
Olfactory Dysfunction in Parkinson’s Disease, Its Functional and Neuroanatomical Correlates

 , ,
, ,  , , ,
, , ,  and
and 
{kind=link}
Abstract
1. Introduction
2. Parkinson’s Disease
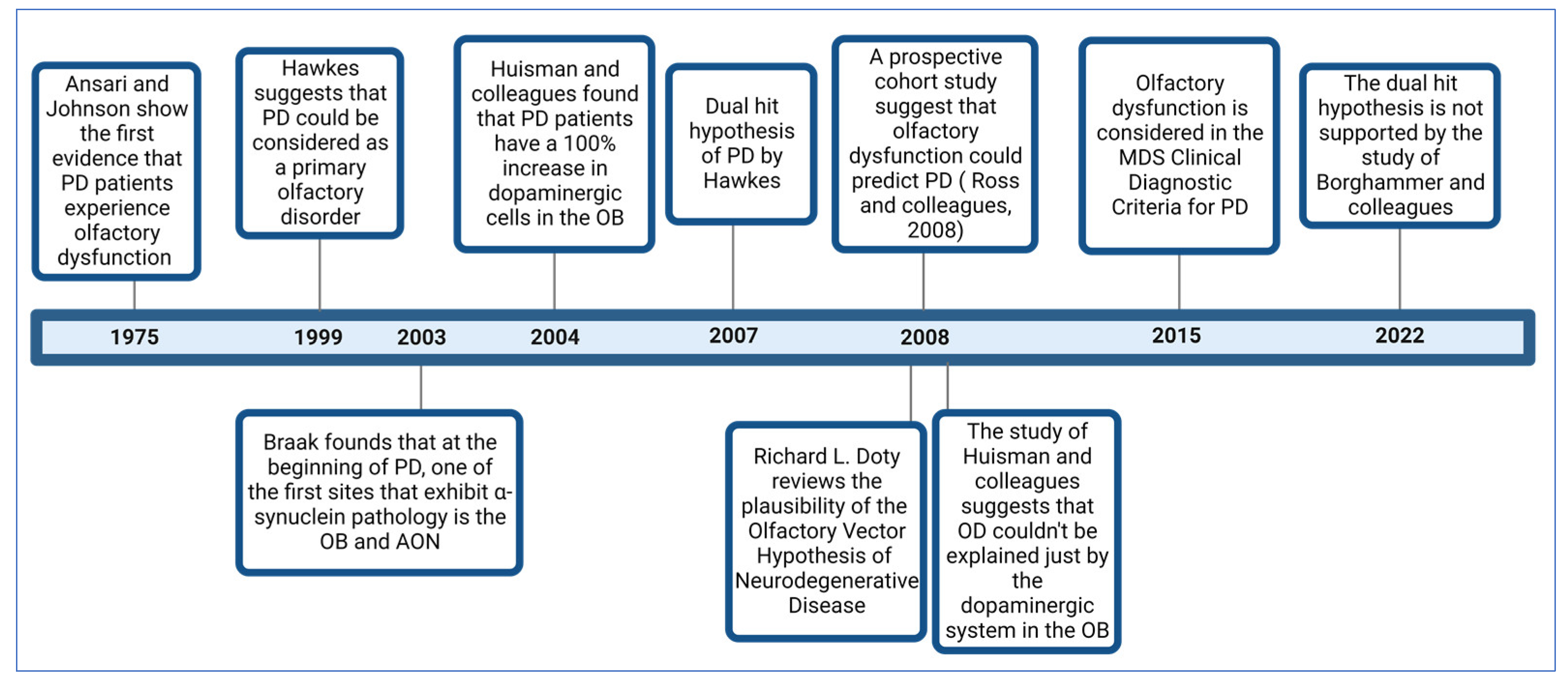
3. A Brief Review of the Evolution of Knowledge about Olfactory Dysfunction in Parkinson’s Disease
4. Prevalence of Olfactory Dysfunction and Its Role in Diagnosis and as a Marker of PD Progression
5. Assessment of the Olfactory System in Patients with PD
6. The Role of α-Synuclein and Its Aggregates in the Pathophysiology of PD
7. Neuroanatomical Alterations in PD Patients
7.1. Olfactory Epithelium
7.2. Olfactory Bulb
7.3. Anterior Olfactory Nucleus
7.4. Olfactory Tract
7.5. Piriform Cortex
7.6. Hippocampus
7.7. Orbitofrontal Cortex
7.8. Amygdala
7.9. Cerebellum
8. Discussion
9. Future Directions
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erkkinen, M.G.; Kim, M.-O.; Geschwind, M.D. Clinical Neurology and Epidemiology of the Major Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2018, 10, a033118. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, E.; Gaig, C.; Santamaría, J.; Compta, Y. Diagnosis and the premotor phase of Parkinson disease. Neurology 2009, 72, S12–S20. [Google Scholar] [CrossRef]
- Stevenson, T.J.; Murray, H.C.; Turner, C.; Faull, R.L.M.; Dieriks, B.V.; Curtis, M.A. α-synuclein inclusions are abundant in non-neuronal cells in the anterior olfactory nucleus of the Parkinson’s disease olfactory bulb. Sci. Rep. 2020, 10, 6682. [Google Scholar] [CrossRef]
- Ross, G.W.; Petrovitch, H.; Abbott, R.D.; Tanner, C.M.; Popper, J.; Masaki, K.; Launer, L.; White, L.R. Association of olfactory dysfunction with risk for future Parkinson’s disease. Ann. Neurol. 2008, 63, 167–173. [Google Scholar] [CrossRef]
- Blomqvist, E.H.; Brämerson, A.; Stjärne, P.; Nordin, S. Consequences of olfactory loss and adopted coping strategies. Rhinol. J. 2004, 42, 189–194. Available online: https://www.rhinologyjournal.com/Abstract.php?id=455 (accessed on 23 April 2023).
