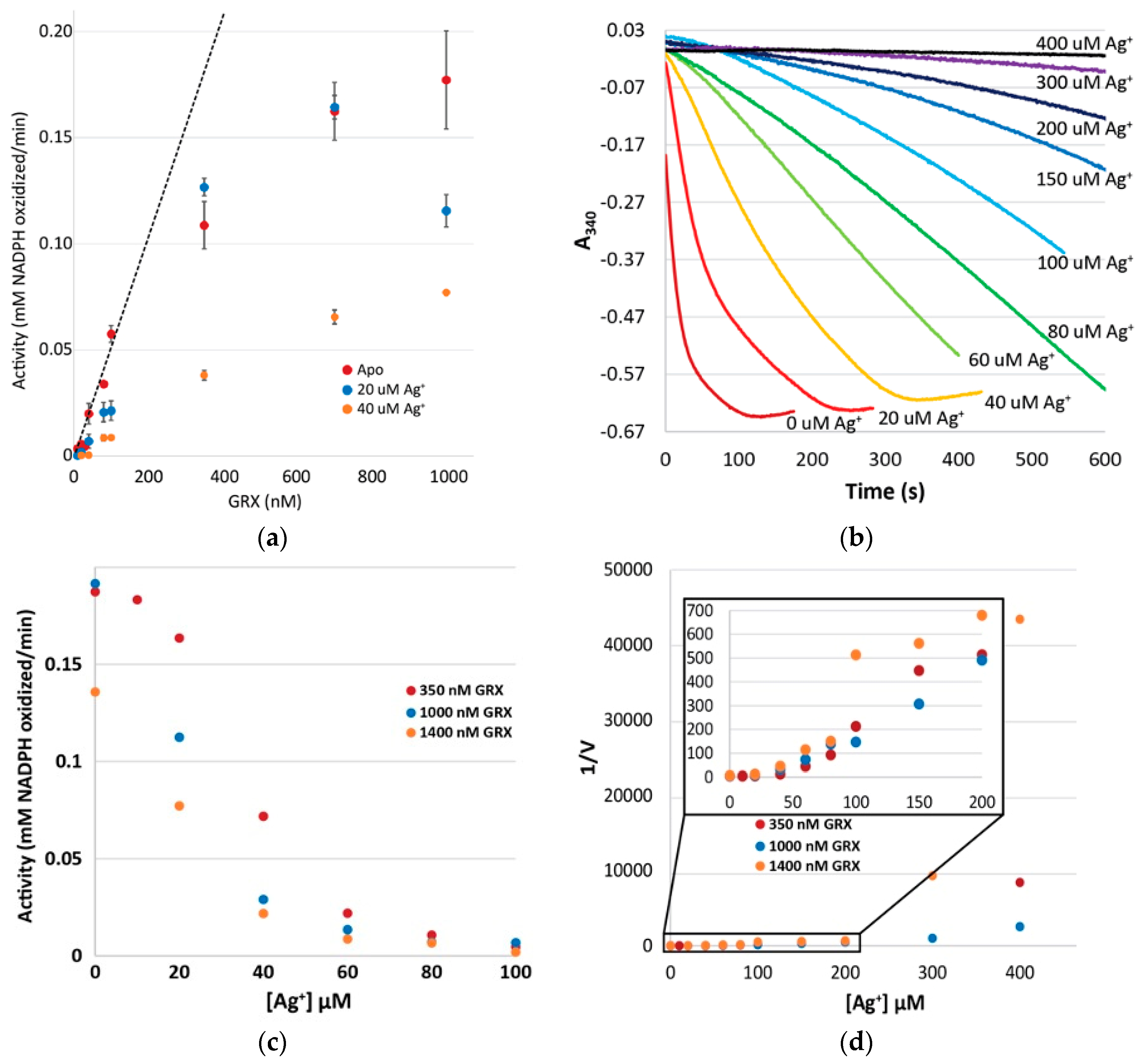
3.9. Glutathione Cannot Completely Out-Compete Metal Ions out of the Active Site
3.10. Enzymology Supports Silver-Mediated Dimer Inactivation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lillig, C.H.; Berndt, C.; Holmgren, A. Glutaredoxin systems. Biochim. Biophys. Acta 2008, 1780, 1304–1317. [Google Scholar] [CrossRef] [PubMed]
- Liedgens, L.; Zimmermann, J.; Wäschenbach, L.; Geissel, F.; Laporte, H.; Gohlke, H.; Morgan, B.; Deponte, M. Quantitative assessment of the determinant structural differences between redox-active and inactive glutaredoxins. Nat. Commun. 2020, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Trnka, D.; Engelke, A.D.; Gellert, M.; Moseler, A.; Hossain, M.F.; Lindenberg, T.T.; Pedroletti, L.; Odermatt, B.; de Souza, J.V.; Bronowska, A.K.; et al. Molecular basis for the distinct functions of redox-active and FeS-transfering glutaredoxins. Nat. Commun. 2020, 11, 1–12. [Google Scholar]
- Banci, L.; Baffoni, S.C.; Gajda, K.; Muzzioli, R.; Peruzzini, R.; Winkelmann, J. N-terminal domains mediate [2Fe-2S] cluster transfer from glutaredoxin-3 to anamorsin. Nat. Chem. Biol. 2015, 11, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Vlamis-Gardikas, A.; Åslund, F.; Holmgren, A.; Rosen, B.P. Reactivity of Glutaredoxins 1, 2, and 3 from Escherichia coli shows that Glutaredoxin 2 is the primary hydrogen donor to arsc-catalyzed arsenate reduction. J. Biol. Chem. 1999, 274, 36039–36042. [Google Scholar] [CrossRef] [PubMed]
- Gladysheva, T.B.; Oden, K.L.; Rosen, B.P. Properties of the Arsenate Reductase of Plasmid R773. Biochemistry 1994, 33, 7288–7293. [Google Scholar] [CrossRef]
- Johansson, C.; Kavanagh, K.L.; Gileadi, O.; Oppermann, U. Reversible sequestration of active site cysteines in a 2Fe-2S-bridged dimer provides a mechanism for Glutaredoxin 2 regulation in human mitochondria. J. Biol. Chem. 2007, 282, 3077–3082. [Google Scholar] [CrossRef]
- Rouhier, N.; Unno, H.; Bandyopadhyay, S.; Masip, L.; Kim, S.K.; Hirasawa, M.; Gualberto, J.M.; Lattard, V.; Kusunoki, M.; Knaff, D.B.; et al. Functional, structural, and spectroscopic characterization of a glutathione-ligated [2Fe–2S] cluster in poplar glutaredoxin C1. Proc. Natl. Acad. Sci. USA 2007, 104, 7379–7384. [Google Scholar] [CrossRef]
- Carvalho, C.; Chew, E.-H.; Hashemy, S.I.; Lu, J.; Holmgren, A. Inhibition of the human thioredoxin system. A molecular mechanism of mercury toxicity. J. Biol. Chem. 2008, 283, 11913–11923. [Google Scholar] [CrossRef]
- Lim, C.M.; Cater, M.A.; Mercer, J.F.; La Fontaine, S. Copper-dependent interaction of glutaredoxin with the N termini of the copper-ATPases (ATP7A and ATP7B) defective in Menkes and Wilson diseases. Biochem. Biophys. Res. Commun. 2006, 348, 428–436. [Google Scholar] [CrossRef]
