
Social Touch: Its Mirror-like Responses and Implications in Neurological and Psychiatric Diseases


Abstract
1. Introduction
2. CT Fibers: The Discovery
3. Properties of CT Fibers
4. Effects of CT Fibers on the Autonomous Nervous System
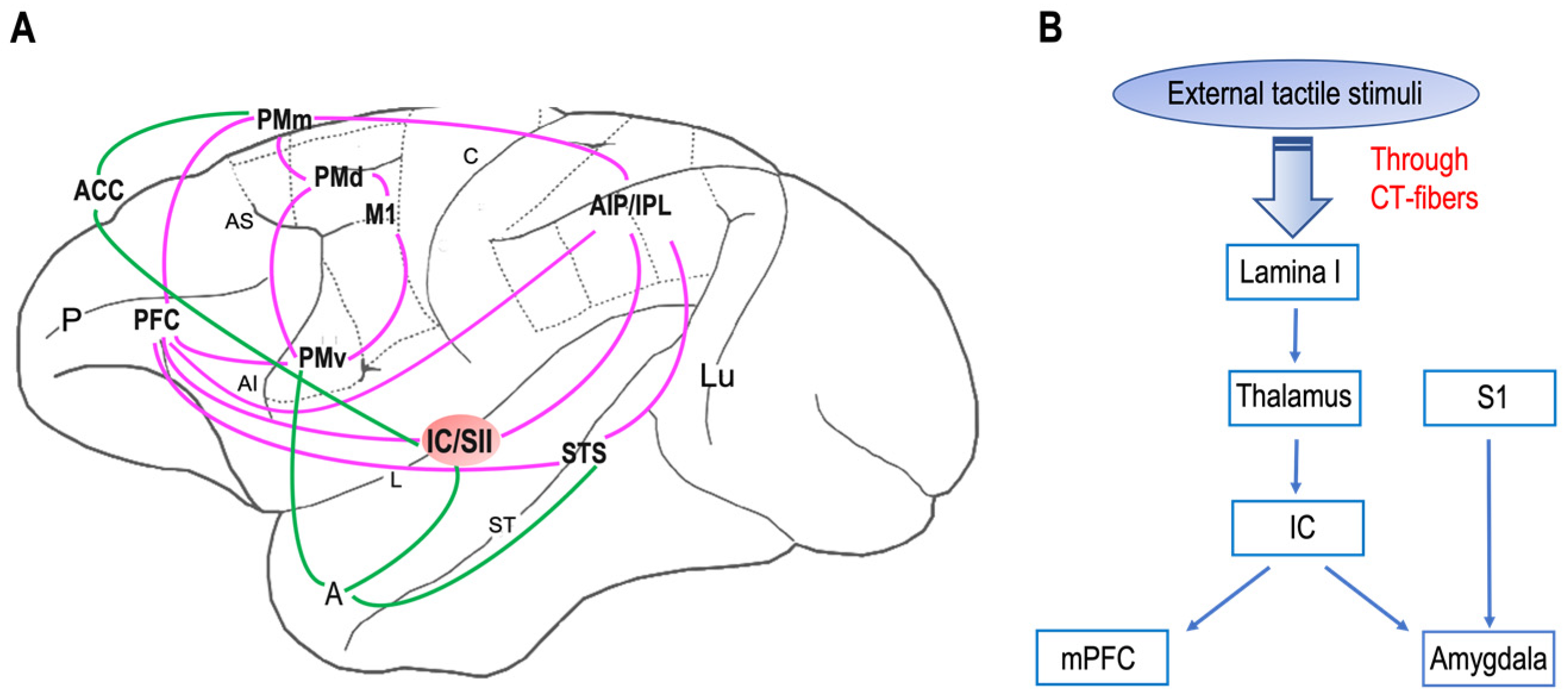
5. Central Nervous System
- The putamen;
- The orbitofrontal cortex;
- The posterosuperior temporal sulcus;
- The medial prefrontal cortex;
- The dorsal–anterior cingulate cortex;
- The pregenual anterior cingulate cortex;
- Superior temporal sulcus.