- White, T.L.; Sadikot, A.F.; Djordjevic, J. Metacognitive knowledge of olfactory dysfunction in Parkinson’s disease. Brain Cogn. 2016, 104, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ball, N.; Teo, W.-P.; Chandra, S.; Chapman, J. Parkinson’s Disease and the Environment. Front. Neurol. 2019, 10, 218. [Google Scholar] [CrossRef]
- Tysnes, O.-B.; Storstein, A. Epidemiology of Parkinson’s disease. J. Neural Transm. 2017, 124, 901–905. [Google Scholar] [CrossRef]
- Doty, R.L. The olfactory vector hypothesis of neurodegenerative disease: Is it viable? Ann. Neurol. 2008, 63, 7–15. [Google Scholar] [CrossRef]
- Pezzoli, G.; Cereda, E. Exposure to pesticides or solvents and risk of Parkinson disease. Neurology 2013, 80, 2035–2041. [Google Scholar] [CrossRef]
- Siderowf, A.; Stern, M.B. Preclinical diagnosis of parkinson’s disease: Are we there yet? Curr. Neurol. Neurosci. Rep. 2006, 6, 295–301. [Google Scholar] [CrossRef]
- Williams, D.R.; Litvan, I. Parkinsonian Syndromes. Contin. Lifelong Learn. Neurol. 2013, 19, 1189–1212. [Google Scholar] [CrossRef]
- Pfeiffer, R.F. Non-motor symptoms in Parkinson’s disease. Park. Relat. Disord. 2015, 22 (Suppl. S1), S119–S122. [Google Scholar] [CrossRef] [PubMed]
- Masala, C.; Solla, P.; Liscia, A.; Defazio, G.; Saba, L.; Cannas, A.; Cavazzana, A.; Hummel, T.; Haehner, A. Correlation among olfactory function, motors’ symptoms, cognitive impairment, apathy, and fatigue in patients with Parkinson’s disease. J. Neurol. 2018, 265, 1764–1771. [Google Scholar] [CrossRef] [PubMed]
- Haehner, A.; Boesveldt, S.; Berendse, H.; Mackay-Sim, A.; Fleischmann, J.; Silburn, P.; Johnston, A.; Mellick, G.; Herting, B.; Reichmann, H.; et al. Prevalence of smell loss in Parkinson’s disease—A multicenter study. Park. Relat. Disord. 2009, 15, 490–494. [Google Scholar] [CrossRef]
- Berendse, H.W.; Roos, D.; Raijmakers, P.; Doty, R.L. Motor and non-motor correlates of olfactory dysfunction in Parkinson’s disease. J. Neurol. Sci. 2011, 310, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Domellöf, M.E.; Lundin, K.-F.; Edström, M.; Forsgren, L. Olfactory dysfunction and dementia in newly diagnosed patients with Parkinson’s disease. Park. Relat. Disord. 2017, 38, 41–47. [Google Scholar] [CrossRef]
- Ansari, K.; Johnson, A. Olfactory function in patients with Parkinson’s disease. J. Chronic Dis. 1975, 28, 493–497. [Google Scholar] [CrossRef]
- Hawkes, C.; Shephard, B.; Daniel, S. Is Parkinson’s disease a primary olfactory disorder? QJM 1999, 92, 473–480. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.; Steur, E.N.J.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Huisman, E.; Uylings, H.B.; Hoogland, P.V. A 100% increase of dopaminergic cells in the olfactory bulb may explain hyposmia in Parkinson’s disease. Mov. Disord. 2004, 19, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Huisman, E.; Uylings, H.B.; Hoogland, P.V. Gender-related changes in increase of dopaminergic neurons in the olfactory bulb of Parkinson’s disease patients. Mov. Disord. 2008, 23, 1407–1413. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.H.; Del Tredici, K.; Braak, H. Parkinson’s disease: A dual-hit hypothesis. Neuropathol. Appl. Neurobiol. 2007, 33, 599–614. [Google Scholar] [CrossRef] [PubMed]
- Roberts, E. Alzheimer’s disease may begin in the nose and may be caused by aluminosilicates. Neurobiol. Aging 1986, 7, 561–567. [Google Scholar] [CrossRef]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015, 30, 1591–1601. [Google Scholar] [CrossRef]
- Borghammer, P.; Just, M.K.; Horsager, J.; Skjærbæk, C.; Raunio, A.; Kok, E.H.; Savola, S.; Murayama, S.; Saito, Y.; Myllykangas, L.; et al. A postmortem study suggests a revision of the dual-hit hypothesis of Parkinson’s disease. NPJ Park. Dis. 2022, 8, 166. [Google Scholar] [CrossRef]
- Borghammer, P. The α-Synuclein Origin and Connectome Model (SOC Model) of Parkinson’s Disease: Explaining Motor Asymmetry, Non-Motor Phenotypes, and Cognitive Decline. J. Park. Dis. 2021, 11, 455–474. [Google Scholar] [CrossRef]
- Haehner, A.; Hummel, T.; Hummel, C.; Sommer, U.; Junghanns, S.; Reichmann, H. Olfactory loss may be a first sign of idiopathic Parkinson’s disease. Mov. Disord. 2007, 22, 839–842. [Google Scholar] [CrossRef]