- Hatori, Y.; Clasen, S.; Hasan, N.M.; Barry, A.; Lutsenko, S. Functional partnership of the copper export machinery and glutathione balance in human cells. J. Biol. Chem. 2012, 287, 26678–26687. [Google Scholar] [CrossRef] [PubMed]
- Singleton, W.C.; McInnes, K.T.; Cater, M.A.; Winnall, W.R.; McKirdy, R.; Yu, Y.; Taylor, P.E.; Ke, B.-X.; Richardson, D.; Mercer, J.F.; et al. Role of Glutaredoxin1 and Glutathione in regulating the activity of the copper-transporting P-type ATPases, ATP7A and ATP7B. J. Biol. Chem. 2010, 285, 27111–27121. [Google Scholar] [CrossRef] [PubMed]
- Mercer, S.W.; Burke, R. Evidence for a role for the putative Drosophila hGRX1 orthologue in copper homeostasis. BioMetals 2016, 29, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Berndt, C.; Fomenko, D.E.; Holmgren, A.; Gladyshev, V.N. A Conserved cis-proline precludes metal binding by the active site thiolates in members of the thioredoxin family of proteins. Biochemistry 2007, 46, 6903–6910. [Google Scholar] [CrossRef] [PubMed]
- Brose, J.; La Fontaine, S.; Wedd, A.G.; Xiao, Z. Redox sulfur chemistry of the copper chaperone Atox1 is regulated by the enzyme glutaredoxin 1, the reduction potential of the glutathione couple GSSG/2GSH and the availability of Cu(i). Metallomics 2014, 6, 793–808. [Google Scholar] [CrossRef]
- Maghool, S.; La Fontaine, S.; Roberts, B.R.; Kwan, A.H.; Maher, M.J. Human glutaredoxin-1 can transfer copper to isolated metal binding domains of the P1B-type ATPase, ATP7B. Sci. Rep. 2020, 10, 4157. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, B.; Uzarska, M.A.; Berndt, C.; Godoy, J.R.; Haunhorst, P.; Lillig, C.H.; Lill, R.; Mühlenhoff, U. The multidomain thioredoxin-monothiol glutaredoxins represent a distinct functional group. Antioxid. Redox Signal 2011, 15, 19–30. [Google Scholar] [CrossRef]
- Manta, B.; Pavan, C.; Sturlese, M.; Medeiros, A.; Crispo, M.; Berndt, C.; Krauth-Siegel, R.L.; Bellanda, M.; Comini, M.A. Iron–sulfur cluster binding by mitochondrial monothiol glutaredoxin-1 of Trypanosoma brucei: Molecular basis of iron–sulfur cluster coordination and relevance for parasite infectivity. Antioxid. Redox Signal 2013, 19, 665–682. [Google Scholar] [CrossRef]
- Roret, T.; Zhang, B.; Moseler, A.; Dhalleine, T.; Gao, X.H.; Couturier, J.; Lemaire, S.D.; Didierjean, C.; Johnson, M.K.; Rouhier, N. Atypical Iron-Sulfur Cluster Binding, Redox Activity and Structural Properties of Chlamydomonas reinhardtii Glutaredoxin 2. Antioxidants 2021, 10, 803. [Google Scholar] [CrossRef]
