
Cytotoxicity of Quillaja saponaria Saponins towards Lung Cells Is Higher for Cholesterol-Rich Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Lines
2.3. Cholesterol Content
2.4. Microfluidic Setup
2.5. Cytotoxicity Measurement in the Micro Scale
2.6. Cytotoxicity Measurement in the Macro Scale Using MTT Test
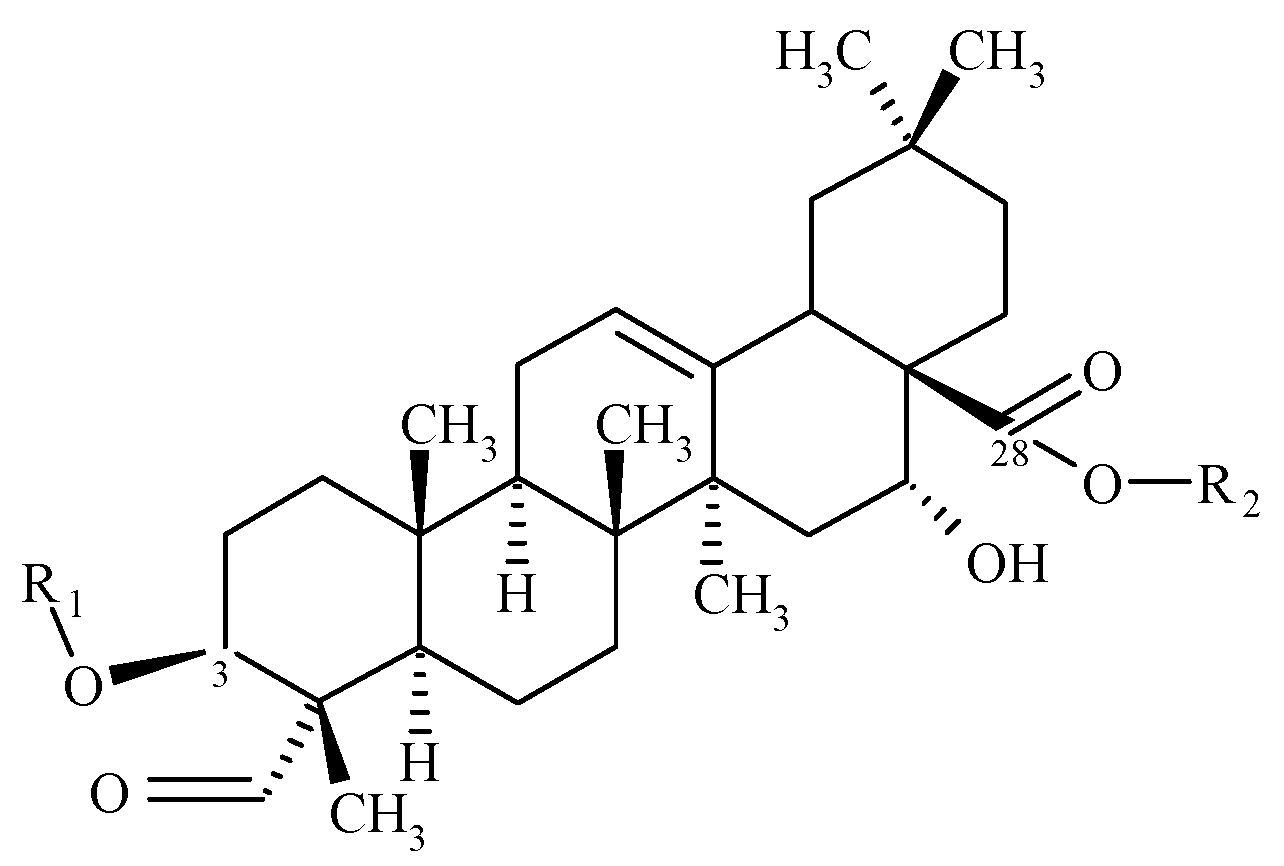
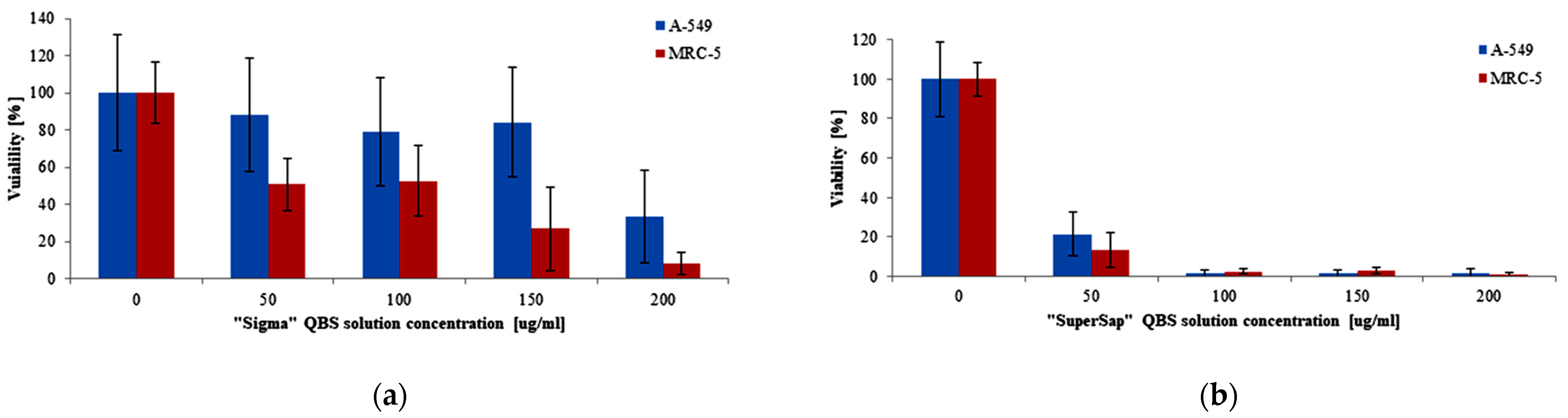
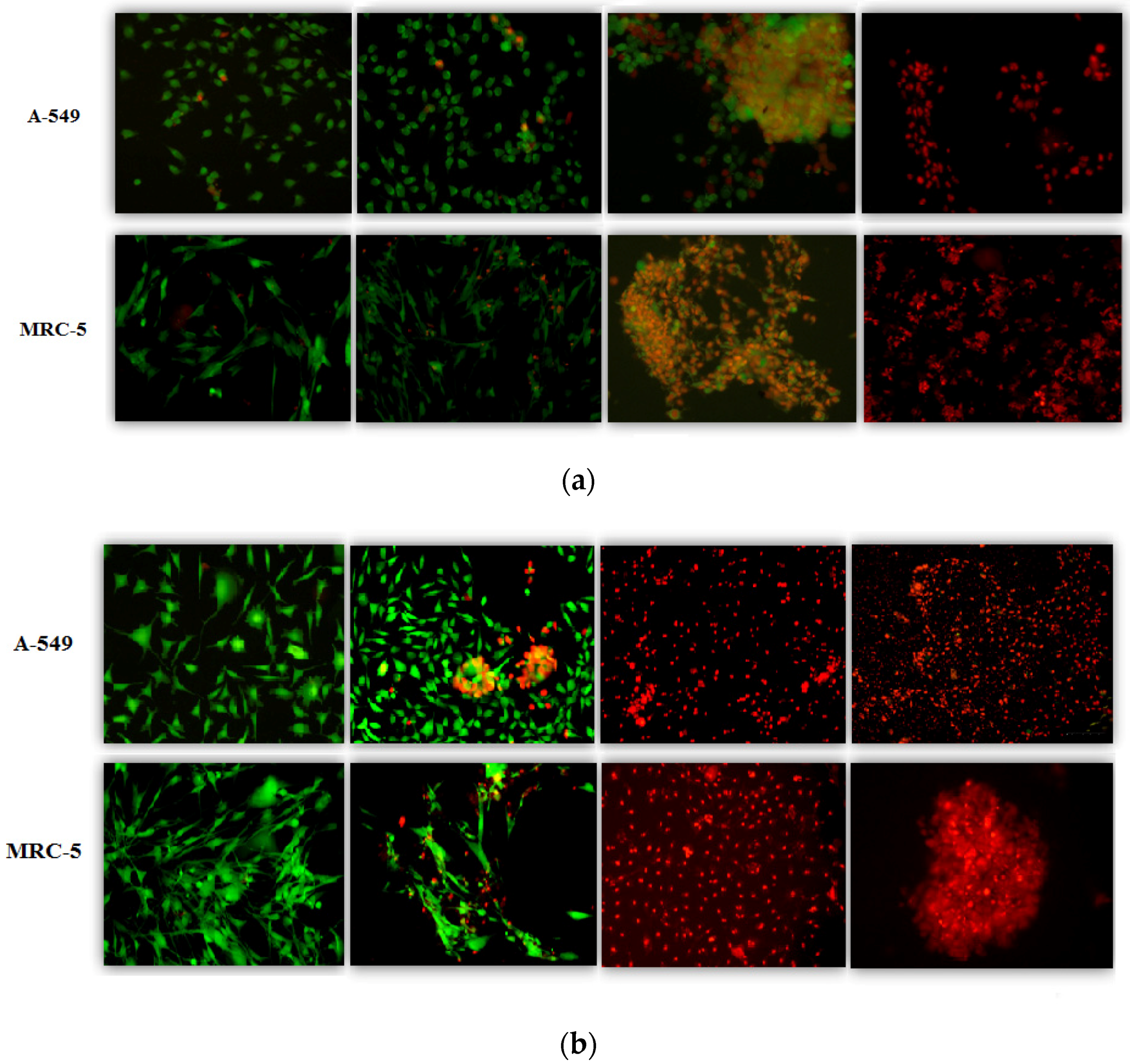
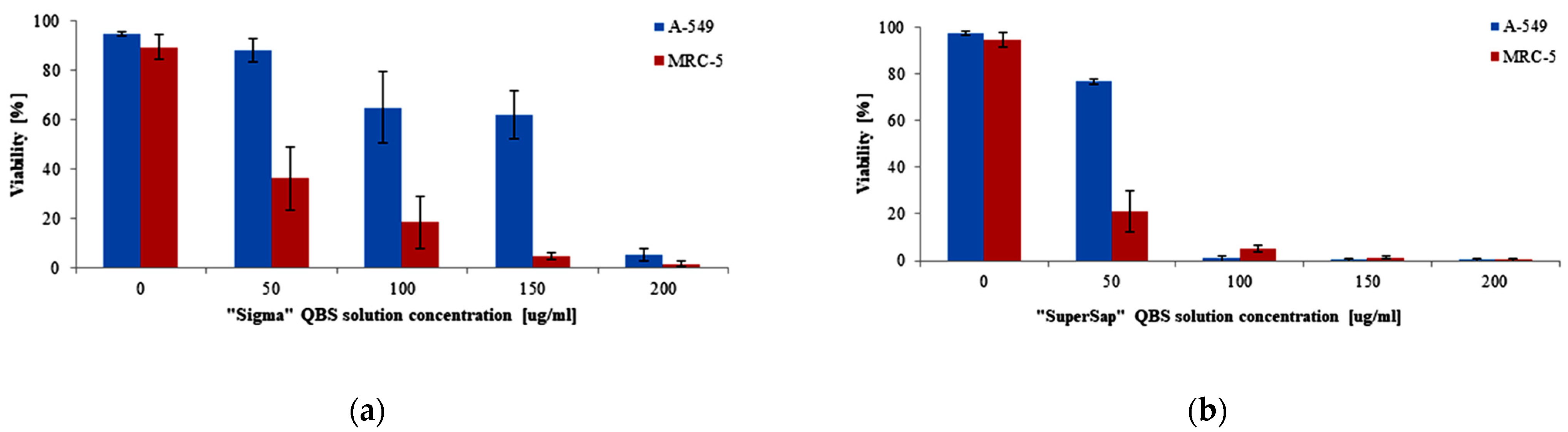
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Francis, G.; Kerem, Z.; Makkar, H.P.S.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587. [Google Scholar] [CrossRef] [PubMed]
- Szakiel, A.; Pączkowski, C.; Henry, M. Influence of environmental biotic factors on the content of saponins in plants. Phytochem. Rev. 2011, 10, 493–502. [Google Scholar] [CrossRef]
- Gus-Mayer, S.; Brunner, H.; Schneider-Poetsch, H.A.W.; Ruediger, W. Avenacosidase from oat: Purification, sequence analysis and biochemical characterization of a new member of the BGA family of b-glucosidases. Plant Mol. Biol. 1994, 26, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Weng, A.; Manunta, M.D.I.; Thakur, M.; Gilabert-Oriol, R.; Tagalakis, A.D.; Eddaoudi, A.; Munye, M.M.; Vink, C.A.; Wiesner, B.; Eichhorst, J.; et al. Improved intracellular delivery of peptide- and lipid-nanoplexes by natural glycosides. J. Control. Release 2015, 206, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, H.; Weng, A.; Gilabert-Oriol, R. Augmenting the efficacy of immunotoxins and other targeted protein toxins by endosomal escape enhancers. Toxins 2016, 8, 200. [Google Scholar] [CrossRef]
- Böttger, S.; Hofmann, K.; Melzig, M.F. Saponins can perturb biologic membranes and reduce the surface tension of aqueous solutions: A correlation? Bioorg. Med. Chem. 2012, 20, 2822–2828. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowski, K.; Piotrowski, M.; Popielarz, W.; Sosnowski, T.R. Short- and mid-term adsorption behaviour of Quillaja Bark Saponin and its mixtures with lysozyme. Food Hydrocoll. 2011, 25. [Google Scholar] [CrossRef]
- Mitra, S.; Dungan, S.R. Micellar Properties of Quillaja Saponin. 1. Effects of Temperature, Salt, and pH on Solution Properties. J. Agric. Food Chem. 1997. [Google Scholar] [CrossRef]
