
Systemic Tumors Can Cause Molecular Changes in the Hippocampus That May Have an Impact on Behavior after Chronic Social Stress

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
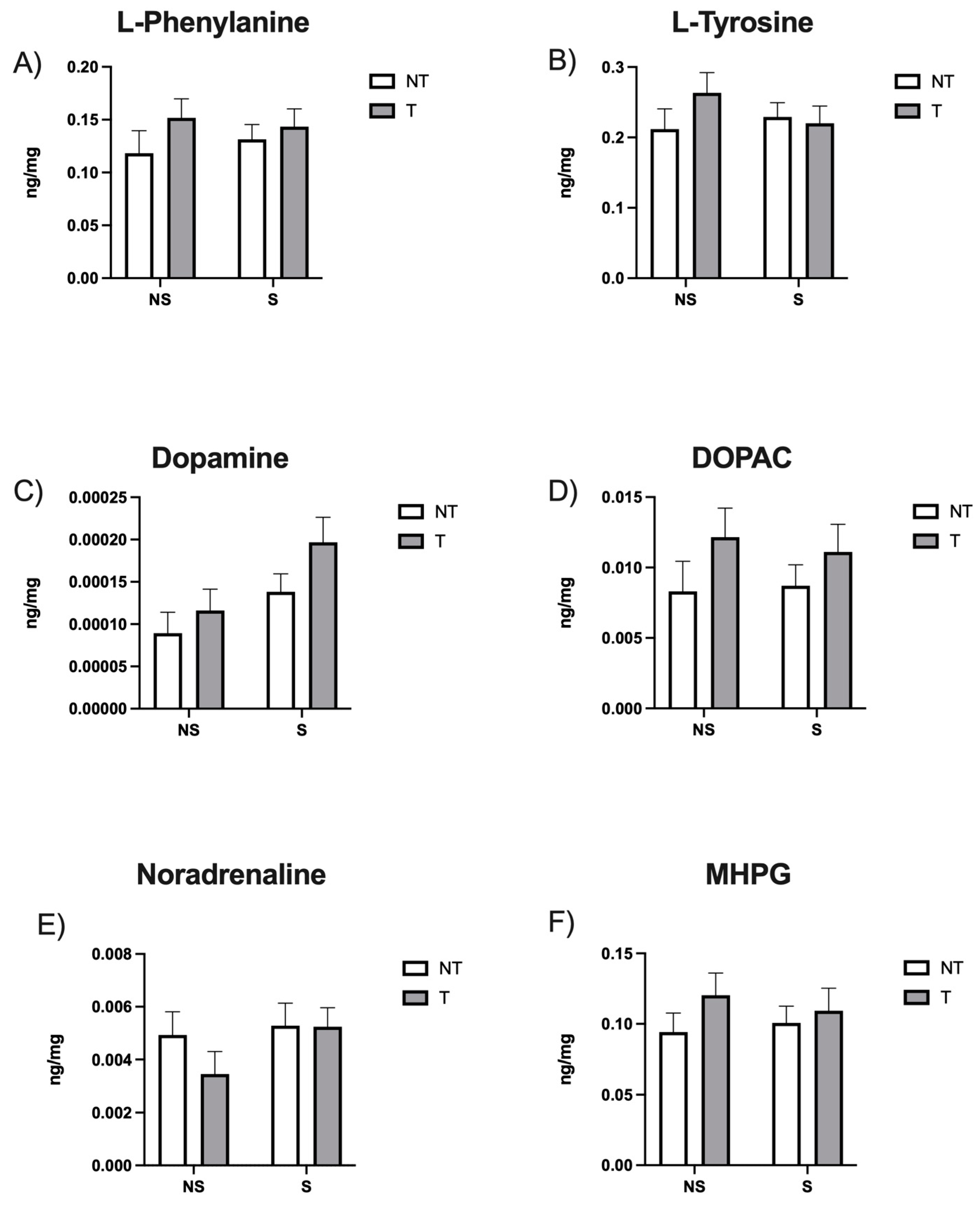
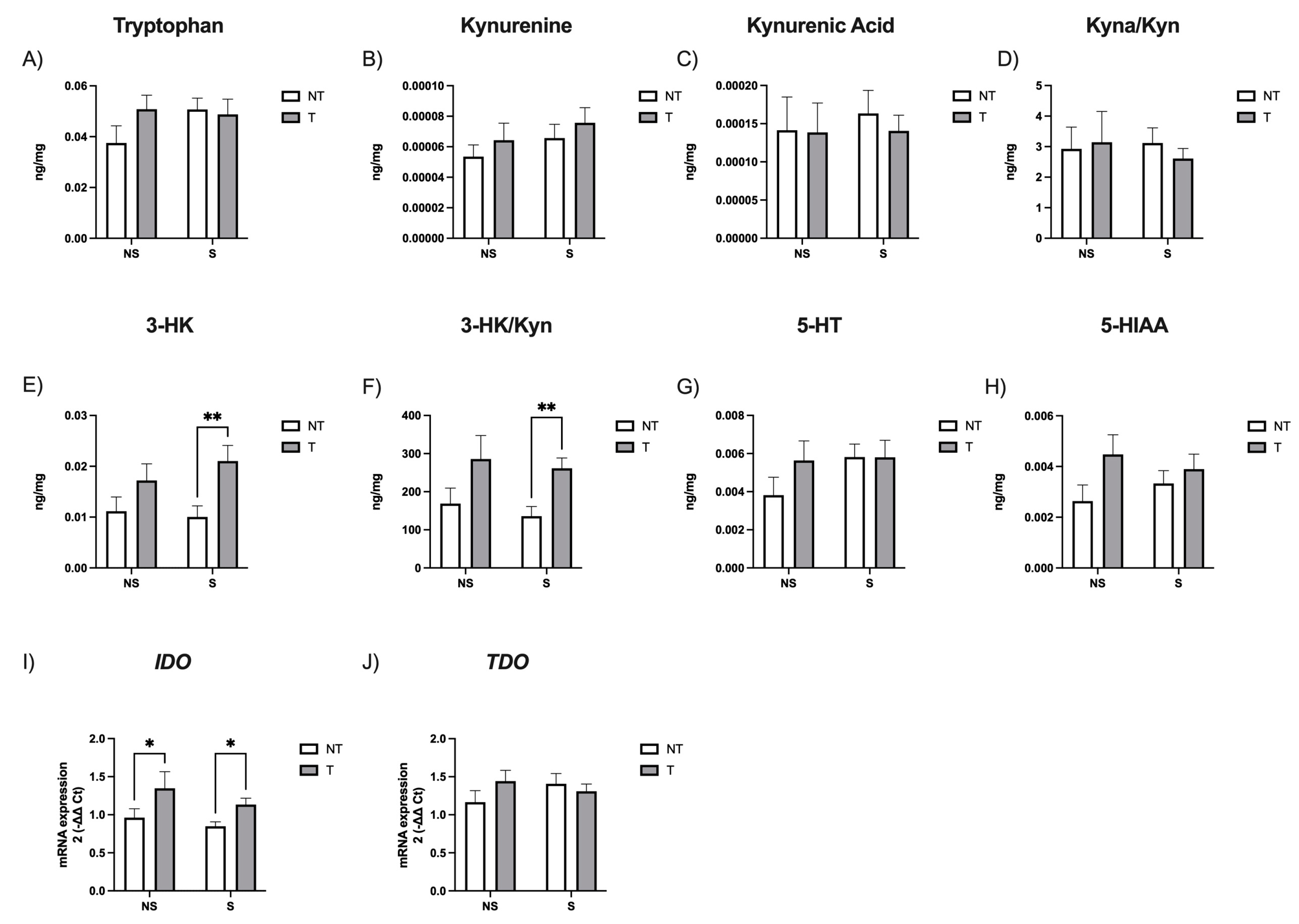
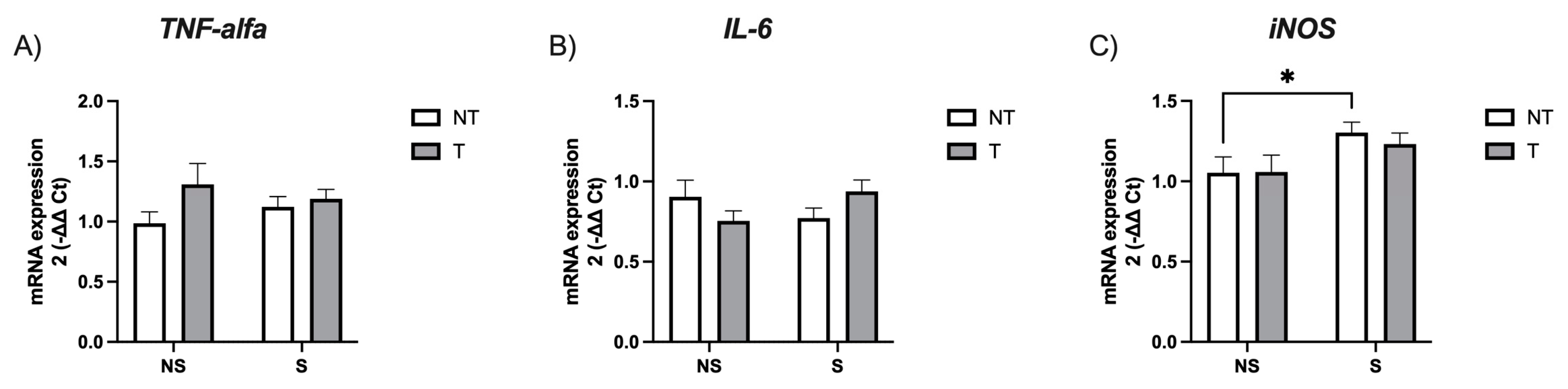
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Cohen, S.; Wills, T.A. Stress, social support, and the buffering hypothesis. Psychol. Bull. 1985, 98, 310–357. [Google Scholar] [CrossRef] [PubMed]
- Godoy, L.D.; Rossignoli, M.T.; Delfino-Pereira, P.; Garcia-Cairasco, N.; de Lima Umeoka, E.H. A Comprehensive Overview on Stress Neurobiology: Basic Concepts and Clinical Implications. Front. Behav. Neurosci. 2018, 12, 127. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.R.; Cole, S.W. Reciprocal regulation of the neural and innate immune systems. Nat. Rev. Immunol. 2011, 11, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Hollis, F.; Isgor, C.; Kabbaj, M. The consequences of adolescent chronic unpredictable stress exposure on brain and behavior. Neuroscience 2013, 249, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Gotlib, I.; Hammen, C. Handbook of Depression; Guilford Press: New York, NY, USA, 2008; p. 708. [Google Scholar]
- Sommershof, A.; Scheuermann, L.; Koerner, J.; Groettrup, M. Chronic stress suppresses anti-tumor T. Brain Behav. Immun. 2017, 65, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Chida, Y.; Hamer, M.; Wardle, J.; Steptoe, A. Do stress-related psychosocial factors contribute to cancer incidence and survival? Nat. Clin. Pract. Oncol. 2008, 5, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Smith, M.; Lutgendorf, S.K.; Sood, A.K. Impact of stress on sleep. Future Oncol. 2010, 6, 1863–1881. [Google Scholar] [CrossRef] [PubMed]
- Lutgendorf, S.K.; Andersen, B.L. Biobehavioral approaches to cancer progression and survival: Mechanisms and interventions. Am. Psychol. 2015, 70, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, D.; Giese-Davis, J. Depression and cancer: Mechanisms and disease progression. Biol. Psychiatry 2003, 54, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, G.; Graca, J.; Klut, C.; Trancas, B.; Papoila, A. Depression and anxiety symptoms following cancer diagnosis: A cross-sectional study. Psychol. Health Med. 2016, 21, 562–570. [Google Scholar] [CrossRef]