- Ceylan, S.; Seidel, V.; Ziebart, N.; Berndt, C.; Dirdjaja, N.; Krauth-Siegel, R.L. The dithiol glutaredoxins of African trypano-somes have distinct roles and are closely linked to the unique trypanothione metabolism. J. Biol. Chem. 2010, 285, 35224–35237. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; La Fontaine, S.; Bush, A.I.; Wedd, A.G. Molecular mechanisms of glutaredoxin enzymes: Versatile hubs for thiol–disulfide exchange between protein thiols and glutathione. J. Mol. Biol. 2018, 431, 158–177. [Google Scholar] [CrossRef]
- Fox, C.L., Jr. Silver sulfadiazine—A new topical therapy for Pseudomonas in burns. Therapy of Pseudomonas infection in burns. Arch. Surg. 1968, 96, 184–188. [Google Scholar] [CrossRef]
- Wright, B.D.; Shah, P.N.; McDonald, L.J.; Shaeffer, M.L.; Wagers, P.O.; Panzner, M.J.; Smolen, J.; Tagaev, J.; Tessier, C.A.; Cannon, C.; et al. Synthesis, characterization, and antimicrobial activity of silver carbene complexes derived from 4,5,6,7-tetrachlorobenzimidazole against antibiotic resistant bacteria. Dalton Trans. 2012, 41, 6500–6506. [Google Scholar] [CrossRef] [PubMed]
- Kascatan-Nebioglu, A.; Melaiye, A.; Hindi, K.; Durmus, S.; Panzner, M.J.; Hogue, L.A.; Mallett, R.J.; Hovis, C.E.; Coughenour, M.; Crosby, S.D.; et al. Synthesis from caffeine of a MixedN-heterocyclic carbene−silver acetate complex active against resistant respiratory pathogens. J. Med. Chem. 2006, 49, 6811–6818. [Google Scholar] [CrossRef] [PubMed]
- Frei, A.; Zuegg, J.; Elliott, A.G.; Baker, M.V.; Braese, S.; Brown, C.; Chen, F.; Dowson, C.G.; Dujardin, G.; Jung, N.; et al. Metal complexes as a promising source for new antibiotics. Chem. Sci. 2020, 11, 2627–2639. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, M.A.; O’Gara, J.P.; Gordon, S.V.; Hackenberg, F.; Healy, C.; Paradisi, F.; Patil, S.; Schaible, B.; Tacke, M. Investigations into the antibacterial activity of the silver-based antibiotic drug candidate SBC. Antibiotics 2012, 1, 25–28. [Google Scholar] [CrossRef]
- Palomero, O.E.; Cunningham, A.L.; Davies, B.W.; Jones, R.A. Antibacterial thiamine inspired silver (I) and gold (I) N-heterocyclic carbene compounds. Inorg. Chim. Acta 2020, 517, 120152. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, T.J.; Deblock, M.C.; Hindi, K.M.; Durmus, S.; Panzner, M.J.; Tessier, C.A.; Youngs, W.J. Synthesis and anticancer properties of gold(I) and silver(I) N-heterocyclic carbene complexes. J. Organomet. Chem. 2011, 696, 1066–1071. [Google Scholar] [CrossRef]
- Holmes, J.; Kearsey, R.J.; Paske, K.A.; Singer, F.N.; Atallah, S.; Pask, C.M.; Phillips, R.M.; Willans, C.E. Tethered N-heterocyclic carbene-carboranyl silver complexes for cancer therapy. Organometallics 2019, 38, 2530–2538. [Google Scholar] [CrossRef]
- Ceramella, J.; Mariconda, A.; Iacopetta, D.; Saturnino, C.; Barbarossa, A.; Caruso, A.; Rosano, C.; Sinicropi, M.S.; Longo, P. From coins to cancer therapy: Gold, silver and copper complexes targeting human topoisomerases. Bioorganic Med. Chem. Lett. 2020, 30, 126905. [Google Scholar] [CrossRef]