6. Social Touch and Mirror Neurons
7. From Social Brain to Social Behavior
8. Social Touch and Its Implication for Neurologic and Psychiatric Disorders
9. Touch as Therapy?
10. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hertenstein, M.J.; Verkamp, J.M.; Kerestes, A.M.; Holmes, R.M. The communicative functions of touch in humans, nonhuman primates, and rats: A review and synthesis of the empirical research. Genet. Soc. Gen. Psychol. Monogr. 2006, 132, 5–94. Available online: https://www.depauw.edu/learn/lab/publications/documents/touch/2006_Touch_The%2520communicative_functions_of_touch_in_humans.pdf (accessed on 20 March 2023). [CrossRef] [PubMed]
- Boccia, M.L. Comparison of the physical characteristics of grooming in two species of macaques (Macaca nemestrina and M. radiata). J. Comp. Psychol. 1989, 103, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Boccia, M.L.; Reite, M.; Laudenslager, M. On the physiology of grooming in a pigtail macaque. Physiol. Behav. 1989, 45, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, R.I. The social role of touch in humans and primates: Behavioural function and neurobiological mechanisms. Neurosci. Biobehav. Rev. 2010, 34, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Gallace, A.; Spence, C. The science of interpersonal touch: An overview. Neurosci. Biobehav. Rev. 2010, 34, 246–259. [Google Scholar] [CrossRef]
- Castiello, U.; Becchio, C.; Zoia, S.; Nelini, C.; Sartori, L.; Blason, L.; D’Ottavio, G.; Bulgheroni, M.; Gallese, V. Wired to be social: The ontogeny of human interaction. PLoS ONE 2010, 5, e13199. [Google Scholar] [CrossRef]
- Gliga, T.; Farroni, T.; Cascio, C.J. Social touch: A new vista for developmental cognitive neuroscience? Dev. Cogn. Neurosci. 2019, 35, 1–4. [Google Scholar] [CrossRef]
- Nelson, C.A.; Fox, N.A.; Zeanah, C.H. Romania’s Abandoned Children. Deprivation, Brain Development, and the Struggle for Recovery; Harvard University Press: Cambridge, UK, 2014. [Google Scholar]
- Cascio, C.J.; Cascio, D.; Moore, F. McGlone Social touch and human development Dev. Cogn. Neurosci. 2019, 35, 5–11. [Google Scholar] [CrossRef]
- Kirsch, L.P.; Krahé, C.; Blom, N.; Crucianelli, L.; Moro, V.; Jenkinson, P.M.; Fotopoulou, A. Reading the mind in the touch: Neurophysiological specificity in the communication of emotions by touch. Neuropsychologia. 2018, 116, 136–149. [Google Scholar] [CrossRef]
- Croy, I.; Luong, A.; Triscoli, C.; Hofmann, E.; Olausson, H.; Sailer, U. Interpersonal stroking touch is targeted to C tactile afferent activation. Behav. Brain Res. 2016, 297, 37–40. [Google Scholar] [CrossRef]
- Morrison, I.; Bjornsdotter, M.; Olausson, H. Vicarious responses to social touch in posterior insular cortex are tuned to pleasant caressing speeds. J. Neurosci. 2011, 31, 554–9562. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.; Lee, S.H. From Neurons to Social Beings: Short Review of the Mirror Neuron System Research and Its Socio-Psychological and Psychiatric Implications. Clin. Psychopharmacol. Neurosci. 2018, 16, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Croy, I.; Geide, H.; Paulus, M.; Weidner, K.; Olausson, H. Affective touch awareness in mental health and disease relates to autistic traits—An explorative neurophysiological investigation. Psychiatry Res. 2016, 245, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Douglas, W.W.; Ritchie, J.M. On the Frequency of Firing of Mammalian Nonmedullated Nerve Fibers. J. Physiol. 1957, 139, 400–407. [Google Scholar] [CrossRef]
- Bessou, P.; Burgess, P.R.; Perl, E.R.; Taylor, C.B. Dynamic Properties of Mechanoreceptors with Unmyelinated (C) Fibers. J. Neurophysiol. 1971, 34, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Iggo, A.; Kornhuber, H.H. A Quantitative Study of C-Mechanoreceptors in Hairy Skin of the Cat. J. Physiol. 1977, 271, 549–565. [Google Scholar] [CrossRef]
- Kumazawa, T.; Perl, E.R. Excitation of Marginal and Substantia Gelatinosa Neurons in the Primate Spinal Cord: Indications of Their Place in Dorsal Horn Functional Organization. J. Comp. Neurol. 1978, 177, 417–434. [Google Scholar] [CrossRef]
- Johansson, R.S.; Trulsson, M.; Olsson, K.A.; Abbs, J.H. Mechanoreceptive afferent activity in the infraorbital nerve in man during speech and chewing movements. Exp. Brain Res. 1988, 72, 209–214. [Google Scholar] [CrossRef]
