1. Introduction
Oxidative stress is a physiological phenomenon that occurs when there is a dramatic increase in the amount of cellular oxidizing equivalents and a concomitant decrease in cellular reducing equivalents. While oxidative stress has many origins, a common mechanism by which it occurs is through the generation/overproduction of reactive oxygen species (ROSs), including oxygen free radicals [
1,
2]. Common ROSs include hydrogen peroxide, singlet oxygen, superoxide anions, and hydroxyl radicals [
3]. Although ROSs produced from O
2 are needed for signaling reactions within the body, when cellular radicals accumulate, toxicity and inflammation can occur [
1]. ROS production and oxidative stress are known to increase with many environmental/chemical exposures, including pollutants, such as traffic exhaust emission, tobacco smoke, alcohol, or high levels of radiation [
3,
4]. Interestingly, multiple diseases, such as Alzheimer’s and Parkinson’s Disease, diabetes, and autoimmune disorders have also been linked to oxidative stress [
5,
6]. Under oxidative stress conditions, cellular function is often affected. As such, our study initiative was to determine whether sperm cell function and/or parameters correlate with alterations in cellular redox environments. Furthering our understanding of factors like oxidative stress and redox imbalance could elucidate potential therapies that could impact a man’s fertility.
The small biothiol glutathione (GSH) and its oxidized form, glutathione disulfide (GSSG), represents the predominant, most robust redox couple in most cells and, as such, is generally predictive of overall cellular redox environments. Glutathione is present within all cells of the body, as it regulates many cellular activities, including proliferation, differentiation, and apoptosis [
2,
3]. Central to GSH function is its cysteine residue, which can undergo reversible oxidation and is involved in reduction–oxidation (redox) reactions at the cellular level, allowing it to be an extremely powerful and useful antioxidant in the body [