- Kankala, S.; Thota, N.; Björkling, F.; Taylor, M.K.; Vadde, R.; Balusu, R. Silver carbene complexes: An emerging class of anti-cancer agents. Drug Dev. Res. 2019, 80, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Morones-Ramirez, J.R.; Winkler, J.A.; Spina, C.; Collins, J.J. Silver enhances antibiotic activity against gram-negative bacteria. Sci. Transl. Med. 2013, 5, 190ra81. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-J.; Kim, J.Y.; Kim, J.; Lee, J.-H.; Hahn, J.-S.; Gu, M.B.; Yoon, J. Silver-ion-mediated reactive oxygen species generation affecting bactericidal activity. Water Res. 2009, 43, 1027–1032. [Google Scholar] [CrossRef]
- Gordon, O.; Slenters, T.V.; Brunetto, P.S.; Villaruz, A.E.; Sturdevant, D.E.; Otto, M.; Landmann, R.; Fromm, K.M. Silver coordination polymers for prevention of implant infection: Thiol interaction, impact on respiratory chain enzymes, and hydroxyl radical induction. Antimicrob. Agents Chemother. 2010, 54, 4208–4218. [Google Scholar] [CrossRef] [PubMed]
- Das, B.; Dash, S.K.; Mandal, D.; Ghosh, T.; Chattopadhyay, S.; Tripathy, S.; Das, S.; Dey, S.K.; Das, D.; Roy, S. Green synthesized silver nanoparticles destroy multidrug resistant bacteria via reactive oxygen species mediated membrane damage. Arab. J. Chem. 2017, 10, 862–876. [Google Scholar] [CrossRef]
- Liau, S.Y.; Read, D.C.; Pugh, W.J.; Furr, J.R.; Russell, A.D. Interaction of silver nitrate with readily identifiable groups: Rela-tionship to the antibacterial action of silver ions. Lett. Appl. Microbiol. 1997, 25, 279–283. [Google Scholar] [CrossRef]
- Nagy, A.; Harrison, A.; Sabbani, S.; Munson, R.S., Jr.; Dutta, P.K.; Waldman, W.J. Silver nanoparticles embedded in zeolite membranes: Release of silver ions and mechanism of antibacterial action. Int. J. Nanomed. 2011, 6, 1833–1852. [Google Scholar]
- Sattler, M.; Schleucher, J.; Griesinger, C. Heteronuclear multidimensional NMR experiments for the structure determination of proteins in solution employing pulsed field gradients. Prog. Nucl. Magn. Reson. Spectrosc. 1999, 34, 93–158. [Google Scholar] [CrossRef]
- Delaglio, F.; Grzesiek, S.; Vuister, G.; Zhu, G.; Pfeifer, J.; Bax, A. NMRPipe: A multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR 1995, 6, 277–293. [Google Scholar] [CrossRef]
- Vranken, W.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins Struct. Funct. Bioinform. 2005, 59, 687–696. [Google Scholar] [CrossRef]