- Nordin, M. Low-threshold mechanoreceptive and nociceptive units with unmyelinated (C) fibers in the human supraorbital nerve. J. Physiol. 1990, 426, 229–240. [Google Scholar] [CrossRef]
- Vallbo, A.B.; Olausson, H.; Wessberg, J.; Kakuda, N. Receptive field characteristics of tactile units with myelinated afferents in hairy skin of human subjects. J. Physiol. 1995, 483, 783–79595. [Google Scholar] [CrossRef]
- Morrison, I.; Löken, L.S.; Olausson, H. The skin as a social organ. Exp. Brain Res. 2010, 204, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Olausson, H.; Wessberg, J.; Morrison, I.; McGlone, F.; Vallbo, A.B. The Neurophysiology of Unmyelinated Tactile Afferents. Neurosci. Biobehav. Rev. 2010, 34, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Vallbo, A.B.; Olausson, H.; Wessberg, J. Unmyelinated afferents constitute a second system coding tactile stimuli of the human hairy skin. J. Neurophysiol. 1999, 81, 2753–2763. [Google Scholar] [CrossRef] [PubMed]
- Wiklund Fernström, K.; Wessberg, J.; Vallbo, A. Temperature response of unmyelinated low-threshold mechanoreceptors (CT) in human hairy skin. Soc. Neurosci. Abstr. 2003, 29, 585.8. [Google Scholar]
- Olausson, H.; Cole, J.; Vallbo, A.B.; McGlone, F.; Elam, M.; Kramer, H.H.; Rylander, K.; Wessberg, J.; Elam, M.; Bushnell, M.C. Unmyelinated tactile afferents have opposite effects on insular and somatosensory cortical processing. Neurosci. Lett. 2008, 436, 128–132. [Google Scholar] [CrossRef]
- Olausson, H.; Cole, J.; Rylander, K.; McGlone, F.; Lamarre, Y.; Gunnar Wallin, B.; Krämer, H.; Wessberg, J.; Elam, M.; Bushnell, M.C.; et al. Functional role of unmyelinated tactile afferents in human hairy skin: Sympathetic response and perceptual localization. Exp. Brain Res. 2008, 184, 135–140. [Google Scholar] [CrossRef]
- Pomeranz, B.; Macaulay, R.J.; Caudill, M.A.; Kutz, I.; Adam, D.; Gordon, D.; Kilborn, K.M.; Barger, A.C.; Shannon, D.C.; Cohen, R.J.; et al. Assessment of autonomic function in humans by heart rate spectral analysis. Am. J. Physiol. 1985, 248, H151–H153. [Google Scholar] [CrossRef]
- Porges, S.W. Cardiac vagal tone: A physiological index of stress. Neurosci. Biobehav. Rev. 1995, 19, 225–233. [Google Scholar] [CrossRef]
- Triscoli, C.; Croy, I.; Olausson, H.; Sailer, U. Touch between romantic partners: Being stroked is more pleasant than stroking and decelerates heart rate. Physiol. Behav. 2017, 177, 169–175. [Google Scholar] [CrossRef]
- Savallampi, M.; Maallo, A.M.S.; Shaikh, S.; McGlone, F.; Bariguian-Revel, F.J.; Olausson, H.; Boehme, R. Social Touch Reduces Pain Perception—An Fmri Study of Cortical Mechanisms. Brain Sci. 2023, 13, 393. [Google Scholar] [CrossRef]
- Merskey, H.; Bogduk, N. Classification of Chronic Pain, 2nd ed.; IASP Press: Seattle, WA, USA, 1994; p. 1. [Google Scholar]
- Meijer, L.L.; Ruis, C.; van der Smagt, M.J.; Dijkerman, H.C. Chronic pain relief after receiving affective touch: A single case report. J. Neuropsychol. 2023, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.A.; Caldwell, J.D.; Drago, F.; Noonan, L.R.; Peterson, G.; Hood, L.E.; Prange, A.J., Jr. Grooming behavioral effects of oxytocin. Pharmacology, ontogeny, and comparisons with other nonapeptides. Ann. N. Y. Acad. Sci. 1988, 525, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Drago, F.; Pedersen, C.A.; Caldwell, J.D.; Prange, A.J., Jr. Oxytocin potently enhances novelty-induced grooming behavior in the rat. Brain Res. 1986, 368, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.; Kano, F.; Yeow, H.; Morimura, N.; Yamamoto, S. Testing the effect of oxytocin on social grooming in bonobos. Am. J. Primatol. 2022, 84, e23444. [Google Scholar] [CrossRef]
- Ellingsen, D.M.; Wessberg, J.; Chelnokova, O.; Olausson, H.; Laeng, B.; Leknes, S. In touch with your emotions: Oxytocin and touch change social impressions while others’ facial expressions can alter touch. Psychoneuroendocrinology 2014, 39, 11–20. [Google Scholar] [CrossRef]
- Kemp, A.H.; Quintana, D.S.; Kuhnert, R.L.; Griffiths, K.; Hickie, I.B.; Guastella, A.J. Oxytocin increases heart rate variability in humans at rest: Implications for social approach-related motivation and capacity for social engagement. PLoS ONE 2012, 7, e44014. [Google Scholar] [CrossRef]
- Boehme, R.; Hauser, S.; Gerling, G.J.; Heilig, M.; Olausson, H. Distinction of self-produced touch and social touch at cortical and spinal cord levels. Proc. Natl. Acad. Sci. USA 2019, 116, 2290–2299. [Google Scholar] [CrossRef]