- Satin, J.R.; Linden, W.; Phillips, M.J. Depression as a predictor of disease progression and mortality in cancer patients: A meta-analysis. Cancer 2009, 115, 5349–5361. [Google Scholar] [CrossRef]
- Antoni, M.H.; Dhabhar, F.S. The impact of psychosocial stress and stress management on immune responses in patients with cancer. Cancer 2019, 125, 1417–1431. [Google Scholar] [CrossRef]
- Dantzer, R. Role of the Kynurenine Metabolism Pathway in Inflammation-Induced Depression: Preclinical Approaches. Curr. Top. Behav. Neurosci. 2017, 31, 117–138. [Google Scholar] [CrossRef]
- Soung, N.K.; Kim, B.Y. Psychological stress and cancer. J. Anal. Sci. Technol. 2015, 6, 4–9. [Google Scholar] [CrossRef]
- Santos, J.C.; Pyter, L.M. Neuroimmunology of behavioral comorbidities associated with cancer and cancer treatments. Front. Immunol. 2018, 9, 1195. [Google Scholar] [CrossRef]
- Gómez-Lázaro, E.; Garmendia, L.; Beitia, G.; Perez-Tejada, J.; Azpiroz, A.; Arregi, A. Effects of a putative antidepressant with a rapid onset of action in defeated mice with different coping strategies. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 38, 317–327. [Google Scholar] [CrossRef]
- Pérez-Tejada, J.; Arregi, A.; Gómez-Lázaro, E.; Vegas, O.; Azpiroz, A.; Garmendia, L. Coping with chronic social stress in mice: Hypothalamic-pituitary-adrenal/sympathetic-adrenal-medullary axis activity, behavioral changes and effects of antalarmin treatment: Implications for the study of stress-related psychopathologies. Neuroendocrinology 2013, 98, 73–88. [Google Scholar] [CrossRef]
- Francis, T.C.; Chandra, R.; Friend, D.M.; Finkel, E.; Dayrit, G.; Miranda, J.; Brooks, J.M.; Iñiguez, S.D.; O’Donnell, P.; Kravitz, A.; et al. Nucleus accumbens medium spiny neuron subtypes mediate depression-related outcomes to social defeat stress. Biol. Psychiatry 2015, 77, 212–222. [Google Scholar] [CrossRef]