- Copaja, S.V.; Blackburn, C.; Carmona, R. Variation of saponin contents in Quillaja saponica molina. Wood Sci. Technol. 2003, 37, 103–108. [Google Scholar] [CrossRef]
- Martín, R.S.; Briones, R. Industrial Uses and Sustainable Supply of Quillaja saponaria (Rosaceae) Saponins. Econ. Bot. 1999, 53, 302–311. [Google Scholar] [CrossRef]
- Güçlü-Üstündağ, Ö.; Mazza, G. Saponins: Properties, Applications and Processing. Crit. Rev. Food Sci. Nutr. 2007, 47, 231–258. [Google Scholar] [CrossRef] [PubMed]
- San Martin, R.; Briones, R. Quality control of commercial quillaja (Quillaja saponaria Molina) extracts by reverse phase HPLC. J. Sci. Food Agric. 2000, 80, 2063–2068. [Google Scholar] [CrossRef]
- Ragupathi, G.; Gardner, J.R.; Livingston, P.O.; Gin, D.Y. Natural and synthetic saponin adjuvant QS-21 for vaccines against cancer. Expert Rev. Vaccines 2011, 10, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowski, K.; Orczyk, M.; Gutberlet, T.; Trapp, M.; Marcinkowski, K.; Kobiela, T.; Geue, T. Unusual penetration of phospholipid mono- and bilayers by Quillaja bark saponin biosurfactant. Biochim. Biophys. Acta Biomembr. 2014, 1838. [Google Scholar] [CrossRef]
- Pagureva, N.; Tcholakova, S.; Golemanov, K.; Denkov, N.; Pelan, E.; Stoyanov, S.D. Surface properties of adsorption layers formed from triterpenoid and steroid saponins. Colloids Surf. A Physicochem. Eng. Asp. 2016, 491, 18–28. [Google Scholar] [CrossRef]
- Orczyk, M.; Wojciechowski, K. Comparison of the effect of two Quillaja bark saponin extracts on DPPC and DPPC/cholesterol Langmuir monolayers. Colloids Surf. B Biointerfaces 2015, 136, 291–299. [Google Scholar] [CrossRef]
- Ransom, F. Saponin und sein Gegengift. Dtsch. Med. Wochenschr. 1901, 27, 194–196. [Google Scholar] [CrossRef][Green Version]
- Windaus, A. Über die Entgiftung der Saponine durch Cholesterin. Ber. Dtsch. Chem. Ges. 1909, 42, 238–246. [Google Scholar] [CrossRef]
- Sidhu, G.S.; Oakenfull, D.G. A mechanism for the hypocholesterolaemic activity of saponins. Br. J. Nutr. 1986, 55, 643–649. [Google Scholar] [CrossRef]