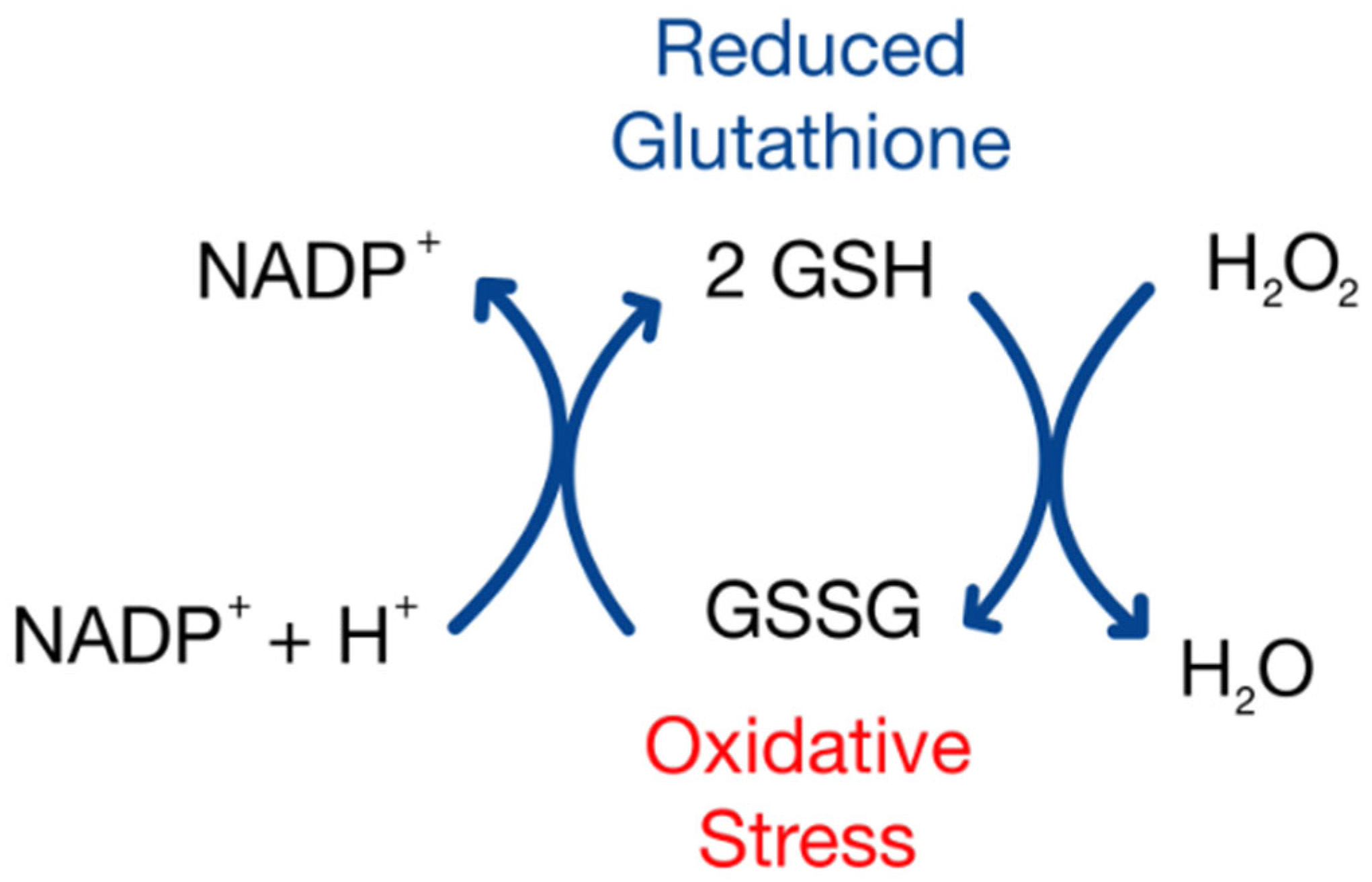
7]. GSH can be directly utilized in the detoxification of ROSs. For instance, hydrogen peroxide can be converted into water via the enzyme glutathione peroxidase (GPx), using two GSH molecules to yield a single GSSG [
8]. Levels of GSSG are mitigated through the actions of glutathione disulfide reductase, using NADPH to convert GSSG back into two GSH molecules, allowing the cycle of ROS detoxification to continue.
Using the Nernst equation, GSH and GSSG concentrations can indicate the cellular redox status, where comparably high concentrations of GSH to lower GSSG levels would suggest more reducing cellular environments. When GSSG increases comparatively, cellular environments become more oxidizing [
9]. Under normal conditions, the GSH:GSSG ratio can approach 100:1, but during periods of oxidative stress, that ratio may shift to 10:1 [
8]. While the GSH/GSSG ratio is often reported, perhaps a more accurate measure of cellular redox status is the cell’s redox potential (
Eh) [
10].
With reproductive capacity declining over the past 50 years, and infertility in men becoming more common [
11], we were interested to understand possible correlations between sperm GSH/GSSG
Eh and various measures of sperm quality. Since oxidized redox states are correlated with poor somatic cellular function, we expect that shifts to more oxidizing GSH/GSSG
Eh in sperm will correlate to lower sperm quality measures, with the converse being true with more reducing
Eh. Understanding how redox states influence sperm quality may serve as a point of therapeutic intervention to improve reproductive outcomes. Thus, the focus of this project is to evaluate GSH/GSSG
Eh in human sperm samples and correlate it with various measures of sperm quality. With greater knowledge of the correlation between redox potential and sperm function, a therapeutic target could be developed as an intervention for male infertility.