- Haehner, A.; Masala, C.; Walter, S.; Reichmann, H.; Hummel, T. Incidence of Parkinson’s disease in a large patient cohort with idiopathic smell and taste loss. J. Neurol. 2018, 266, 339–345. [Google Scholar] [CrossRef]
- Fullard, M.E.; Morley, J.F.; Duda, J.E. Olfactory Dysfunction as an Early Biomarker in Parkinson’s Disease. Neurosci. Bull. 2017, 33, 515–525. [Google Scholar] [CrossRef]
- Rusz, J.; Janzen, A.; Tykalová, T.; Novotný, M.; Zogala, D.; Timmermann, L.; Růžička, E.; Šonka, K.; Dušek, P.; Oertel, W. Dysprosody in Isolated REM Sleep Behavior Disorder with Impaired Olfaction but Intact Nigrostriatal Pathway. Mov. Disord. 2022, 37, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Sportelli, C.; Urso, D.; Jenner, P.; Chaudhuri, K.R. Metformin as a Potential Neuroprotective Agent in Prodromal Parkinson’s Disease—Viewpoint. Front. Neurol. 2020, 11, 556. [Google Scholar] [CrossRef] [PubMed]
- Adler, C.H. Premotor Symptoms and Early Diagnosis of Parkinson’s Disease. Int. J. Neurosci. 2011, 121, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L.; Golbe, L.I.; McKeown, D.A.; Stern, M.B.; Lehrach, C.M.; Crawford, D. Olfactory testing differentiates between progressive supranuclear palsy and idiopathic Parkinson’s disease. Neurology 1993, 43, 962–965. [Google Scholar] [CrossRef]
- Ercoli, T.; Masala, C.; Cadeddu, G.; Mascia, M.M.; Orofino, G.; Gigante, A.F.; Solla, P.; Defazio, G.; Rocchi, L. Does Olfactory Dysfunction Correlate with Disease Progression in Parkinson’s Disease? A Systematic Review of the Current Literature. Brain Sci. 2022, 12, 513. [Google Scholar] [CrossRef]
- Cavaco, S.; Gonçalves, A.; Mendes, A.; Vila-Chã, N.; Moreira, I.; Fernandes, J.; Damásio, J.; Teixeira-Pinto, A.; Lima, A.B. Abnormal Olfaction in Parkinson’s Disease Is Related to Faster Disease Progression. Behav. Neurol. 2015, 2015, 976589. [Google Scholar] [CrossRef]
- Doty, R.L.; Deems, D.A.; Stellar, S. Olfactory dysfunction in parkinsonism: A general deficit unrelated to neurologic signs, disease stage, or disease duration. Neurology 1988, 38, 1237–1244. [Google Scholar] [CrossRef]
- Melis, M.; Sollai, G.; Masala, C.; Pisanu, C.; Cossu, G.; Melis, M.; Sarchioto, M.; Oppo, V.; Morelli, M.; Crnjar, R.; et al. Odor Identification Performance in Idiopathic Parkinson’s Disease Is Associated With Gender and the Genetic Variability of the Olfactory Binding Protein. Chem. Senses 2019, 44, 311–318. [Google Scholar] [CrossRef]
- Doty, R.L. Psychophysical testing of smell and taste function. Handb. Clin. Neurol. 2019, 164, 229–246. [Google Scholar] [CrossRef]
- Chase, B.A.; Markopoulou, K. Olfactory Dysfunction in Familial and Sporadic Parkinson’s Disease. Front. Neurol. 2020, 11, 447. [Google Scholar] [CrossRef]
- Doty, R.L.; Shaman, P.; Dann, M. Development of the university of pennsylvania smell identification test: A standardized microencapsulated test of olfactory function. Physiol. Behav. 1984, 32, 489–502. [Google Scholar] [CrossRef]
- Hummel, T.; Sekinger, B.; Wolf, S.; Pauli, E.; Kobal, G. ‘Sniffin’ Sticks’: Olfactory Performance Assessed by the Combined Testing of Odour Identification, Odor Discrimination and Olfactory Threshold. Chem. Senses 1997, 22, 39–52. [Google Scholar] [CrossRef]
- Doty, R.L.; Laing, D.G. Psychophysical Measurement of Human Olfactory Function. In Handbook of Olfaction and Gustation, 3rd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 225–260. [Google Scholar] [CrossRef]
- Murphy, J.; McKernan, D.P. The Effect of Aggregated Alpha Synuclein on Synaptic and Axonal Proteins in Parkinson’s Disease—A Systematic Review. Biomolecules 2022, 12, 1199. [Google Scholar] [CrossRef]
- Chandra, S. Synucleins. In Encyclopedia of Neuroscience; Academic Press: Cambridge, MA, USA, 2009; pp. 833–837. [Google Scholar] [CrossRef]
- Braak, H.; Ghebremedhin, E.; Rüb, U.; Bratzke, H.; Del Tredici, K. Stages in the development of Parkinson’s disease-related pathology. Cell Tissue Res. 2004, 318, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Lashuel, H.A.; Overk, C.R.; Oueslati, A.; Masliah, E. The many faces of α-synuclein: From structure and toxicity to therapeutic target. Nat. Rev. Neurosci. 2013, 14, 38–48. [Google Scholar] [CrossRef]