- Smith, W.S.; Baker, E.J.; Holmes, S.E.; Koster, G.; Hunt, A.N.; Johnston, D.A.; Flavell, S.U.; Flavell, D.J. Membrane cholesterol is essential for triterpenoid saponin augmentation of a saporin-based immunotoxin directed against CD19 on human lymphoma cells. Biochim. Biophys. Acta Biomembr. 2017, 1859, 993–1007. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Singh, J.P.; Singh, N.; Kaur, A. Saponins in pulses and their health promoting activities: A review. Food Chem. 2017, 233, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Korchowiec, B.; Janikowska-Sagan, M.; Kwiecińska, K.; Stachowicz-Kuśnierz, A.; Korchowiec, J. The role of cholesterol in membrane activity of digitonin: Experimental and theoretical model studies. J. Mol. Liq. 2021, 323. [Google Scholar] [CrossRef]
- Malabed, R.; Hanashima, S.; Murata, M.; Sakurai, K. Interactions of OSW-1 with Lipid Bilayers in Comparison with Digitonin and Soyasaponin. Langmuir 2020, 36, 3600–3610. [Google Scholar] [CrossRef]
- Orczyk, M.; Wojciechowski, K.; Brezesinski, G. The influence of steroidal and triterpenoid saponins on monolayer models of the outer leaflets of human erythrocytes, E. coli and S. cerevisiae cell membranes. J. Colloid Interface Sci. 2020, 563, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Geisler, R.; Dargel, C.; Hellweg, T. The biosurfactant β-aescin: A review on the physico-chemical properties and its interaction with lipid model membranes and langmuir monolayers. Molecules 2020, 25, 117. [Google Scholar] [CrossRef]
- Verstraeten, S.L.; Deleu, M.; Janikowska-Sagan, M.; Claereboudt, E.J.S.; Lins, L.; Tyteca, D.; Mingeot-Leclercq, M.-P. The activity of the saponin ginsenoside Rh2 is enhanced by the interaction with membrane sphingomyelin but depressed by cholesterol. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- De Groot, C.; Müsken, M.; Müller-Goymann, C.C. The bidesmosidic triterpene saponins hederacoside C and ginsenoside Rb1 exhibit low affinity to cholesterol in liposomal membranes. J. Drug Deliv. Sci. Technol. 2019, 53. [Google Scholar] [CrossRef]
- Goodman, J.R.; Jarnagin, L.P.; Meier, R.M.; Shonley, I.A. Determination of Free and Esterified Cholesterol by a Modified Digitonin-Anthrone Method. Anal. Chem. 1963, 35, 760–763. [Google Scholar] [CrossRef]