- Friedman, A.K.; Walsh, J.J.; Juarez, B.; Ku, S.M.; Chaudhury, D.; Wang, J.; Li, X.; Dietz, D.M.; Pan, N.; Vialou, V.F.; et al. Enhancing depression mechanisms in midbrain dopamine neurons achieves homeostatic resilience. Science 2014, 344, 313–319. [Google Scholar] [CrossRef]
- Hodes, G.E.; Pfau, M.L.; Leboeuf, M.; Golden, S.A.; Christoffel, D.J.; Bregman, D.; Rebusi, N.; Heshmati, M.; Aleyasin, H.; Warren, B.L.; et al. Individual differences in the peripheral immune system promote resilience versus susceptibility to social stress. Proc. Natl. Acad. Sci. USA 2014, 111, 16136–16141. [Google Scholar] [CrossRef]
- Isingrini, E.; Perret, L.; Rainer, Q.; Amilhon, B.; Guma, E.; Tanti, A.; Martin, G.; Robinson, J.; Moquin, L.; Marti, F.; et al. Resilience to chronic stress is mediated by noradrenergic regulation of dopamine neurons. Nat. Neurosci. 2016, 19, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Avitsur, R.; Powell, N.; Padgett, D.A.; Sheridan, J.F. Social Interactions, stress, and immunity. Immunol. Allergy Clin. N. Am. 2009, 29, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Ballestin, R.; Alegre-Zurano, L.; Ferrer-Perez, C.; Cantacorps, L.; Miarro, J.; Valverde, O.; Rodriguez-Arias, M. Neuroinflammatory and behavioral susceptibility profile of mice exposed to social stress towards cocaine effects. Progress. Neuropsychopharmacol. Biol. Psychiatry 2021, 105, 110123. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Tejada, J.; Arregi, A.; Azpiroz, A.; Beitia, G.; Gómez-Lázaro, E.; Garmedia, L. Central immune alterations in passive strategy following chronic defeat stress. Behav. Brain Res. 2016, 298, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Lázaro, E.; Arregi, A.; Beitia, G.; Vegas, O.; Azpiroz, A.; Garmendia, L. Individual differences in chronically defeated male mice: Behavioral, endocrine, immune, and neurotrophic changes as markers of vulnerability to the effects of stress. Stress 2011, 14, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Reguilón, M.D.; Ballestín, R.; Miñarro, J.; Rodríguez-Arias, M. Resilience to social defeat stress in adolescent male mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 119, 110591. [Google Scholar] [CrossRef] [PubMed]