- Srinivasan, E.; Chandrasekhar, G.; Anbarasu, K.; Vickram, A.S.; Karunakaran, R.; Rajasekaran, R.; Srikumar, P.S. Alpha-Synuclein Aggregation in Parkinson’s Disease. Front. Med. (Lausanne) 2021, 8, 736978. [Google Scholar] [CrossRef] [PubMed]
- Ghiglieri, V.; Calabrese, V.; Calabresi, P. Alpha-Synuclein: From Early Synaptic Dysfunction to Neurodegeneration. Front. Neurol. 2018, 9, 295. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Rangel, C.; Marin, G.; Hernández-Contreras, K.A.; Vichi-Ramírez, M.M.; Zarate-Calderon, C.; Torres-Pineda, O.; Diaz-Chiguer, D.L.; González, D.D.l.M.; Apo, E.G.; Teco-Cortes, J.A.; et al. Neuroinflammation in Parkinson’s Disease: From Gene to Clinic: A Systematic Review. Int. J. Mol. Sci. 2023, 24, 5792. [Google Scholar] [CrossRef]
- Hawkes, C.H.; Del Tredici, K.; Braak, H. A timeline for Parkinson’s disease. Park. Relat. Disord. 2010, 16, 79–84. [Google Scholar] [CrossRef]
- Smith, T.D.; Bhatnagar, K.P. Anatomy of the olfactory system. Handb. Clin. Neurol. 2019, 164, 17–28. [Google Scholar] [CrossRef]
- Duda, J.E.; Shaha, U.; Arnold, S.E.; Lee, V.M.-Y.; Trojanowski, J.Q. The Expression of α-, β-, and γ-Synucleins in Olfactory Mucosa from Patients with and without Neurodegenerative Diseases. Exp. Neurol. 1999, 160, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Shioya, A.; Sano, T.; Sumikura, H.; Murata, M.; Murayama, S. Lewy body pathology involves the olfactory cells in Parkinson’s disease and related disorders. Mov. Disord. 2016, 31, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Witt, M.; Bormann, K.; Gudziol, V.; Pehlke, K.; Barth, K.; Minovi, A.; Hähner, A.; Reichmann, H.; Hummel, T. Biopsies of olfactory epithelium in patients with Parkinson’s disease. Mov. Disord. 2009, 24, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.A.; Boettner, M.; Alexoudi, A.; Zorenkov, D.; Deuschl, G.; Wedel, T. Can we use peripheral tissue biopsies to diagnose Parkinson’s disease? A review of the literature. Eur. J. Neurol. 2016, 23, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Daniel, S.; Hawkes, C. Preliminary diagnosis of Parkinson’s disease by olfactory bulb pathology. Lancet 1992, 340, 186. [Google Scholar] [CrossRef]
- Del Tredici, K.; Rüb, U.; De Vos, R.A.; Bohl, J.R.; Braak, H. Where Does Parkinson Disease Pathology Begin in the Brain? J. Neuropathol. Exp. Neurol. 2002, 61, 413–426. [Google Scholar] [CrossRef]
- Hawkes, C.H.; Shephard, B.C.; Daniel, S.E. Olfactory dysfunction in Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 1997, 62, 436–446. [Google Scholar] [CrossRef]
- Mundiñano, I.-C.; Caballero, M.-C.; Ordóñez, C.; Hernandez, M.; DiCaudo, C.; Marcilla, I.; Erro, M.-E.; Tuñon, M.-T.; Luquin, M.-R. Increased dopaminergic cells and protein aggregates in the olfactory bulb of patients with neurodegenerative disorders. Acta Neuropathol. 2011, 122, 61–74. [Google Scholar] [CrossRef]
- Flores-Cuadrado, A.; Saiz-Sanchez, D.; Mohedano-Moriano, A.; Lamas-Cenjor, E.; Leon-Olmo, V.; Martinez-Marcos, A.; Ubeda-Bañon, I. Astrogliosis and sexually dimorphic neurodegeneration and microgliosis in the olfactory bulb in Parkinson’s disease. NPJ Park. Dis. 2021, 7, 11. [Google Scholar] [CrossRef]
- Cave, J.W.; Fujiwara, N.; Weibman, A.R.; Baker, H. Cytoarchitectural changes in the olfactory bulb of Parkinson’s disease patients. NPJ Park. Dis. 2016, 2, 16011. [Google Scholar] [CrossRef]
- Pearce, R.K.B.; Hawkes, C.H.; Daniel, S.E. The anterior olfactory nucleus in Parkinson’s disease. Mov. Disord. 1995, 10, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Zapiec, B.; Dieriks, B.V.; Tan, S.; Faull, R.L.M.; Mombaerts, P.; Curtis, M.A. A ventral glomerular deficit in Parkinson’s disease revealed by whole olfactory bulb reconstruction. Brain 2017, 140, 2722–2736. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gu, C.-Z.; Su, J.-B.; Zhu, L.-H.; Zhou, Y.; Huang, H.-Y.; Liu, C.-F. Changes in Olfactory Bulb Volume in Parkinson’s Disease: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0149286. [Google Scholar] [CrossRef]