- Light, A.R.; Perl, E.R. Spinal Termination of Functionally Identified Primary Afferent Neurons with Slowly Conducting Myelinated Fibers. J. Comp. Neurol. 1979, 186, 133–150. [Google Scholar] [CrossRef]
- Sugiura, Y.; Lee, C.L.; Perl, E.R. Central Projections of Identified, Unmyelinated (C) Afferent Fibers Innervating Mammalian Skin. Science 1986, 234, 358–361. [Google Scholar] [CrossRef]
- Björnsdotter, M.; Löken, L.; Olausson, H.; Vallbo, A.; Wessberg, J. Somatotopic organization of gentle touch processing in the posterior insular cortex. J. Neurosci. 2009, 29, 9314–9320. [Google Scholar] [CrossRef]
- Kringelbach, M.L.; Rolls, E.T. The functional neuroanatomy of the human orbitofrontal cortex: Evidence from neuroimaging and neuropsychology. Prog. Neurobiol. 2004, 72, 341–372. [Google Scholar] [CrossRef] [PubMed]
- Olausson, H.; Lamarre, Y.; Backlund, H.; Morin, C.; Wallin, B.G.; Starck, G.; Ekholm, S.; Strigo, I.; Worsely, K.; Vallbo, A.B.; et al. Unmyelinated tactile afferents signal touch and project to insular cortex. Nat. Neurosci. 2002, 5, 900–904. [Google Scholar] [CrossRef] [PubMed]
- Gordon, I.; Voos, A.C.; Bennett, R.H.; Bolling, D.Z.; Pelphrey, K.A.; Kaiser, M.D. Brain mechanisms for processing affective touch. Hum. Brain Mapp. 2013, 34, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, L.; Westling, G.; Brulin, C.; Lehtipalo, S.; Andersson, M.; Nyberg, L. Pleasant human touch is represented in pregenual anterior cingulate cortex. Neuroimage 2012, 59, 3427–3432. [Google Scholar] [CrossRef]
- McGlone, F.; Valbo, A.B.; Olausson, H.; Löken, L.; Wessberg, J. Discriminative touch and emotional touch. Can. J. Exp. Psychol. 2007, 61, 173–183. [Google Scholar] [CrossRef]
- McGlone, F.; Olausson, H.; Boyle, J.A.; Jones-Gotman, M.; Dancer, C.; Guest, S.; Essick, G. Touching and feeling: Differences in pleasant touch processing between glabrous and hairy skin in humans. Eur. J. Neurosci. 2012, 35, 1782–1788. [Google Scholar] [CrossRef]
- Sailer, U.; Triscoli, C.; Häggblad, G.; Hamilton, P.; Olausson, H.; Croy, I. Temporal dynamics of brain activation during 40 minutes of pleasant touch. Neuroimage 2016, 139, 360–367. [Google Scholar] [CrossRef]
- Bennett, R.H.; Bolling, D.Z.; Anderson, L.C.; Pelphrey, K.A.; Kaiser, M.D. fNIRS detects temporal lobe response to affective touch. Soc. Cogn. Affect. Neurosci. 2014, 9, 470–476. [Google Scholar] [CrossRef]
- McGlone, F.; Wessberg, H.; Olausson, H. Discriminative and affective touch: Sensing and feeling. Neuron 2014, 82, 737–755. [Google Scholar] [CrossRef]
- Cacioppo, J.T.; Decety, J. Social neuroscience: Challenges and opportunities in the study of complex behavior. Ann. N. Y. Acad. Sci. 2011, 1224, 162–173. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Fogassi, L.; Gallese, V. Neurophysiological mechanisms underlying the understanding and imitation of action. Nat. Rev. Neurosci. 2001, 2, 661–670. [Google Scholar] [CrossRef] [PubMed]
- di Pellegrino, G.; Fadiga, L.; Fogassi, L.; Gallese, V.; Rizzolatti, G. Understanding motor events: A neurophysiological study. Exp. Brain Res. 1992, 91, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Rozzi, S.; Ferrari, P.F.; Bonini, L.; Rizzolatti, G.; Fogassi, L. Functional organization of inferior parietal lobule convexity in the macaque monkey: Electrophysiological characterization of motor, sensory and mirror responses and their correlation with cytoarchitectonic areas. Eur. J. Neurosci. 2008, 28, 1569–1588. [Google Scholar] [CrossRef] [PubMed]
- Goldman, A.; Gallese, V. Reply to Schulkin. Trends Cogn. Sci. 2000, 4, 255–256. [Google Scholar] [CrossRef]
- Keysers, C.; Wicker, B.; Gazzola, V.; Anton, J.L.; Fogassi, L.; Gallese, V. A touching sight: SII/PV activation during the observation and experience of touch. Neuron 2004, 42, 335–346. [Google Scholar] [CrossRef]
- Holle, H.; Banissy, M.J.; Ward, J. Functional and structural brain differences associated with mirror-touch synaesthesia. Neuroimage 2013, 83, 1041–1050. [Google Scholar] [CrossRef]
- Gallese, V. The manifold nature of interpersonal relations: The quest for a common mechanism. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2003, 358, 517–528. [Google Scholar] [CrossRef]