- Cheung, M.-S.; Maguire, M.L.; Stevens, T.J.; Broadhurst, R.W. DANGLE: A Bayesian inferential method for predicting protein backbone dihedral angles and secondary structure. J. Magn. Reson. 2010, 202, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Güntert, P.; Mumenthaler, C.; Wüthrich, K. Torsion angle dynamics for NMR structure calculation with the new program Dyana. J. Mol. Biol. 1997, 273, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Güntert, P. Automated NMR structure calculation with CYANA. In Protein NMR Techniques; Humana Press: Totowa, NJ, USA, 2004; pp. 353–378. [Google Scholar]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.M.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Konagurthu, A.; Whisstock, J.; Stuckey, P.; Lesk, A. MUSTANG: A multiple structural alignment algorithm. Proteins Struct. Funct. Bioinform. 2006, 64, 559–574. [Google Scholar] [CrossRef]
- Morris, G.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Duan, Y.; Wu, C.; Chowdhury, S.; Lee, M.C.; Xiong, G.; Zhang, W.; Yang, R.; Cieplak, P.; Luo, R.; Lee, T.; et al. A point-charge force field for molecular mechanics simulations of proteins based on condensed-phase quantum mechanical calculations. J. Comput. Chem. 2003, 24, 1999–2012. [Google Scholar] [CrossRef]
- Nagai, S.; Black, S. A Thiol-disulfide transhydrogenase from yeast. J. Biol. Chem. 1968, 243, 1942–1947. [Google Scholar] [CrossRef]
- Zaffagnini, M.; Michelet, L.; Massot, V.; Trost, P.; Lemaire, S. Biochemical characterization of glutaredoxins from chlamydomonas reinhardtii reveals the unique properties of a chloroplastic CGFS-type glutaredoxin. J. Biol. Chem. 2008, 283, 8868–8876. [Google Scholar] [CrossRef]
- Gitschier, J.; Moffat, B.; Reilly, D.; Wood, W.I.; Fairbrother, W.J. Solution structure of the fourth metal-binding domain from the Menkes copper-transporting ATPase. Nat. Genet. 1998, 5, 47–54. [Google Scholar] [CrossRef]
- Changela, A.; Chen, K.; Xue, Y.; Holschen, J.; Outten, C.E.; O’Halloran, T.V.; Mondragón, A. Molecular basis of metal-ion selectivity and zeptomolar sensitivity by CueR. Science 2003, 301, 1383–1387. [Google Scholar] [CrossRef]
- Xue, Y.; Davis, A.V.; Balakrishnan, G.; Stasser, J.P.; Staehlin, B.M.; Focia, P.; Spiro, T.G.; Penner-Hahn, J.E.; O’Halloran, T.V. Cu(I) recognition via cation-pi and methio-nine interactions in CusF. Nat. Chem. Biol. 2008, 4, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Long, F.; Su, C.-C.; Zimmermann, M.; Boyken, S.; Rajashankar, K.R.; Jernigan, R.L.; Yu, E.W. Crystal structures of the CusA efflux pump suggest methionine-mediated metal transport. Nature 2010, 467, 484–488. [Google Scholar] [CrossRef]