- Nishikawa, M.; Nojima, S.; Akiyama, T.; Sankawa, U.; Inoue, K. Interaction of digitonin and its analogs with membrane cholesterol. J. Biochem. 1984, 96, 1231–1239. [Google Scholar] [CrossRef]
- Holmes, S.E.; Bachran, C.; Fuchs, H.; Weng, A.; Melzig, M.F.; Flavell, S.U.; Flavell, D.J. Triterpenoid saponin augmention of saporin-based immunotoxin cytotoxicity for human leukaemia and lymphoma cells is partially immunospecific and target molecule dependent. Immunopharmacol. Immunotoxicol. 2015, 37, 42–55. [Google Scholar] [CrossRef]
- Tatia, R.; Zalaru, C.; Tarcomnicu, I.; Moldovan, L.; Craciunescu, O.; Calinescu, I. Isolation and characterization of hederagenin from hedera helix L. Extract with antitumor activity. Rev. Chim. 2019, 70, 1157–1161. [Google Scholar] [CrossRef]
- Jaramillo-Carmona, S.; Guillén-Bejarano, R.; Jiménez-Araujo, A.; Rodríguez-Arcos, R.; López, S. In Vitro Toxicity of Asparagus Saponins in Distinct Multidrug-Resistant Colon Cancer Cells. Chem. Biodivers. 2018, 15. [Google Scholar] [CrossRef] [PubMed]
- Dutt, R.; Garg, V.; Khatri, N.; Madan, A.K. Phytochemicals in anticancer drug development. Anticancer Agents Med. Chem. 2019, 19, 172–183. [Google Scholar] [CrossRef]
- Dahibhate, N.L.; Saddhe, A.A.; Kumar, K. Mangrove plants as a source of bioactive compounds: A review. Nat. Prod. J. 2019, 9, 86–97. [Google Scholar] [CrossRef]
- Xu, J.; Wang, Z.; Huang, Y.; Wang, Y.; Xiang, L.; He, X. A spirostanol saponin isolated from Tupistra chinensis Baker simultaneously induces apoptosis and autophagy by regulating the JNK pathway in human gastric cancer cells. Steroids 2020, 164. [Google Scholar] [CrossRef]
- Garrido, C.M.; Henkels, K.M.; Rehl, K.M.; Liang, H.; Zhou, Y.; Gutterman, J.U.; Cho, K.-J. Avicin G is a potent sphingomyelinase inhibitor and blocks oncogenic K- and H-Ras signaling. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Barr, A.M.; Silva, A.; Prato, S.; Belz, G.T.; Maraskovsky, E.; Baz Morelli, A. Therapeutic ISCOMATRIXTM adjuvant vaccine elicits effective anti-tumor immunity in the TRAMP-C1 mouse model of prostate cancer. Cancer Immunol. Immunother. 2020, 69, 1959–1972. [Google Scholar] [CrossRef]