- Pitcher, D.; Garrido, L.; Walsh, V.; Duchaine, B.C. Transcranial magnetic stimulation disrupts the perception and embodiment of facial expressions. J. Neurosci. 2008, 28, 8929–8933. [Google Scholar] [CrossRef]
- Ebisch, S.J.; Ferri, F.; Salone, A.; Perrucci, M.G.; D’Amico, L.; Ferro, F.M.; Romani, G.L.; Gallese, V. Differential involvement of somatosensory and interoceptive cortices during the observation of affective touch. J. Cogn. Neurosci. 2011, 23, 1808–1822. [Google Scholar] [CrossRef]
- Ishida, H.; Fornia, L.; Grandi, L.C.; Umiltà, M.A.; Gallese, V. Somato-motor haptic processing in posterior inner perisylvian region (SII/pIC) of the macaque monkey. PLoS ONE 2013, 30, e69931. [Google Scholar] [CrossRef]
- Schaefer, M.; Heinze, H.J.; Rotte, M. Touch and personality: Extraversion predicts somatosensory brain response. Neuroimage 2012, 62, 432–438. [Google Scholar] [CrossRef] [PubMed]
- McCabe, C.; Rolls, E.T.; Bilderbeck, A.; McGlone, F. Cognitive influences on the affective representation of touch and the sight of touch in the human brain. Soc. Cogn. Affect. Neurosci. 2008, 3, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Macefield, V.G.; Norcliffe-Kaufmann, L.; Löken, L.; Axelrod, F.B.; Kaufmann, H. Disturbances in affective touch in hereditary sensory & autonomic neuropathy type III. Int. J. Psychophysiol. 2014, 93, 56–61. [Google Scholar] [PubMed]
- Martinez, V.R.; Giovanola, Y.; Ionta, S. Social Touch Somatotopically Affects Mental Body Representations. Neuroscience 2022, 494, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, G.S.P.; Salgado, J.A.D.; Staempfli, P.; Seifritz, E.; Gassert, R.; Ionta, S. Illusory Body Ownership Affects the Cortical Response to Vicarious Somatosensation. Cereb. Cortex 2022, 32, 312–328. [Google Scholar] [CrossRef]
- Gallese, V.; Rochat, M.J.; Berchio, C. The mirror mechanism and its potential role in autism spectrum disorder. Dev. Med. Child. Neurol. 2013, 55, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Gallese, V. The roots of empathy: The shared manifold hypothesis and the neural basis of intersubjectivity. Psychopathology 2003, 36, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Gallese, V. Embodied simulation: From neurons to phenomenal experience. Phenomenol. Cogn. Sci. 2005, 4, 23–48. [Google Scholar] [CrossRef]
- Gallese, V. Embodied simulation theory: Imagination and narrative. Neuropsychoanalysis 2011, 13, 196–200. [Google Scholar] [CrossRef]
- Gallese, V.; Sinigaglia, C. What is so special about embodied simulation? Trends Cogn. Sci. 2011, 15, 512–519. [Google Scholar] [CrossRef]
- Gallese, V. Intentional attunement: A neurophysiological perspective on social cognition and its disruption in autism. Brain Res. 2006, 1079, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Bonini, L.; Rotunno, C.; Arcuri, E.; Gallese, V. Mirror neurons 30 years later: Implications and applications. Trends Cogn. Sci. 2022, 26, 767–781. [Google Scholar] [CrossRef] [PubMed]
- Lanzilotto, M.; Ferroni, C.G.; Livi, A.; Gerbella, M.; Maranesi, M.; Borra, E.; Passarelli, L.; Gamberini, M.; Fogassi, L.; Bonini, L.; et al. Anterior Intraparietal Area: A Hub in the Observed Manipulative Action Network. Cereb. Cortex 2019, 29, 1816–1833. [Google Scholar] [CrossRef] [PubMed]
- Bruni, S.; Gerbella, M.; Bonini, L.; Borra, E.; Coudé, G.; Ferrari, P.F.; Fogassi, L.; Maranesi, M.; Rodà, F.; Simone, L.; et al. Cortical and subcortical connections of parietal and premotor nodes of the monkey hand mirror neuron network. Brain Struct. Funct. 2018, 223, 1713–1729. [Google Scholar] [CrossRef]
- Albertini, D.; Gerbella, M.; Lanzilotto, M.; Livi, A.; Maranesi, M.; Ferroni, C.G.; Bonini, L. Connectional gradients underlie functional transitions in monkey pre-supplementary motor area. Prog. Neurobiol. 2020, 184, 101699. [Google Scholar] [CrossRef] [PubMed]
- Ferroni, C.G.; Albertini, D.; Lanzilotto, M.; Livi, A.; Maranesi, M.; Bonini, L. Local and system mechanisms for action execution and observation in parietal and premotor cortices. Curr. Biol. 2021, 31, 2819–2830.e4. [Google Scholar] [CrossRef] [PubMed]
- Vigneswaran, G.; Philipp, R.; Lemon, R.N.; Kraskov, A. M1 corticospinal mirror neurons and their role in movement suppression during action observation. Curr. Biol. 2013, 23, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Bonini 2010 Bonini, L.; Rozzi, S.; Serventi, F.U.; Simone, L.; Ferrari, P.F.; Fogassi, L. Ventral premotor and inferior parietal cortices make distinct contribution to action organization and intention understanding. Cereb. Cortex 2010, 20, 1372–1385. [Google Scholar] [CrossRef]