- Ródenas-González, F.; Blanco-Gandía, M.D.C.; Miñarro López, J.; Rodriguez-Arias, M. Behavioral and neuroimmune characterization of resilience to social stress: Rewarding effects of cocaine. Adicciones 2021, 33, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Liu, H.; Tan, P.; Hu, Z.; Ma, Y.; Ye, M.; Gu, Y.; Wang, Y.; Ye, T.; Lu, X.; et al. Innate immune stimulation prevents the development of anxiety-like behaviors in chronically stressed mice. Neuropharmacology 2022, 207, 108950. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Xiang, H.; Gu, Y.; Ye, T.; Lu, X.; Huang, C. Innate immune stimulation by monophosphoryl lipid A prevents chronic social defeat stress-induced anxiety-like behaviors in mice. J. Neuroinflammation 2022, 19, 12. [Google Scholar] [CrossRef]
- Hersey, M.; Hashemi, P.; Reagan, L.P. Integrating the monoamine and cytokine hypotheses of depression: Is histamine the missing link? Eur. J. Neurosci. 2022, 55, 2895–2911. [Google Scholar] [CrossRef]
- Felger, J.C.; Lotrich, F.E. Inflammatory cytokines in depression: Neurobiological mechanisms and therapeutic implications. Neuroscience 2013, 246, 199–229. [Google Scholar] [CrossRef] [PubMed]
- Delgado, P.L. Depression: The case for a monoamine deficiency. J. Clin. Psychiatry 2000, 61 (Suppl. S6), 7–11. [Google Scholar] [PubMed]
- Eisenhofer, G.; Kopin, I.J.; Goldstein, D.S. Catecholamine metabolism: A contemporary view with implications for physiology and medicine. Pharmacol. Rev. 2004, 56, 331–349. [Google Scholar] [CrossRef] [PubMed]
- Höglund, E.; Øverli, Ø.; Winberg, S. Tryptophan Metabolic Pathways and Brain Serotonergic Activity: A Comparative Review. Front. Endocrinol. 2019, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- O’Farrell, K.; Harkin, A. Stress-related regulation of the kynurenine pathway: Relevance to neuropsychiatric and degenerative disorders. Neuropharmacology 2017, 112 Pt B, 307–323. [Google Scholar] [CrossRef]