- Mbaveng, A.T.; Chi, G.F.; Nguenang, G.S.; Abdelfatah, S.; Tchangna Sop, R.V.; Ngadjui, B.T.; Kuete, V.; Efferth, T. Cytotoxicity of a naturally occuring spirostanol saponin, progenin III, towards a broad range of cancer cell lines by induction of apoptosis, autophagy and necroptosis. Chem. Biol. Interact. 2020, 326. [Google Scholar] [CrossRef]
- Magedans, Y.V.S.; Yendo, A.C.A.; Costa, F.D.; Gosmann, G.; Fett-Neto, A.G. Foamy matters: An update on Quillaja saponins and their use as immunoadjuvants. Future Med. Chem. 2019, 11, 1485–1499. [Google Scholar] [CrossRef] [PubMed]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as cytotoxic agents: A review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef]
- Roner, M.R.; Sprayberry, J.; Spinks, M.; Dhanji, S. Antiviral activity obtained from aqueous extracts of the Chilean soapbark tree (Quillaja saponaria Molina). J. Gen. Virol. 2007, 88, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Roner, M.R.; Tam, K.I.; Kiesling-Barrager, M. Prevention of rotavirus infections in vitro with aqueous extracts of Quillaja Saponaria Molina. Future Med. Chem. 2010, 2, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- Arabski, M.; Wȩgierek-Ciuk, A.; Czerwonka, G.; Lankoff, A.; Kaca, W. Effects of saponins against clinical E. coli strains and eukaryotic cell line. J. Biomed. Biotechnol. 2012, 2012. [Google Scholar] [CrossRef]
- Van De Ven, H.; Vermeersch, M.; Shunmugaperumal, T.; Vandervoort, J.; Maes, L.; Ludwig, A. Solid lipid nanoparticle (SLN) formulations as a potential tool for the reduction of cytotoxicity of saponins. Pharmazie 2009, 64, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lu, H. Microfluidics in systems biology-hype or truly useful? Curr. Opin. Biotechnol. 2016, 39, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Tokarska, K.; Bazylinska, U.; Jastrzebska, E.; Chudy, M.; Dybko, A.; Wilk, K.A.; Brzozka, Z. Selective cancer-killing ability of new efficient porphyrin-based nanophotosensitizer in Lab-on-a-chip system. Sens. Actuators B Chem. 2019, 282, 665–674. [Google Scholar] [CrossRef]
- Pandey, C.M.; Augustine, S.; Kumar, S.; Kumar, S.; Nara, S.; Srivastava, S.; Malhotra, B.D. Microfluidics Based Point-of-Care Diagnostics. Biotechnol. J. 2018, 13. [Google Scholar] [CrossRef]