- Lanzilotto, M.; Maranesi, M.; Livi, A.; Ferroni, C.G.; Orban, G.A.; Bonini, L. Stable readout of observed actions from format-dependent activity of monkey’s anterior intraparietal neurons. Proc. Natl. Acad. Sci. USA 2020, 117, 16596–16605. [Google Scholar] [CrossRef]
- Papadourakis, V.; Raos, V. Neurons in the Macaque Dorsal Premotor Cortex Respond to Execution and Observation of Actions. Cereb. Cortex 2019, 29, 4223–4237. [Google Scholar] [CrossRef]
- Albertini, D.; Lanzilotto, M.; Maranesi, M.; Bonini, L. Largely shared neural codes for biological and nonbiological observed movements but not for executed actions in monkey premotor areas. J. Neurophysiol. 2021, 126, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Livi, A.; Lanzilotto, M.; Maranesi, M.; Fogassi, L.; Rizzolatti, G.; Bonini, L. Agent-based representations of objects and actions in the monkey pre-supplementary motor area. Proc. Natl. Acad. Sci. USA 2019, 116, 2691–2700. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Saito, N.; Iriki, A.; Isoda, M. Representation of others’ action by neurons in monkey medial frontal cortex. Curr. Biol. 2011, 21, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Falcone, R.; Cirillo, R.; Ferraina, S.; Genovesio, A. Neural activity in macaque medial frontal cortex represents others’ choices. Sci. Rep. 2017, 7, 12663. [Google Scholar] [CrossRef] [PubMed]
- Simone, L.; Bimbi, M.; Rodà, F.; Fogassi, L.; Rozzi, S. Action observation activates neurons of the monkey ventrolateral prefrontal cortex. Sci. Rep. 2017, 7, 44378. [Google Scholar] [CrossRef] [PubMed]
- Hihara, S.; Taoka, M.; Tanaka, M.; Iriki, A. Visual Responsiveness of Neurons in the Secondary Somatosensory Area and its Surrounding Parietal Operculum Regions in Awake Macaque Monkeys. Cereb. Cortex 2015, 25, 4535–4550. [Google Scholar] [CrossRef]
- Errante, A.; Fogassi, L. Activation of cerebellum and basal ganglia during the observation and execution of manipulative actions. Sci. Rep. 2020, 10, 12008. [Google Scholar] [CrossRef]
- Livneh, U.; Resnik, J.; Shohat, Y.; Paz, R. Self-monitoring of social facial expressions in the primate amygdala and cingulate cortex. Proc. Natl. Acad. Sci. USA 2012, 109, 18956–18961. [Google Scholar] [CrossRef]
- Caruana, F.; Jezzini, A.; Sbriscia-Fioretti, B.; Rizzolatti, G.; Gallese, V. Emotional and social behaviors elicited by electrical stimulation of the insula in the macaque monkey. Curr. Biol. 2011, 21, 195–199. [Google Scholar] [CrossRef]
- Wicker, B.; Keysers, C.; Plailly, J.; Royet, J.P.; Gallese, V.; Rizzolatti, G. Both of us disgusted in my insula: The common neural basis of seeing and feeling disgust. Neuron 2003, 40, 655–664. [Google Scholar] [CrossRef]
- Davidovic, M.; Starck, G.; Olausson, H. Processing of affective and emotionally neutral tactile stimuli in the insular cortex. Dev. Cogn. Neurosci. 2019, 35, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, E.E.; Ackerley, R.; Lundqvist, D.; Schneiderman, J.; Jousmäki, V.; Wessberg, J. Spatio-temporal profile of brain activity during gentle touch investigated with magnetoencephalography. NeuroImage 2019, 201, 116024. [Google Scholar] [CrossRef] [PubMed]
- Lee Masson, H.; de Beeck, H.O.; Boets, B. Reduced task-dependent modulation of functional network architecture for positive versus negative affective touch processing in autism spectrum disorders. NeuroImage 2020, 219, 117009. [Google Scholar] [CrossRef]
- Perini, I.; Gustafsson, P.A.; Igelström, K.; Jasiunaite-Jokubaviciene, B.; Kämpe, R.; Mayo, L.M.; Molander, J.; Olausson, H.; Zetterqvist, M.; Heilig, M. Altered relationship between subjective perception and central representation of touch hedonics in adolescents with autism-spectrum disorder. Transl. Psychiatry 2021, 11, 224. [Google Scholar] [CrossRef]
- Sterman, A.B.; Schaumburg, H.H.; Asbury, A.K. The acute sensory neuronopathy syndrome: A distinct clinical entity. Ann. Neurol. 1980, 7, 354–358. [Google Scholar] [CrossRef]
- Cole, J.D.; Sedgwick, E.M. The perceptions of force and of movement in a man without large myelinated sensory afferents below the neck. J. Physiol. 1992, 449, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Larsson, E.; Kuma, R.; Norberg, A.; Minde, J.; Holmberg, M. Nerve growth factor R221W responsible for insensitivity to pain is defectively processed and accumulates as proNGF. Neurobiol. Dis. 2009, 33, 221–228. [Google Scholar] [CrossRef]