- Panzner, M.J.; Bilinovich, S.M.; Parker, J.A.; Bladholm, E.L.; Ziegler, C.J.; Berry, S.M.; Leeper, T.C. Isomorphic deactivation of a Pseudomonas aeruginosa oxidoreductase: The crystal structure of Ag(I) metallated azurin at 1.7 Å. J. Inorg. Biochem. 2013, 128, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Loftin, I.R.; Franke, S.; Blackburn, N.J.; McEvoy, M.M. Unusual Cu(I)/Ag(I) coordination of Escherichia coliCusF as revealed by atomic resolution crystallography and X-ray absorption spectroscopy. Protein Sci. 2007, 16, 2287–2293. [Google Scholar] [CrossRef]
- Su, C.-C.; Yang, F.; Long, F.; Reyon, D.; Routh, M.D.; Kuo, D.W.; Mokhtari, A.K.; Van Ornam, J.D.; Rabe, K.L.; Hoy, J.A.; et al. Crystal structure of the membrane fusion protein CusB from Escherichia coli. J. Mol. Biol. 2009, 393, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Roberts, S.A.; McDevitt, S.F.; Weichsel, A.; Wildner, G.F.; Grass, G.B.; Rensing, C.; Montfort, W.R. Crystal structures of multicopper oxidase CueO Bound to Copper(I) and Silver(I): Functional role of a methionine-rich sequence. J. Biol. Chem. 2011, 286, 37849–37857. [Google Scholar] [CrossRef]
- Anastassopoulou, I.; Banci, L.; Bertini, I.; Cantini, F.; Katsari, E.; Rosato, A. Solution structure of the Apo and Copper(I)-loaded human metallochaperone HAH1. Biochemistry 2004, 43, 13046–13053. [Google Scholar] [CrossRef]
- Brouwer, M.; Brouwer-Hoexum, T. Interaction of copper-metallothionein from the American lobster, Homarus americanus, with glutathione. Arch. Biochem. Biophys. 1991, 290, 207–213. [Google Scholar] [CrossRef]
- Sharma, D.; Rajarathnam, K. 13C NMR chemical shifts can predict disulfide bond formation. J. Biomol. NMR 2000, 18, 165–171. [Google Scholar] [CrossRef]
- Kornhaber, G.J.; Snyder, D.; Moseley, H.N.B.; Montelione, G.T. Identification of zinc-ligated cysteine residues based on 13Calpha and 13Cbeta chemical shift data. J. Biomol. NMR. 2006, 34, 259–269. [Google Scholar] [CrossRef]
- Leeper, T.; Zhang, S.; Van Voorhis, W.C.; Myler, P.J.; Varani, G. Comparative analysis of glutaredoxin domains from bacterial opportunistic pathogens. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Iwema, T.; Picciocchi, A.; Traore, D.; Ferrer, J.-L.; Chauvat, F.; Jacquamet, L. Structural basis for delivery of the intact [Fe2S2] cluster by monothiol glutaredoxin. Biochemistry 2009, 48, 6041–6043. [Google Scholar] [CrossRef] [PubMed]
- Johansson, C.; Roos, A.K.; Montano, S.J.; Sengupta, R.; Filippakopoulos, P.; Guo, K.; Von Delft, F.; Holmgren, A.; Oppermann, U.; Kavanagh, K.L. The crystal structure of human GLRX5: Iron–sulfur cluster co-ordination, tetrameric assembly and monomer activity. Biochem. J. 2011, 433, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Couturier, J.; Ströher, E.; Albetel, A.-N.; Roret, T.; Muthuramalingam, M.; Tarrago, L.; Seidel, T.; Tsan, P.; Jacquot, J.P.; Johnson, M.K.; et al. Arabidopsis chloroplastic glu-taredoxin C5 as a model to explore molecular determinants for iron-sulfur cluster binding into glutaredoxins. J. Biol. Chem. 2011, 286, 27515–27527. [Google Scholar] [CrossRef] [PubMed]