- Bruijns, B.; van Asten, A.; Tiggelaar, R.; Gardeniers, H. Microfluidic devices for forensic DNA analysis: A review. Biosensors 2016, 6, 41. [Google Scholar] [CrossRef]
- Monošík, R.; Angnes, L. Utilisation of micro- and nanoscaled materials in microfluidic analytical devices. Microchem. J. 2015, 119, 159–168. [Google Scholar] [CrossRef]
- Sousa, M.P.; Arab-Tehrany, E.; Cleymand, F.; Mano, J.F. Surface Micro- and Nanoengineering: Applications of Layer-by-Layer Technology as a Versatile Tool to Control Cellular Behavior. Small 2019, 15. [Google Scholar] [CrossRef]
- Lee, S.; Kim, H.; Lee, W.; Kim, J. Microfluidic-based cell handling devices for biochemical applications. J. Micromech. Microeng. 2018, 28. [Google Scholar] [CrossRef]
- Hu, C.; Chen, Y.; Tan, M.J.A.; Ren, K.; Wu, H. Microfluidic technologies for vasculature biomimicry. Analyst 2019, 144, 4461–4471. [Google Scholar] [CrossRef]
- Wang, Z.; Kim, M.-C.; Marquez, M.; Thorsen, T. High-density microfluidic arrays for cell cytotoxicity analysis. Lab Chip 2007, 7, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Ziolkowska, K.; Jedrych, E.; Kwapiszewski, R.; Lopacinska, J.; Skolimowski, M.; Chudy, M. PDMS/glass microfluidic cell culture system for cytotoxicity tests and cells passage. Sens. Actuators B Chem. 2010, 145, 533–542. [Google Scholar] [CrossRef]
- Wojciechowski, K.; Orczyk, M.; Gutberlet, T.; Geue, T. Complexation of phospholipids and cholesterol by triterpenic saponins in bulk and in monolayers. Biochim. Biophys. Acta Biomembr. 2016, 1858, 363–373. [Google Scholar] [CrossRef]
- Korchowiec, B.; Gorczyca, M.; Wojszko, K.; Janikowska, M.; Henry, M.; Rogalska, E. Impact of two different saponins on the organization of model lipid membranes. Biochim. Biophys. Acta Biomembr. 2015, 1848, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- Glauert, A.M.; Dingle, J.T.; Lucy, J.A.; Bangham, A.D.; Horne, R.W. Action of Saponin on Biological Cell Membranes. Nature 1962, 196, 952–953. [Google Scholar] [CrossRef]
- Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochemistry 2011, 72, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Lorent, J.; Le Duff, C.S.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.-P. Induction of highly curved structures in relation to membrane permeabilization and budding by the triterpenoid saponins, α- And δ-hederin. J. Biol. Chem. 2013, 288, 14000–14017. [Google Scholar] [CrossRef]