- Minde, J.; Svensson, O.; Holmberg, M.; Solders, G.; Toolanen, G. Orthopedic aspects of familial insensitivity to pain due to a novel nerve growth factor beta mutation. Acta Orthop. 2006, 77, 198–202. [Google Scholar] [CrossRef]
- Morrison, I.; Löken, L.S.; Minde, J.; Wessberg, J.; Perini, I.; Nennesmo, I.; Olausson, H. Reduced C-afferent fibre density affects perceived pleasantness and empathy for touch. Brain 2011, 134, 1116–1126. [Google Scholar] [CrossRef]
- Crucianelli, L.; Cardi, V.; Treasure, J.; Jenkinson, P.M.; Fotopoulou, A. The perception of affective touch in anorexia nervosa. Psychiatry Res. 2016, 239, 72–78. [Google Scholar] [CrossRef]
- Lee Masson, H.; Pillet, I.; Amelynck, S.; Van De Plas, S.; Hendriks, M.; de Beeck, H.O.; Boets, B. Intact neural representations of affective meaning of touch but lack of embodied resonance in autism: A multi-voxel pattern analysis study. Mol. Autism 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Haggarty, C.J.; Malinowski, P.; McGlone, F.P.; Walker, S.C. Autistic traits modulate cortical responses to affective but not discriminative touch. Eur. J. Neurosci. 2020, 51, 1844–1855. [Google Scholar] [CrossRef] [PubMed]
- Buvuktaskin 2021 Buyuktaskin, D.; Iseri, E.; Guney, E.; Gunendi, Z.; Cengiz, B. Somatosensory temporal discrimination in autism spectrum disorder. Autism Res. 2021, 14, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.M.; Han, Y.M. Differential mirror neuron system (MNS) activation during action observation with and without social-emotional components in autism: A meta analysis of neuroimaging studies. Mol. Autism 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Peled-Avron, L.; Glasner, L.; Gvirts, H.Z.; Shamay-Tsoory, S.G. The role of the inferior frontal gyrus in vicarious social touch: A transcranial direct current stimulation (tDCS) study. Dev. Cogn. Neurosci. 2019, 35, 115–121. [Google Scholar] [CrossRef]
- Gernsbacher, M.A.; Yergeau, M. Empirical failures of the claim that autistic people lack a theory of mind. Arch. Sci. Psychol. 2019, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Haggarty, C.J.; Trotter, P.D.; McGlone, F.; Walker, S.C. Children’s vicarious ratings of social touch are tuned to the velocity but not the location of a caress. PLoS ONE 2021, 16, e0256303. [Google Scholar] [CrossRef]
- Kang, Z.; Xing, H.; Lin, Q.; Meng, F.; Gong, L. Effectiveness of therapeutic massage for improving motor symptoms in Parkinson’s disease: A systematic review and meta-analysis. Front. Neurol. 2022, 13, 915232. [Google Scholar] [CrossRef]
- Billhult, A.; Lindholm, C.; Gunnarsson, R.; Stener-Victorin, E. The effect of massage on immune function and stress in women with breast cancer—A randomized controlled trial. Auton. Neurosci. 2009, 150, 111–115. [Google Scholar] [CrossRef]
- Garner, B.; Phillips, L.J.; Schmidt, H.M.; Markulev, C.; O’Connor, J.; Wood, S.J.; Berger, G.E.; Burnett, P.; McGorry, P.D. Pilot study evaluating the effect of massage therapy on stress, anxiety and aggression in a young adult psychiatric inpatient unit. Aust. N. Z. J. Psychiatry 2008, 42, 414–422. [Google Scholar] [CrossRef]
- Schroeder, B.; Doig, J.; Premkumar, K. The effects of massage therapy on multiple sclerosis patients’ quality of life and leg function. Evid. Based Compl. Alternat. Med. 2014, 2014, 640916. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Q.; Zhang, Q.; Kou, J.; Zhang, Y.; Cui, H.; Wernicke, J.; Montag, C.; Becker, B.; Kendrick, K.M.; et al. The Effects of Intranasal Oxytocin on Neural and Behavioral Responses to Social Touch in the Form of Massage. Front. Neurosci. 2020, 14, 589878. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, S.; Yuhi, T.; Furuhara, K.; Ohta, S.; Shimizu, Y.; Higashida, H. Salivary oxytocin concentrations in seven boys with autism spectrum disorder received massage from their mothers: A pilot study. Front. Psychiatry 2015, 6, 58. [Google Scholar] [CrossRef]
- Carpenter, L.L.; Kronenberg, E.F.; Tirrell, E.; Kokdere, F.; Beck, Q.M.; Temereanca, S.; Fukuda, A.M.; Garikapati, S.; Hagberg, S. Mechanical Affective Touch Therapy for Anxiety Disorders: Feasibility, Clinical Outcomes, and Electroencephalography Bi-omarkers From an Open-Label Trial. Front. Psychiatry 2022, 13, 877574. [Google Scholar] [CrossRef]