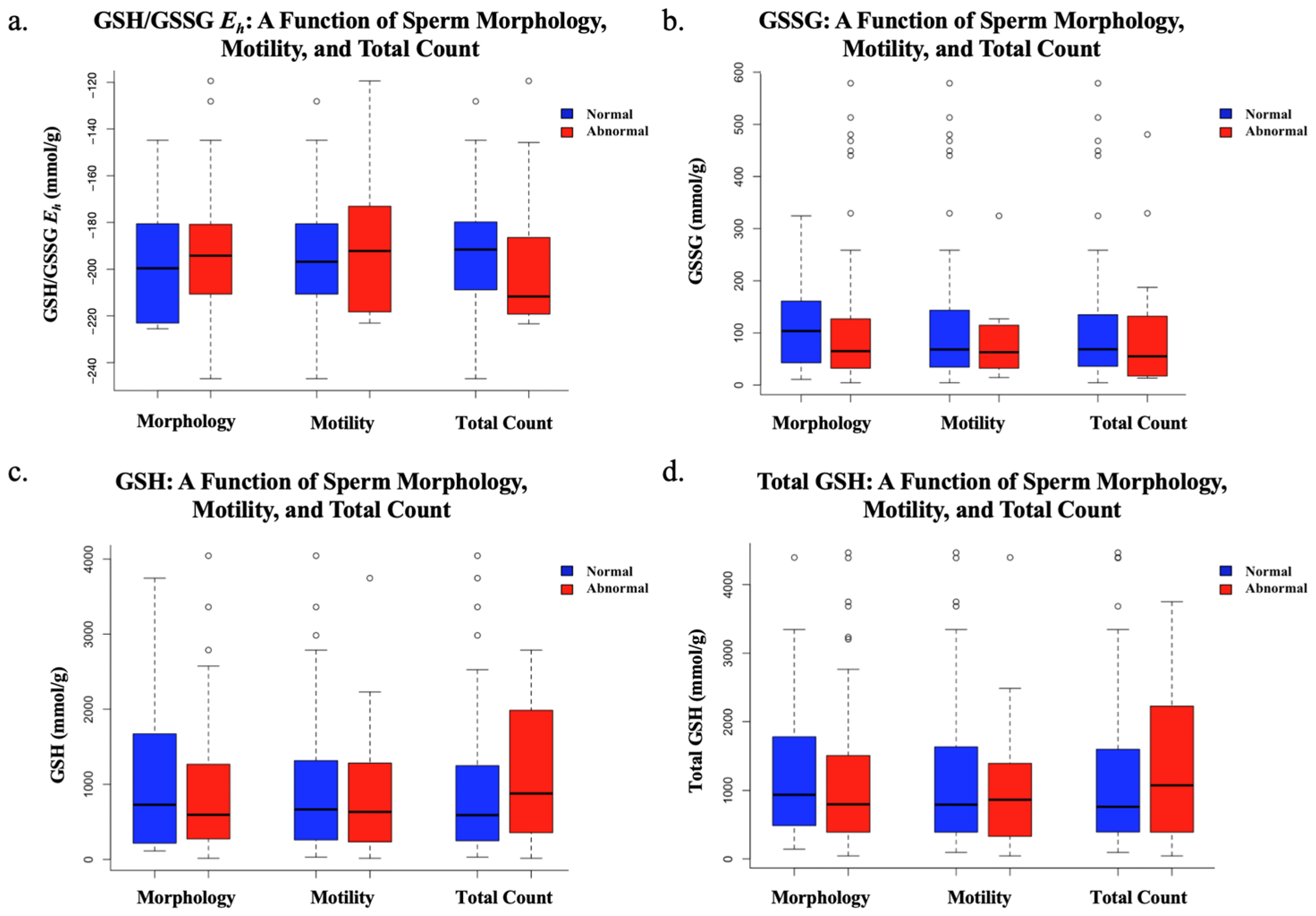
4. Discussion
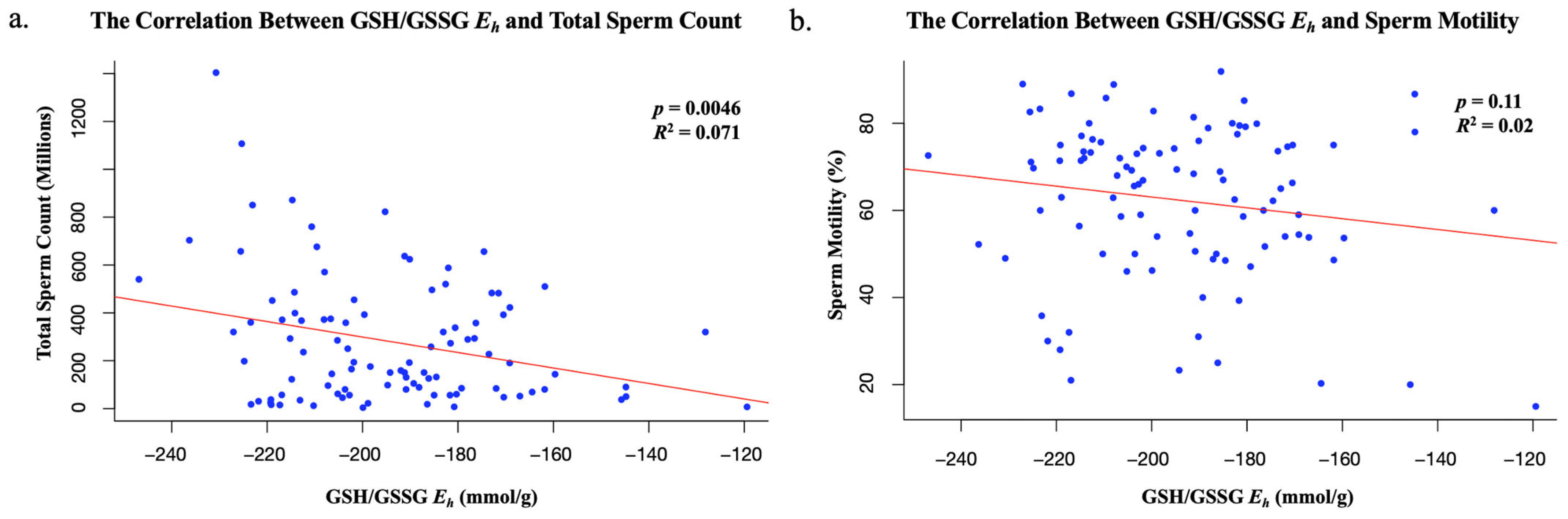
This study suggests that total sperm count decreases as redox potential increases. The inverse correlation between sperm total count and GSH/GSSG
Eh in semen analyses indicates that more oxidized sperm, with more positive redox potentials, may contribute to male infertility. Despite this finding being statistically significant, this study suggests that GSH/GSSG
Eh does not impact an individual’s total sperm motility or morphology. Additionally, while a statistically significant inverse correlation between GSH/GSSG
Eh and total sperm count was identified (
p = 0.0046), the relationship was relatively weak (
R2 = 0.071), indicating that only a small portion of the variability in sperm count could be explained by changes in the redox state. This suggests that other biological or environmental factors most likely play a more prominent role in determining sperm count, and that GSH/GSSG
Eh may not be a strong predictor in this context. This variation should be considered when interpreting these results, as using GSH/GSSG
Eh is not the only predictor of redox potential, and past studies have found sperm parameters such as motility and viability to be highly sensitive to ROSs [
15,
16]. Future studies could explore additional indicators of redox status, including direct quantification of reactive oxygen species (ROSs). The evaluation of additional redox couples, such as cysteine/cystine (Cys/CySS), may also help to further elucidate the relationship between redox homeostasis and sperm function. Blinded research with larger sample sizes and a fertile (normal) control group is essential to confirm and strengthen this correlation. If validated, this correlation would provide a potential therapeutic target for intervention amongst patients who suffer from asthenozoospermia, as one could receive glutathione treatment to increase the levels of GSH, therefore shifting the glutathione redox couple to a more negative and reducing state. Studies have shown that glutathione has a protective effect on ROS-mediated DNA damage, and the protein can have a reversible effect on H
2O
2-induced cytotoxicity and ROS generation (
Figure 4) [
17]. This poses the idea that glutathione treatments could potentially reverse a decline in sperm count in men over the course of time, thus increasing one’s ability to conceive a viable embryo.