- Lebeña, A.; Vegas, O.; Gómez-Lázaro, E.; Arregi, A.; Garmendia, L.; Beitia, G.; Azpiroz, A. Melanoma tumors alter proinflammatory cytokine production and monoamine brain function, and induce depressive-like behavior in male mice. Behav. Brain Res. 2014, 272, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Goñi-Balentziaga, O.; Garmendia, L.; Labaka, A.; Lebeña, A.; Beitia, G.; Gómez-Lázaro, E.; Vegas, O. Behavioral coping strategies predict tumor development and behavioral impairment after chronic social stress in mice. Physiol. Behav. 2020, 214, 112747. [Google Scholar] [CrossRef] [PubMed]
- Díez-Solinska, A.; Azkona, G.; Muñoz-Culla, M.; Beitia-Oyarzabal, G.; Goñi-Balentziaga, O.; Gómez-Lazaro, E.; Vegas, O. The role of sociability in social instability stress: Behavioral, neuroendocrine and monoaminergic effects. Physiol. Behav. 2023, 270, 114306. [Google Scholar] [CrossRef] [PubMed]
- Díez-Solinska, A.; Lebeña, A.; Garmendia, L.; Labaka, A.; Azkona, G.; Perez-Tejada, J.; Vegas, O. Chronic social instability stress down-regulates IL-10 and up-regulates CX3CR1 in tumor-bearing and non-tumor-bearing female mice. Behav. Brain Res. 2022, 435, 114063. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Fuertig, R.; Azzinnari, D.; Bergamini, G.; Cathomas, F.; Sigrist, H.; Seifritz, E.; Vavassori, S.; Luippold, A.; Hengerer, B.; Ceci, A.; et al. Mouse chronic social stress increases blood and brain kynurenine pathway activity and fear behaviour: Both effects are reversed by inhibition of indoleamine 2,3-dioxygenase. Brain Behav. Immun. 2016, 54, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Favoretto, C.A.; Nunes, Y.C.; Macedo, G.C.; Lopes, J.S.R.; Quadros, I.M.H. Chronic social defeat stress: Impacts on ethanol-induced stimulation, corticosterone response, and brain monoamine levels. J. Psychopharmacol. 2020, 34, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Hestad, K.; Alexander, J.; Rootwelt, H.; Aaseth, J.O. The Role of Tryptophan Dysmetabolism and Quinolinic Acid in Depressive and Neurodegenerative Diseases. Biomolecules 2022, 12, 998. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K. Etiological classification of depression based on the enzymes of tryptophan metabolism. BMC Psychiatry 2014, 14, 372. [Google Scholar] [CrossRef] [PubMed]