- Baumgart, S.B.; Baumbach-Kraft, A.; Lorenz, J. Effect of Psycho-Regulatory Massage Therapy on Pain and Depression in Women with Chronic and/or Somatoform Back Pain: A Randomized Controlled Trial. Brain Sci. 2020, 10, 721. [Google Scholar] [CrossRef] [PubMed]
- Ang, J.Y.; Lua, J.L.; Mathur, A.; Thomas, R.; Asmar, B.I.; Savasan, S.; Buck, S.; Long, M.; Shankaran, S. A randomized placebo-controlled trial of massage therapy on the immune system of preterm infants. Pediatrics 2012, 130, e1549–e1558. [Google Scholar] [CrossRef]
- Courtiol, E.; Wilson, D.A.; Shah, R.; Sullivan, R.M.; Teixeira, C.M. Maternal Regulation of Pups’ Cortical Activity: Role of Serotonergic Signaling. eNeuro 2018, 5, ENEURO.0093-18.2018. [Google Scholar] [CrossRef]
- Kymre, I.G.; Bondas, T. Skin-to-skin care for dying preterm newborns and their parents—A phenomenological study from the perspective of NICU nurses. Scand J. Caring Sci. 2013, 27, 669–676. [Google Scholar] [CrossRef]
- Wyatt, J.; Hain, R. Redirecting Care: Compassionate Management of the Sick or Preterm Neonate at the End of Life. Children 2022, 9, 344. [Google Scholar] [CrossRef]
- Hignell, A.; Carlyle, K.; Bishop, C.; Murphy, M.; Valenzano, T.; Turner, S.; Sgro, M. The Infant Cuddler Study: Evaluating the effectiveness of volunteer cuddling in infants with neonatal abstinence syndrome. Paediatr. Child Health 2019, 25, 414–418. [Google Scholar] [CrossRef]
- Cassileth, B.R.; Vickers, A.J. Massage therapy for symptom control: Outcome study at a major cancer center. J. Pain Symptom Manag. 2004, 28, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Müller-Oerlinghausen, B.; Eggart, M. Touch Research-Quo Vadis? A Plea for High-Quality Clinical Trials. Brain Sci. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Kopf, D. Massage and touch-based therapy: Clinical evidence, neurobiology and applications in older patients with psychiatric symptoms. Z. Gerontol. Geriatr. 2021, 54, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Field, T.M. Social touch, CT touch and massage therapy: A narrative review. Dev. Rev. 2019, 51, 123–145. [Google Scholar] [CrossRef]


{kind=link}
| Disorder | Causes | Affected Fibers | Perception of Touch CT Related | Perception of Touch Not-CT Related |
|---|---|---|---|---|
| Anorexia nervosa | Multifactorial | Less pleasant than control | ||
| Rare sensory neuropaty | After infection (e.g., mononucleosis) | Absence of myelinated Aβ fibers on their large skin areas. Nevertheless, they have intact unmyelinated CT fibers | The ability to spatially localize CT stimulation is very poor | No perception |
| Disorder associated with a nerve growth factor beta (NGFB) | Gene mutation that determines the loss of NGFB function | Severe to moderate reduction of unmyelinated C fibers and a moderate reduction of thinly myelinated Aδ fibers | The pleasantness ratings of these patients were lower and the rating pattern across the different velocities (0.3–30 cm/s) deviated from the typical and normal rating pattern, therefore to the typical inverted U-shaped curve, correlated with CT discharge across velocities | |
| Psychiatric disorder | Multifactorial | Less pleasant than healthy subjects | ||
| Autism spectrum | Multifactorial | Aberrant behaviour (defensiveness); Reduction of the central network involved in CT related | Impaired; Increase of the central network involved in discriminative touch |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grandi, L.C.; Bruni, S. Social Touch: Its Mirror-like Responses and Implications in Neurological and Psychiatric Diseases. NeuroSci 2023, 4, 118-133. https://doi.org/10.3390/neurosci4020012
Grandi LC, Bruni S. Social Touch: Its Mirror-like Responses and Implications in Neurological and Psychiatric Diseases. NeuroSci. 2023; 4(2):118-133. https://doi.org/10.3390/neurosci4020012
Chicago/Turabian StyleGrandi, Laura Clara, and Stefania Bruni. 2023. "Social Touch: Its Mirror-like Responses and Implications in Neurological and Psychiatric Diseases" NeuroSci 4, no. 2: 118-133. https://doi.org/10.3390/neurosci4020012
APA StyleGrandi, L. C., & Bruni, S. (2023). Social Touch: Its Mirror-like Responses and Implications in Neurological and Psychiatric Diseases. NeuroSci, 4(2), 118-133. https://doi.org/10.3390/neurosci4020012