- Armah, C.N.; Mackie, A.R.; Roy, C.; Price, K.; Osbourn, A.E.; Bowyer, P.; Ladha, S. The membrane-permeabilizing effect of avenacin A-1 involves the reorganization of bilayer cholesterol. Biophys. J. 1999, 76, 281–290. [Google Scholar] [CrossRef]
- Keukens, E.A.J.; De Vrije, T.; Jansen, L.A.M.; De Boer, H.; Janssen, M.; De Kroon, A.I.P.M.; Jongen, W.M.F.; De Kruijff, B. Glycoalkaloids selectively permeabilize cholesterol containing biomembranes. Biochim. Biophys. Acta Biomembr. 1996, 1279, 243–250. [Google Scholar] [CrossRef]
- Giard, D.J.; Aaronson, S.A.; Todaro, G.J.; Arnstein, P.; Kersey, J.H.; Dosik, H.; Parks, W.P. In vitro cultivation of human tumors: Establishment of cell lines derived from a series of solid tumors. J. Natl. Cancer Inst. 1973, 51, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Maier, C.; Conrad, J.; Carle, R.; Weiss, J.; Schweiggert, R.M. Phenolic constituents in commercial aqueous quillaja (Quillaja saponaria Molina) Wood extracts. J. Agric. Food Chem. 2015, 63, 1756–1762. [Google Scholar] [CrossRef] [PubMed]
- Tarahovsky, Y.S. Plant polyphenols in cell-cell interaction and communication. Plant Signal. Behav. 2008, 3, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Grootaert, C.; Kamiloglu, S.; Capanoglu, E.; Van Camp, J. Cell systems to investigate the impact of polyphenols on cardiovascular health. Nutrients 2015, 7, 9229–9255. [Google Scholar] [CrossRef] [PubMed]
- Chwalek, M.; Lalun, N.; Bobichon, H.; Plé, K.; Voutquenne-Nazabadioko, L. Structure–activity relationships of some hederagenin diglycosides: Haemolysis, cytotoxicity and apoptosis induction. Biochim. Biophys. Acta Gen. Subj. 2006, 1760, 1418–1427. [Google Scholar] [CrossRef]
- Gevrenova, R.; Weng, A.; Voutguenne-Nazabadioko, L.; Thakur, M.; Doytchinova, I. Quantitative Structure–Activity Relationship Study on Saponins as Cytotoxicity Enhancers. Lett. Drug Des. Discov. 2014, 12, 166–171. [Google Scholar] [CrossRef]
- De Groot, C.; Müller-Goymann, C.C. Saponin Interactions with Model Membrane Systems-Langmuir Monolayer Studies, Hemolysis and Formation of ISCOMs. Planta Med. 2016, 82, 1496–1512. [Google Scholar] [CrossRef]