- Tremblay, C.; Mei, J.; Frasnelli, J. Olfactory bulb surroundings can help to distinguish Parkinson’s disease from non-parkinsonian olfactory dysfunction. NeuroImage Clin. 2020, 28, 102457. [Google Scholar] [CrossRef]
- Brodoehl, S.; Klingner, C.; Volk, G.F.; Bitter, T.; Witte, O.W.; Redecker, C. Decreased olfactory bulb volume in idiopathic Parkinson’s disease detected by 3.0-Tesla magnetic resonance imaging. Mov. Disord. 2012, 27, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Beach, T.G.; White, C.L.; Hladik, C.L.; Sabbagh, M.N.; Connor, D.J.; Shill, H.A.; Sue, L.I.; Sasse, J.; Bachalakuri, J.; Henry-Watson, J.; et al. Olfactory bulb α-synucleinopathy has high specificity and sensitivity for Lewy body disorders. Acta Neuropathol. 2009, 117, 169–174. [Google Scholar] [CrossRef]
- Ubeda-Bañon, I.; Saiz-Sanchez, D.; de la Rosa-Prieto, C.; Argandoña-Palacios, L.; Garcia-Muñozguren, S.; Martinez-Marcos, A. α-Synucleinopathy in the human olfactory system in Parkinson’s disease: Involvement of calcium-binding protein- and substance P-positive cells. Acta Neuropathol. 2010, 119, 723–735. [Google Scholar] [CrossRef]
- Cersosimo, M.G. Propagation of alpha-synuclein pathology from the olfactory bulb: Possible role in the pathogenesis of dementia with Lewy bodies. Cell Tissue Res. 2018, 373, 233–243. [Google Scholar] [CrossRef]
- Hubbard, P.S.; Esiri, M.M.; Reading, M.; McShane, R.; Nagy, Z. Alpha-synuclein pathology in the olfactory pathways of dementia patients. J. Anat. 2007, 211, 117–124. [Google Scholar] [CrossRef]
- González-Reimers, E.; Martín-González, C.; Romero-Acevedo, L.; Quintero-Platt, G.; Gonzalez-Arnay, E.; Santolaria-Fernández, F. Effects of Alcohol on the Corpus Callosum. In Neuroscience of Alcohol: Mechanisms and Treatment; Academic Press: Cambridge, MA, USA, 2019; pp. 143–152. [Google Scholar] [CrossRef]
- Morgan, J.T.; Nordahl, C.W.; Schumann, C.M. The Amygdala in Autism Spectrum Disorders. In The Neuroscience of Autism Spectrum Disorders; Academic Press: Cambridge, MA, USA, 2013; pp. 297–312. [Google Scholar] [CrossRef]
- Nigro, P.; Chiappiniello, A.; Simoni, S.; Paoletti, F.P.; Cappelletti, G.; Chiarini, P.; Filidei, M.; Eusebi, P.; Guercini, G.; Santangelo, V.; et al. Changes of olfactory tract in Parkinson’s disease: A DTI tractography study. Neuroradiology 2021, 63, 235–242. [Google Scholar] [CrossRef]
- Chen, S.; Tan, H.-Y.; Wu, Z.-H.; Sun, C.-P.; He, J.-X.; Li, X.-C.; Shao, M. Imaging of olfactory bulb and gray matter volumes in brain areas associated with olfactory function in patients with Parkinson’s disease and multiple system atrophy. Eur. J. Radiol. 2014, 83, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D. Olfactory Cortex Physiology. In Encyclopedia of Neuroscience; Academic Press: Cambridge, MA, USA, 2009; pp. 95–100. [Google Scholar] [CrossRef]
- Zilles, K.; Amunts, K. Architecture of the Cerebral Cortex. In The Human Nervous System, Third Edition; Academic Press: Cambridge, MA, USA, 2012; pp. 836–895. [Google Scholar] [CrossRef]
- Vaughan, D.N.; Jackson, G. The Piriform Cortex and Human Focal Epilepsy. Front. Neurol. 2014, 5, 259. [Google Scholar] [CrossRef] [PubMed]
- Plailly, J.; Bensafi, M.; Pachot-Clouard, M.; Delon-Martin, C.; Kareken, D.A.; Rouby, C.; Segebarth, C.; Royet, J.-P. Involvement of right piriform cortex in olfactory familiarity judgments. Neuroimage 2005, 24, 1032–1041. [Google Scholar] [CrossRef]
- Silveira-Moriyama, L.; Holton, J.L.; Kingsbury, A.; Ayling, H.; Petrie, A.; Sterlacci, W.; Poewe, W.; Maier, H.; Lees, A.J.; Revesz, T. Regional differences in the severity of Lewy body pathology across the olfactory cortex. Neurosci. Lett. 2009, 453, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.-Y.; Eslinger, P.J.; Du, G.; Kong, L.; Lewis, M.M.; Huang, X. Olfactory-related cortical atrophy is associated with olfactory dysfunction in Parkinson’s disease. Mov. Disord. 2014, 29, 1205–1208. [Google Scholar] [CrossRef]
- Wattendorf, E.; Welge-Lüssen, A.; Fiedler, K.; Bilecen, D.; Wolfensberger, M.; Fuhr, P.; Hummel, T.; Westermann, B. Olfactory Impairment Predicts Brain Atrophy in Parkinson’s Disease. J. Neurosci. 2009, 29, 15410–15413. [Google Scholar] [CrossRef]