- Meireson, A.; Devos, M.; Brochez, L. IDO Expression in Cancer: Different Compartment, Different Functionality? Front. Immunol. 2020, 11, 531491. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Toups, M.; Nemeroff, C.B. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef] [PubMed]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef]
- Beheshti, F.; Hashemzehi, M.; Hosseini, M.; Marefati, N.; Memarpour, S. Inducible nitric oxide synthase plays a role in depression- and anxiety-like behaviors chronically induced by lipopolysaccharide in rats: Evidence from inflammation and oxidative stress. Behav. Brain Res. 2020, 392, 112720. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goñi-Balentziaga, O.; Díez-Solinska, A.; Beitia-Oyarzabal, G.; Muñoz-Culla, M.; Azkona, G.; Vegas, O. Systemic Tumors Can Cause Molecular Changes in the Hippocampus That May Have an Impact on Behavior after Chronic Social Stress. NeuroSci 2024, 5, 192-200. https://doi.org/10.3390/neurosci5020014
Goñi-Balentziaga O, Díez-Solinska A, Beitia-Oyarzabal G, Muñoz-Culla M, Azkona G, Vegas O. Systemic Tumors Can Cause Molecular Changes in the Hippocampus That May Have an Impact on Behavior after Chronic Social Stress. NeuroSci. 2024; 5(2):192-200. https://doi.org/10.3390/neurosci5020014
Chicago/Turabian StyleGoñi-Balentziaga, Olatz, Alina Díez-Solinska, Garikoitz Beitia-Oyarzabal, Maider Muñoz-Culla, Garikoitz Azkona, and Oscar Vegas. 2024. "Systemic Tumors Can Cause Molecular Changes in the Hippocampus That May Have an Impact on Behavior after Chronic Social Stress" NeuroSci 5, no. 2: 192-200. https://doi.org/10.3390/neurosci5020014
APA StyleGoñi-Balentziaga, O., Díez-Solinska, A., Beitia-Oyarzabal, G., Muñoz-Culla, M., Azkona, G., & Vegas, O. (2024). Systemic Tumors Can Cause Molecular Changes in the Hippocampus That May Have an Impact on Behavior after Chronic Social Stress. NeuroSci, 5(2), 192-200. https://doi.org/10.3390/neurosci5020014