The average total sperm count has been found to have declined to around 62.3% in the past 45 years, with many possible causes contributing to this decline [
11]. Sperm counts since 2000 have decreased at a rate −2.64%/year compared to the rates in 1972 of −1.16%/year. Our study elucidates the idea that the redox state of sperm could be a contributor to this outcome. With a statistically significant correlation being made between redox state and total sperm count but not sperm motility, our results suggest that redox state is more closely associated with spermatogenesis than sperm function. Using plasma redox states as a marker of systemic redox states, studies have identified plasma redox couple shifts that occur during aging (ages 19–85), of which GSH/GSSG
Eh was a couple that was evaluated [
13]. The CyS/CySS redox couple is the primary redox couple in plasma, mostly being derived as a breakdown product from GSH/GSSG secreted from cells. Interestingly, CyS/CySS
Eh almost immediately becomes increasingly oxidized at a linear rate of nearly +0.16 mV/year throughout life. Conversely, the GSH/GSSG
Eh showed a completely different pattern, where redox states were relatively stable until age 45, after which it becomes more rapidly oxidized at a rate of +0.7 mV/year. Because these two redox couples are related but appear independently regulated, it suggests that the individual couples may have specified function and control of cellular activities. Additional studies have shown that total sperm counts become significantly lower starting at ages between 41 and 50 [
18], overlaying on periods where shifts in GSH/GSSG
Eh begin to become increasingly oxidized in humans. These results are highly suggestive of a GSH/GSSG centric, homeostatic redox mechanism to support normal spermatogenesis.
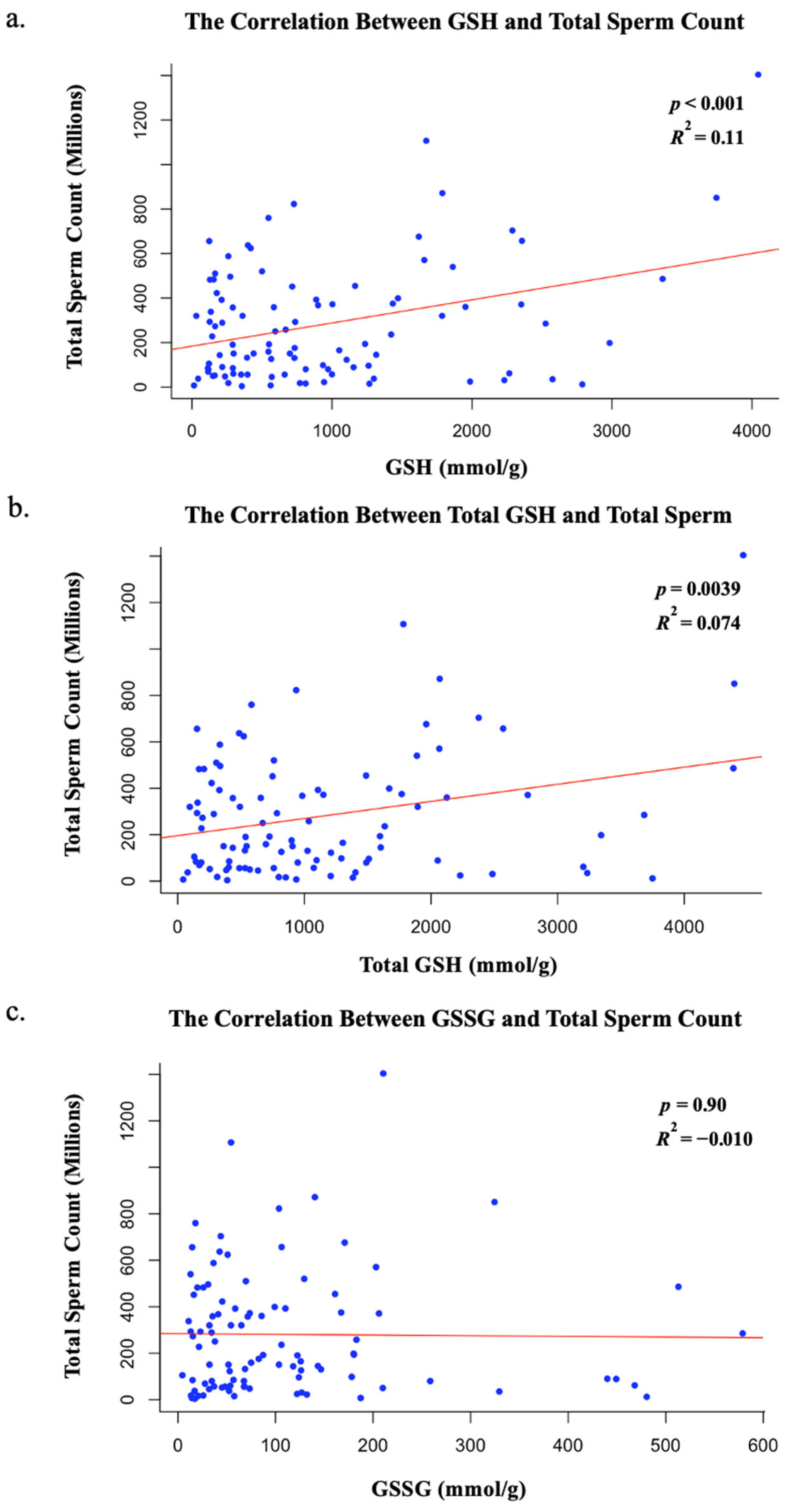
Intriguingly, our data show that GSH and total GSH levels, not GSSG levels, correlate with total sperm count. Thus, these data suggest that a decline in GSH accumulation or absence of reduced glutathione within sperm cells, and not an increase in GSSG accumulation, may be associated with decreased sperm counts in ejaculated sperm samples. However, because our measurements were performed on ejaculated sperm, the redox status primarily reflects the extracellular environment of seminal plasma, not the intratesticular environment during spermatogenesis. While reduced antioxidants like GSH significantly contribute to ROS detoxification, we cannot conclude from our data that decreased GSH levels directly impair spermatogenesis. Nevertheless, Sertoli cells actively synthesize GSH, and adequate antioxidant protection from GSH may be required within the seminiferous epithelium for proper sperm development and maturation [