- Wu, X.; Yu, C.; Fan, F.; Zhang, K.; Zhu, C.; Wu, T.; Li, K.; Chan, P. Correlation between Progressive Changes in Piriform Cortex and Olfactory Performance in Early Parkinson’s Disease. Eur. Neurol. 2011, 66, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Duvernoy, H.; Cattin, F.; Risold, P.-Y. Anatomy. In The Human Hippocampus; Springer: Berlin/Heidelberg, Germany, 2013; pp. 39–68. [Google Scholar] [CrossRef]
- Xiao, Q.; Chen, S.; Le, W. Hyposmia: A possible biomarker of Parkinson’s disease. Neurosci. Bull. 2014, 30, 134–140. [Google Scholar] [CrossRef]
- Knierim, J.J. The hippocampus. Curr. Biol. 2015, 25, R1116–R1121. [Google Scholar] [CrossRef]
- Roh, H.; Kang, J.; Koh, S.; Kim, J.H. Hippocampal volume is related to olfactory impairment in Parkinson’s disease. J. Neuroimaging 2021, 31, 1176–1183. [Google Scholar] [CrossRef]
- Bohnen, N.I.; Gedela, S.; Kuwabara, H.; Constantine, G.M.; Mathis, C.A.; Studenski, S.A.; Moore, R.Y. Selective hyposmia and nigrostriatal dopaminergic denervation in Parkinson’s disease. J. Neurol. 2007, 254, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Westermann, B.; Wattendorf, E.; Schwerdtfeger, U.; Husner, A.; Fuhr, P.; Gratzl, O.; Hummel, T.; Bilecen, D.; Welge-Lussen, A. Functional imaging of the cerebral olfactory system in patients with Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 2008, 79, 19–24. [Google Scholar] [CrossRef]
- Jonesgotman, M.; Zatorre, R. Odor Recognition Memory in Humans: Role of Right Temporal and Orbitofrontal Regions. Brain Cogn. 1993, 22, 182–198. [Google Scholar] [CrossRef] [PubMed]
- Frasnelli, J.; Lundström, J.N.; Boyle, J.A.; Djordjevic, J.; Zatorre, R.J.; Jones-Gotman, M. Neuroanatomical correlates of olfactory performance. Exp. Brain Res. 2010, 201, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dahmani, L.; Patel, R.M.; Yang, Y.; Chakravarty, M.M.; Fellows, L.K.; Bohbot, V.D. An intrinsic association between olfactory identification and spatial memory in humans. Nat. Commun. 2018, 9, 4162. [Google Scholar] [CrossRef]
- Li, W.; Lopez, L.; Osher, J.; Howard, J.D.; Parrish, T.B.; Gottfried, J.A. Right Orbitofrontal Cortex Mediates Conscious Olfactory Perception. Psychol. Sci. 2010, 21, 1454–1463. [Google Scholar] [CrossRef]
- Royet, J.-P.; Plailly, J.; Delon-Martin, C.; Kareken, D.A.; Segebarth, C. fMRI of emotional responses to odors: Influence of hedonic valence and judgment, handedness, and gender. Neuroimage 2003, 20, 713–728. [Google Scholar] [CrossRef]
- Bitter, T.; Gudziol, H.; Burmeister, H.P.; Mentzel, H.-J.; Guntinas-Lichius, O.; Gaser, C. Anosmia Leads to a Loss of Gray Matter in Cortical Brain Areas. Chem. Senses 2010, 35, 407–415. [Google Scholar] [CrossRef]
- Masaoka, Y.; Yoshimura, N.; Inoue, M.; Kawamura, M.; Homma, I. Impairment of odor recognition in Parkinson’s disease caused by weak activations of the orbitofrontal cortex. Neurosci. Lett. 2007, 412, 45–50. [Google Scholar] [CrossRef]
- Jesmanas, S.; Gleiznienė, R.; Baranauskas, M.; Matijošaitis, V.; Rastenytė, D. Odor Identification and Regional Gray Matter Atrophy in Patients with Alzheimer’s Disease, Parkinson’s Disease, and the Healthy Elderly: A Cross-Sectional Structural MRI Study. Brain Sci. 2021, 11, 1296. [Google Scholar] [CrossRef]
- Ibarretxe-Bilbao, N.; Junque, C.; Marti, M.-J.; Valldeoriola, F.; Vendrell, P.; Bargallo, N.; Zarei, M.; Tolosa, E. Olfactory impairment in Parkinson’s disease and white matter abnormalities in central olfactory areas: A voxel-based diffusion tensor imaging study. Mov. Disord. 2010, 25, 1888–1894. [Google Scholar] [CrossRef]
- Noto, T.; Zhou, G.; Yang, Q.; Lane, G.; Zelano, C. Human Primary Olfactory Amygdala Subregions Form Distinct Functional Networks, Suggesting Distinct Olfactory Functions. Front. Syst. Neurosci. 2021, 15, 752320. [Google Scholar] [CrossRef]
- Harding, A.J.; Stimson, E.; Henderson, J.M.; Halliday, G.M. Clinical correlates of selective pathology in the amygdala of patients with Parkinson’s disease. Brain 2002, 125, 2431–2445. [Google Scholar] [CrossRef]
- Wang, J.; Sun, L.; Chen, L.; Sun, J.; Xie, Y.; Tian, D.; Gao, L.; Zhang, D.; Xia, M.; Wu, T. Common and distinct roles of amygdala subregional functional connectivity in non-motor symptoms of Parkinson’s disease. NPJ Park. Dis. 2023, 9, 28. [Google Scholar] [CrossRef]