- Erickson, H.P. Size and shape of protein molecules at the nanometer level determined by sedimentation, gel filtration, and electron microscopy. Biol. Proced. Online 2009, 11, 32–51. [Google Scholar] [CrossRef]
- Banci, L.; Brancaccio, D.; Baffoni, S.C.; Del Conte, R.; Gadepalli, R.; Mikolajczyk, M.; Neri, S.; Piccioli, M.; Winkelmann, J. [2Fe-2S] cluster transfer in iron-sulfur protein biogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 6203–6208. [Google Scholar] [CrossRef]
- Rossi, P.; Swapna, G.V.T.; Huang, Y.J.; Aramini, J.M.; Anklin, C.; Conover, K.; Hamilton, K.; Xiao, R.; Acton, T.B.; Ertekin, A.; et al. A microscale protein NMR sample screening pipeline. J. Biomol. NMR 2009, 46, 11–22. [Google Scholar] [CrossRef]
- Noguera, V.; Walker, O.; Rouhier, N.; Jacquot, J.-P.; Krimm, I.; Lancelin, J.-M. NMR reveals a novel glutaredoxin–glutaredoxin interaction interface. J. Mol. Biol. 2005, 353, 629–641. [Google Scholar] [CrossRef]
- Petros, A.M.; Mueller, L.; Kopple, K.D. NMR identification of protein surfaces using paramagnetic probes. Biochemistry 1990, 29, 10041–10048. [Google Scholar] [CrossRef]
- Liepinsh, E.; Baryshev, M.; Sharipo, A.; Ingelman-Sundberg, M.; Otting, G.; Mkrtchian, S. Thioredoxin fold as homodimeri-zation module in the putative chaperone ERp29: NMR structures of the domains and experimental model of the 51 kDa dimer. Structure 2001, 9, 457–471. [Google Scholar] [CrossRef]
- Almeida, R.M.; Geraldes, C.F.G.C.; Pauleta, S.R.; Moura, J.J.G. Gd(III) Chelates as NMR probes of protein–protein interactions. Case study: Rubredoxin and Cytochrome c3. Inorg. Chem. 2011, 50, 10600–10607. [Google Scholar] [CrossRef] [PubMed]
- Vlamis, A.; Åslund, F.; Spyrou, G.; Bergman, T.; Holmgren, A. Cloning, overexpression, and characterization of glutaredoxin 2, an atypical glutaredoxin from Escherichia coli. J. Biol. Chem. 1997, 272, 11236–11243. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Zhou, S.-M.; Xia, Y.; Zou, H.; Meng, F.; Yan, Y.-B. Inactivation and unfolding of the hyperthermophilic inorganic pyrophosphatase from thermus thermophilus by sodium dodecyl sulfate. Int. J. Mol. Sci. 2009, 10, 2849–2859. [Google Scholar] [CrossRef] [PubMed]
- Lorey, S.; Stöckel-Maschek, A.; Faust, J.; Brandt, W.; Stiebitz, B.; Gorrell, M.D.; Kähne, T.; Mrestani-Klaus, C.; Wrenger, S.; Reinhold, D.; et al. Different modes of dipeptidyl peptidase IV (CD26) inhibition by oligopeptides derived from the N-terminus of HIV-1 Tat indicate at least two inhibitor binding sites. JBIC J. Biol. Inorg. Chem. 2003, 270, 2147–2156. [Google Scholar] [CrossRef]