19]. Future studies evaluating redox balance within testicular tissue itself are needed to clarify this relationship. Thus, while our findings reveal a correlation between seminal plasma GSH levels and sperm count, further mechanistic research is required to determine whether these associations extend to redox regulation during spermatogenesis. Alternatively, lowered levels of GSH and a more oxidizing redox state in the seminal environment may be reflective of broader oxidative stress that could contribute to impaired spermatogenesis, potentially resulting in decreased total sperm count, as observed in our cohort.
With an increase in the rate of total sperm count decline, it is difficult to determine a single factor that is responsible, but there are likely many environmental influences that are contributory, like pollution, alcohol consumption, or medications [
20], many of which may promote oxidative stress and alterations to lower GSH levels. Numerous studies have shown that specific environmental/chemical influences that promote oxidative stress and injury are positively correlated with poor sperm quality and a decrease in positive reproductive outcomes. For example, cigarettes contain several chemicals that promote free radical production and oxidative injury, promoting changes to cellular redox states, including reproductive toxicants such as cadmium and oxidants such as various peroxides. Generally speaking, sperm concentrations in smokers are lower than in non-smokers [
21]. In terms of smoking and infertility, human studies showed that infertile, non-smoking men had sperm concentrations that were decreased by nearly 60% (25.5 × 10
6/mL vs. 62.8 × 10
6/mL in controls), but in infertile, smoking men, sperm concentrations dropped even more, by 73% (17.0 × 10
6/mL). Interestingly, similar trends were noted when comparing spermatozoa GSH levels, with infertile, non-smoking samples showing significantly higher levels (35.4 pmol/10
6 spermatozoa) than infertile, smoking samples (31.4 pmol/10
6 spermatozoa). Additionally, markers of oxidative-stress-induced damage, such as malondialdehyde (MDA) and protein carbonyls, measures of lipid peroxidation and protein oxidation, respectively, and ROS itself, were all significantly higher in the spermatozoa from infertile, smoking men than those from infertile, non-smoking men [
22].