- Yoneyama, N.; Watanabe, H.; Kawabata, K.; Bagarinao, E.; Hara, K.; Tsuboi, T.; Tanaka, Y.; Ohdake, R.; Imai, K.; Masuda, M.; et al. Severe hyposmia and aberrant functional connectivity in cognitively normal Parkinson’s disease. PLoS ONE 2018, 13, e0190072. [Google Scholar] [CrossRef] [PubMed]
- Ibarretxe-Bilbao, N.; Junque, C.; Tolosa, E.; Marti, M.-J.; Valldeoriola, F.; Bargallo, N.; Zarei, M. Neuroanatomical correlates of impaired decision-making and facial emotion recognition in early Parkinson’s disease. Eur. J. Neurosci. 2009, 30, 1162–1171. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, H.; Faber, J.; Timmann, D.; Klockgether, T. Update cerebellum and cognition. J. Neurol. 2021, 268, 3921–3925. [Google Scholar] [CrossRef] [PubMed]
- Sobel, N.; Prabhakaran, V.; Hartley, C.A.; Desmond, J.E.; Zhao, Z.; Glover, G.H.; Gabrieli, J.D.; Sullivan, E.V. Odorant-Induced and Sniff-Induced Activation in the Cerebellum of the Human. J. Neurosci. 1998, 18, 8990–9001. [Google Scholar] [CrossRef]
- Savic, I.; Gulyas, B.; Larsson, M.; Roland, P. Olfactory Functions Are Mediated by Parallel and Hierarchical Processing. Neuron 2000, 26, 735–745. [Google Scholar] [CrossRef]
- Ferdon, S.; Murphy, C. The cerebellum and olfaction in the aging brain: A functional magnetic resonance imaging study. Neuroimage 2003, 20, 12–21. [Google Scholar] [CrossRef]
- Cerf-Ducastel, B.; Murphy, C. Neural substrates of cross-modal olfactory recognition memory: An fMRI study. Neuroimage 2006, 31, 386–396. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, X.; Jing, B.; Hu, B.; Ai, Z.; Xing, B.; Jiang, T.; Peng, P. Cerebellar involvement in olfaction: An fMRI Study. J. Neuroimaging 2021, 31, 517–523. [Google Scholar] [CrossRef] [PubMed]
- García, L.; García-Bañuelos, P.; Aranda-Abreu, G.; Herrera-Meza, G.; Coria-Avila, G.; Manzo, J. Activation of the cerebellum by olfactory stimulation in sexually naive male rats. Neurologia 2015, 30, 264–269. [Google Scholar] [CrossRef]
- Gao, X.; Su, B.; Sun, Z.; Xu, L.; Wei, Y.; Wu, D. Patterns of Gray and White Matter Volume Alterations in Patients With Post-Traumatic Anosmia: A Voxel-Based Morphometry Study. Front. Neurol. 2022, 13, 690760. [Google Scholar] [CrossRef] [PubMed]
- Galvez, V.; Diaz, R.; Hernandez-Castillo, C.R.; Campos-Romo, A.; Fernandez-Ruiz, J. Olfactory performance in spinocerebellar ataxia type 7 patients. Park. Relat. Disord. 2014, 20, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Abele, M.; Riet, A.; Hummel, T.; Klockgether, T. Olfactory dysfunction in cerebellar ataxia and multiple system atrophy. J. Neurol. 2003, 250, 1453–1455. [Google Scholar] [CrossRef]
- Sobel, N.; Thomason, M.E.; Stappen, I.; Tanner, C.M.; Tetrud, J.W.; Bower, J.M.; Sullivan, E.V.; Gabrieli, J.D.E. An impairment in sniffing contributes to the olfactory impairment in Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2001, 98, 4154–4159. [Google Scholar] [CrossRef]
- Mainland, J.D.; Johnson, B.N.; Khan, R.; Ivry, R.B.; Sobel, N. Olfactory Impairments in Patients with Unilateral Cerebellar Lesions Are Selective to Inputs from the Contralesional Nostril. J. Neurosci. 2005, 25, 6362–6371. [Google Scholar] [CrossRef]
- Sharer, J.D.; Leon-Sarmiento, F.E.; Morley, J.F.; Weintraub, D.; Doty, R.L. Olfactory dysfunction in Parkinson’s disease: Positive effect of cigarette smoking. Mov. Disord. 2015, 30, 859–862. [Google Scholar] [CrossRef]
- Iannilli, E.; Stephan, L.; Hummel, T.; Reichmann, H.; Haehner, A. Olfactory impairment in Parkinson’s disease is a consequence of central nervous system decline. J. Neurol. 2017, 264, 1236–1246. [Google Scholar] [CrossRef]
- Rey, N.L.; Wesson, D.W.; Brundin, P. The olfactory bulb as the entry site for prion-like propagation in neurodegenerative diseases. Neurobiol. Dis. 2018, 109, 226–248. [Google Scholar] [CrossRef]
- Hummel, T.; Witt, M.; Reichmann, H.; Welge-Luessen, A.; Haehner, A. Immunohistochemical, volumetric, and functional neuroimaging studies in patients with idiopathic Parkinson’s disease. J. Neurol. Sci. 2010, 289, 119–122. [Google Scholar] [CrossRef]