- Sondi, I.; Salopek-Sondi, B. Silver nanoparticles as antimicrobial agent: A case study on E. coli as a model for Gram-negative bacteria. J. Colloid Interface Sci. 2004, 275, 177–182. [Google Scholar] [CrossRef]
- Ibricevic, A.; Brody, S.L.; Youngs, W.J.; Cannon, C. ATP7B detoxifies silver in ciliated airway epithelial cells. Toxicol. Appl. Pharmacol. 2010, 243, 315–322. [Google Scholar] [CrossRef]
- Wernimont, A.K.; Huffman, D.L.; Lamb, A.; O’Halloran, T.V.; Rosenzweig, A.C. Structural basis for copper transfer by the metallochaperone for the Menkes/Wilson disease proteins. Nat. Struct. Mol. Biol. 2000, 7, 766–771. [Google Scholar]
- Kippner, L.E.; Finn, N.A.; Shukla, S.; Kemp, M.L. Systemic remodeling of the redox regulatory network due to RNAi pertur-bations of glutaredoxin 1, thioredoxin 1, and glucose-6-phosphate dehydrogenase. BMC Syst. Biol. 2011, 5, 164. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal | Binds? | Binding Residues | Exchange |
|---|---|---|---|
| Ag(I) | Yes | CPYC | Slow |
| Cu(I) | Yes | CPYC | Slow |
| Cu(II) | Yes | CPYC | Slow |
| Hg(II) | Yes | CPYC | Slow |
| Co(II) | No | N/A | N/A |
| Ni(II) | No | N/A | N/A |
| Zn(II) | Possibly, precipitates | N/A | N/A |
| Restraints Statistics | |
| Distances | 1299 |
| Short | 685 |
| Medium | 258 |
| Long | 356 |
| Dihedral | 52 |
| H-Bonds | 104 |
| Ensemble Convergence | |
| Backbone RMSD | 0.6 ± 0.16 |
| Heavy Atom RMSD | 1.04 ± 0.14 |
| Ramachandran Plot Statistics | |
| Most Favored % | 79.6 |
| Additionally Allowed % | 18.9 |
| Generously Allowed % | 1.4 |
| Disallowed % | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilinovich, S.M.; Morris, D.L.; Prokop, J.W.; Caporoso, J.A.; Taraboletti, A.; Duangjumpa, N.; Panzner, M.J.; Shriver, L.P.; Leeper, T.C. Silver Binding to Bacterial Glutaredoxins Observed by NMR. Biophysica 2021, 1, 359-376. https://doi.org/10.3390/biophysica1040027
Bilinovich SM, Morris DL, Prokop JW, Caporoso JA, Taraboletti A, Duangjumpa N, Panzner MJ, Shriver LP, Leeper TC. Silver Binding to Bacterial Glutaredoxins Observed by NMR. Biophysica. 2021; 1(4):359-376. https://doi.org/10.3390/biophysica1040027
Chicago/Turabian StyleBilinovich, Stephanie M., Daniel L. Morris, Jeremy W. Prokop, Joel A. Caporoso, Alexandra Taraboletti, Nilubol Duangjumpa, Matthew J. Panzner, Leah P. Shriver, and Thomas C. Leeper. 2021. "Silver Binding to Bacterial Glutaredoxins Observed by NMR" Biophysica 1, no. 4: 359-376. https://doi.org/10.3390/biophysica1040027
APA StyleBilinovich, S. M., Morris, D. L., Prokop, J. W., Caporoso, J. A., Taraboletti, A., Duangjumpa, N., Panzner, M. J., Shriver, L. P., & Leeper, T. C. (2021). Silver Binding to Bacterial Glutaredoxins Observed by NMR. Biophysica, 1(4), 359-376. https://doi.org/10.3390/biophysica1040027