Other studies on infertile human males also show similar trends, where ROS levels in smokers are significantly higher in smokers vs. non-smokers. However, when infertile non-smokers are compared to fertile men (normal donors), ROS levels are still significantly greater in infertile non-smokers [
21], suggesting that smoking is not the sole rationale for ROS differences, but rather the observed ROS increase is connected to infertility in another way. Interestingly, in this same study, data show that infertile smokers have a decreased sperm concentration by approximately 40% from normal donors, but infertile non-smokers have an even greater decrease, a 57% decrease in sperm concentrations [
21]. These data further support the possibility that sperm counts are, at least in part, correlated to changes in ROS availability and potential oxidation. DNA damage was higher in infertile smokers (
p = 0.04) but not higher in infertile non-smokers (
p = 0.06), even though comparisons between groups show that the amount of damage was essentially the same [
21]. Glutathione addition to collected sperm improved a variety of parameters related to sperm quality, especially related to kinematic measures [
23]. ATP levels were increased with supplementation, while ROS availability was significantly decreased. In vivo GSH supplementation (600 mg/day IM) for 60 days in infertile men also led to similar improvements on kinematic measurements, significantly increasing total motility, linear motility, and velocity [
24]. This approach did not affect the total sperm concentration, however.
While not performed specifically in humans, mice with Wilson’s-disease-associated oligospermia that received GSH supplementation showed a significant increase in sperm counts. GSH as a supplement has already shown to improve the sperm count in mice while reducing ROS availability and diminishing MDA levels and occur concomitantly with corrections in mitochondrial health and a reduction in testicular apoptosis [
25]. While these studies highlight the direct support for GSH in positive male reproductive outcomes, they do not directly address the role of redox states’ contributions to male infertility.
Our primary finding related GSH/GSSG
Eh to changes in sperm concentrations in human subjects. Glutathione redox states have been highly studied in regard to cellular proliferation. Early work showed that fibroblasts, both normal, wild-type fibroblasts, which display contact inhibition, and fibrosarcoma cells, which do not exhibit contact inhibition, exhibited very reducing GSH/GSSG
Eh at low confluency, where proliferation rates were relatively high. As confluency increased, normal fibroblast proliferation decreased, which coincided with oxidation in GSH redox states, but fibrosarcoma cells, which continued constant growth, did not show GSH redox state oxidation but rather remained unchanged, suggesting that proliferating cells prefer a reducing intracellular redox environment. Using wild-type fibroblasts, GSH/GSSG
Eh was pharmacologically oxidized (+24 mV), and cells demonstrated a decrease in cellular proliferation by 54% compared to untreated cells [
26]. Inversely, pharmacologically reducing GSH/GSSG
Eh (−10 mV) in fibroblasts resulted in increased proliferation (26% increase from control). With more reducing systems, cells appear to proliferate at a much higher rate. While spermatozoa are not mitotic, precursors are actively dividing and proliferating. Lowered GSH and more oxidizing GSH/GSSG
Eh at this level may cause a low sperm yield, reflected in lower sperm counts in the semen.
Our study shows that a more reduced redox state correlates highly with higher sperm counts. More oxidized GSH/GSSG Eh is associated with much lower counts, but, interestingly, shifts to more oxidizing GSH/GSSG Eh are not driven by increases in GSSG levels, which did not show a significant correlation, but rather strongly correlated to decreased GSH concentrations. As such, a decrease in total GSH in sperm, also correlated with lowered sperm counts, is reflective of a loss of the reduced form of glutathione, GSH. Direct supplemental GSH or other pharmacological means to increase GSH levels to increase sperm counts could be a potential approach to increase fertility. Clearly, further work is required to better understand meaningful therapies regarding GSH/GSSG Eh regulatory control of sperm concentrations in men, but these findings do support a feasible and potential point of intervention.
In conclusion, we believe our study provides data acknowledging that the GSH/GSSG Eh shift to a more positive, oxidizing redox potential in sperm drives lower sperm counts in men. Alternatively, more reduced GSH/GSSG Eh promotes increased sperm counts in men and is indicative of less oxidative stress and a healthier cellular environment. Thus, we infer that preconception lifestyle, including factors such as smoking, diet, and environmental exposures, can all contribute to cellular GSH/GSSG Eh, and ultimately sperm count or production. A greater understanding of both the culminating reasons behind the depletion of GSH levels in sperm, as well as more data pertaining to sperm production under oxidative stress, is needed to develop redox-based strategies to improve infertility.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}