- Garcia-Esparcia, P.; Schlüter, A.; Carmona, M.; Moreno, J.; Ansoleaga, B.; Torrejón-Escribano, B.; Gustincich, S.; Pujol, A.; Ferrer, I. Functional Genomics Reveals Dysregulation of Cortical Olfactory Receptors in Parkinson Disease: Novel Putative Chemoreceptors in the Human Brain. J. Neuropathol. Exp. Neurol. 2013, 72, 524–539. [Google Scholar] [CrossRef]
- Kohl, Z.; Schlachetzki, J.C.M.; Feldewerth, J.; Hornauer, P.; Münch, M.; Adame, A.; Riemenschneider, M.J.; Winkler, J.; Masliah, E. Distinct Pattern of Microgliosis in the Olfactory Bulb of Neurodegenerative Proteinopathies. Neural Plast. 2017, 2017, 3851262. [Google Scholar] [CrossRef]
- Borghammer, P.; Berge, N.V.D. Brain-First versus Gut-First Parkinson’s Disease: A Hypothesis. J. Park. Dis. 2019, 9, S281–S295. [Google Scholar] [CrossRef] [PubMed]
- Kjelvik, G.; Evensmoen, H.R.; Brezova, V.; Håberg, A.K. The human brain representation of odor identification. J. Neurophysiol. 2012, 108, 645–657. [Google Scholar] [CrossRef]
- Barbey, A.K.; Koenigs, M.; Grafman, J. Orbitofrontal Contributions to Human Working Memory. Cereb. Cortex 2011, 21, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.S.; Chung, S.J.; Lee, Y.H.; Ye, B.S.; Sohn, Y.H.; Lee, P.H. Association between Olfactory Deficit and Motor and Cognitive Function in Parkinson’s Disease. J. Mov. Disord. 2020, 13, 133–141. [Google Scholar] [CrossRef]
- Nag, S.; Yu, L.; VanderHorst, V.G.; Schneider, J.A.; Bennett, D.A.; Buchman, A.S.; Wilson, R.S. Neocortical Lewy bodies are associated with impaired odor identification in community-dwelling elders without clinical PD. J. Neurol. 2019, 266, 3108–3118. [Google Scholar] [CrossRef]
- Hedner, M.; Larsson, M.; Arnold, N.; Zucco, G.M.; Hummel, T. Cognitive factors in odor detection, odor discrimination, and odor identification tasks. J. Clin. Exp. Neuropsychol. 2010, 32, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Burton, E.J.; Mukaetova-Ladinska, E.B.; Perry, R.H.; Jaros, E.; Barber, R.; O’Brien, J.T. Neuropathological correlates of volumetric MRI in autopsy-confirmed Lewy body dementia. Neurobiol. Aging 2012, 33, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Mason, D.M.; Nouraei, N.; Pant, D.B.; Miner, K.M.; Hutchison, D.F.; Luk, K.C.; Stolz, J.F.; Leak, R.K. Transmission of α-synucleinopathy from olfactory structures deep into the temporal lobe. Mol. Neurodegener. 2016, 11, 49. [Google Scholar] [CrossRef] [PubMed]
- Ross, G.W.; Abbott, R.D.; Petrovitch, H.; Tanner, C.M.; Davis, D.G.; Nelson, J.; Markesbery, W.R.; Hardman, J.; Masaki, K.; Launer, L.; et al. Association of olfactory dysfunction with incidental Lewy bodies. Mov. Disord. 2006, 21, 2062–2067. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.S.; Yu, L.; Schneider, J.A.; Arnold, S.E.; Buchman, A.S.; Bennett, D.A. Lewy Bodies and Olfactory Dysfunction in Old Age. Chem. Senses 2011, 36, 367–373. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Pasillas, G.; Chi-Castañeda, D.; Carrillo-Castilla, P.; Marín, G.; Hernández-Aguilar, M.E.; Aranda-Abreu, G.E.; Manzo, J.; García, L.I. Olfactory Dysfunction in Parkinson’s Disease, Its Functional and Neuroanatomical Correlates. NeuroSci 2023, 4, 134-151. https://doi.org/10.3390/neurosci4020013
Torres-Pasillas G, Chi-Castañeda D, Carrillo-Castilla P, Marín G, Hernández-Aguilar ME, Aranda-Abreu GE, Manzo J, García LI. Olfactory Dysfunction in Parkinson’s Disease, Its Functional and Neuroanatomical Correlates. NeuroSci. 2023; 4(2):134-151. https://doi.org/10.3390/neurosci4020013
Chicago/Turabian StyleTorres-Pasillas, Gabriel, Donají Chi-Castañeda, Porfirio Carrillo-Castilla, Gerardo Marín, María Elena Hernández-Aguilar, Gonzalo Emiliano Aranda-Abreu, Jorge Manzo, and Luis I. García. 2023. "Olfactory Dysfunction in Parkinson’s Disease, Its Functional and Neuroanatomical Correlates" NeuroSci 4, no. 2: 134-151. https://doi.org/10.3390/neurosci4020013
APA StyleTorres-Pasillas, G., Chi-Castañeda, D., Carrillo-Castilla, P., Marín, G., Hernández-Aguilar, M. E., Aranda-Abreu, G. E., Manzo, J., & García, L. I. (2023). Olfactory Dysfunction in Parkinson’s Disease, Its Functional and Neuroanatomical Correlates. NeuroSci, 4(2), 134-151. https://doi.org/10.3390/neurosci4020013