- Jaiare, N.; Ithara, A.; Kumapav, K. Cytotoxic saponin against lung cancer cells from Dioscorea birmanica Prain & Burkill. J. Med. Assoc. Thail. 2010, 93, 192–197. [Google Scholar]
- Trapani, L.; Segatto, M.; Pallottini, V. Regulation and deregulation of cholesterol homeostasis: The liver as a metabolic “power station”. World J. Hepatol. 2012, 4, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Inbar, M.; Goldman, R.; Inbar, L.; Bursuker, I.; Goldman, B.; Akstein, E.; Segal, P.; Ipp, E.; Ben-Bassat, I. Fluidity difference of membrane lipids in human normal and leukemic lymphocytes as controlled by serum components. Cancer Res. 1977, 37, 3037–3041. [Google Scholar]
- Kuzu, O.F.; Noory, M.A.; Robertson, G.P. The role of cholesterol in cancer. Cancer Res. 2016, 76, 2063–2070. [Google Scholar] [CrossRef]
- Krycer, J.R.; Brown, A.J. Cholesterol accumulation in prostate cancer: A classic observation from a modern perspective. Biochim. Biophys. Acta Rev. Cancer 2013, 1835, 219–229. [Google Scholar] [CrossRef]
- Inbar, M.; Shinitzky, M. Cholesterol as a bioregulator in the development and inhibition of leukemia. Proc. Natl. Acad. Sci. USA 1974, 71, 4229–4231. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozińska, N.; Tokarska, K.; Chudy, M.; Wojciechowski, K. Cytotoxicity of Quillaja saponaria Saponins towards Lung Cells Is Higher for Cholesterol-Rich Cells. Biophysica 2021, 1, 126-136. https://doi.org/10.3390/biophysica1020010
Kozińska N, Tokarska K, Chudy M, Wojciechowski K. Cytotoxicity of Quillaja saponaria Saponins towards Lung Cells Is Higher for Cholesterol-Rich Cells. Biophysica. 2021; 1(2):126-136. https://doi.org/10.3390/biophysica1020010
Chicago/Turabian StyleKozińska, Natalia, Katarzyna Tokarska, Michał Chudy, and Kamil Wojciechowski. 2021. "Cytotoxicity of Quillaja saponaria Saponins towards Lung Cells Is Higher for Cholesterol-Rich Cells" Biophysica 1, no. 2: 126-136. https://doi.org/10.3390/biophysica1020010
APA StyleKozińska, N., Tokarska, K., Chudy, M., & Wojciechowski, K. (2021). Cytotoxicity of Quillaja saponaria Saponins towards Lung Cells Is Higher for Cholesterol-Rich Cells. Biophysica, 1(2), 126-136. https://doi.org/10.3390/biophysica1020010