
Minor Contribution by Biomineralizing Phytoplankton to Surface Ocean Biomineral Pools in the Late Stratified Period


Abstract
:
1. Introduction
2. Materials and Methods
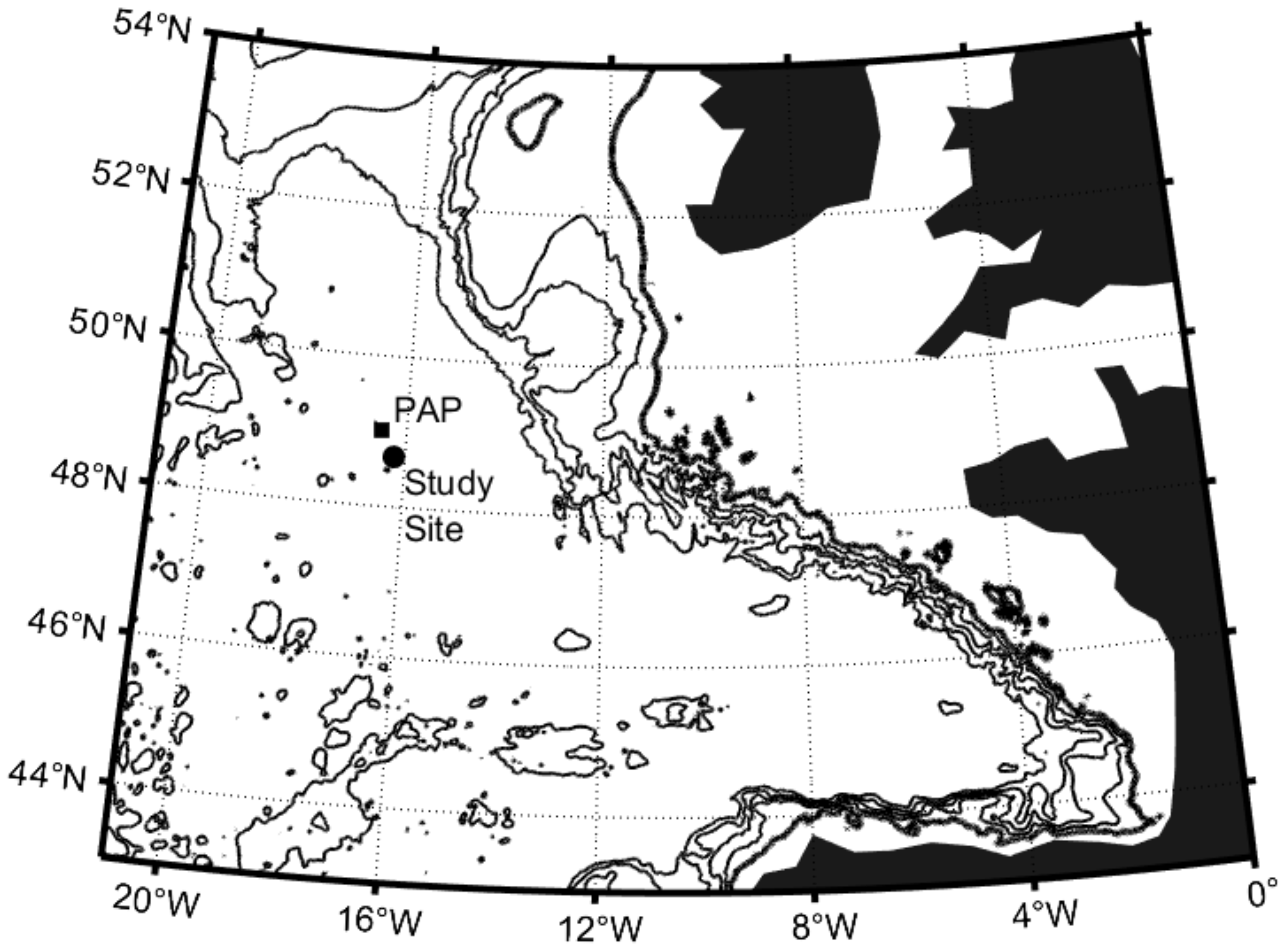
2.1. Sampling and Environmental Characterisation
2.2. Irradiance Conditions
2.3. Nutrients
2.4. Total and Size-Fractionated Chlorophyll-a
2.5. Biogenic Silica
2.6. Particulate Inorganic Carbon
2.7. Phytoplankton Community
2.8. Biomineral Budgets
3. Results
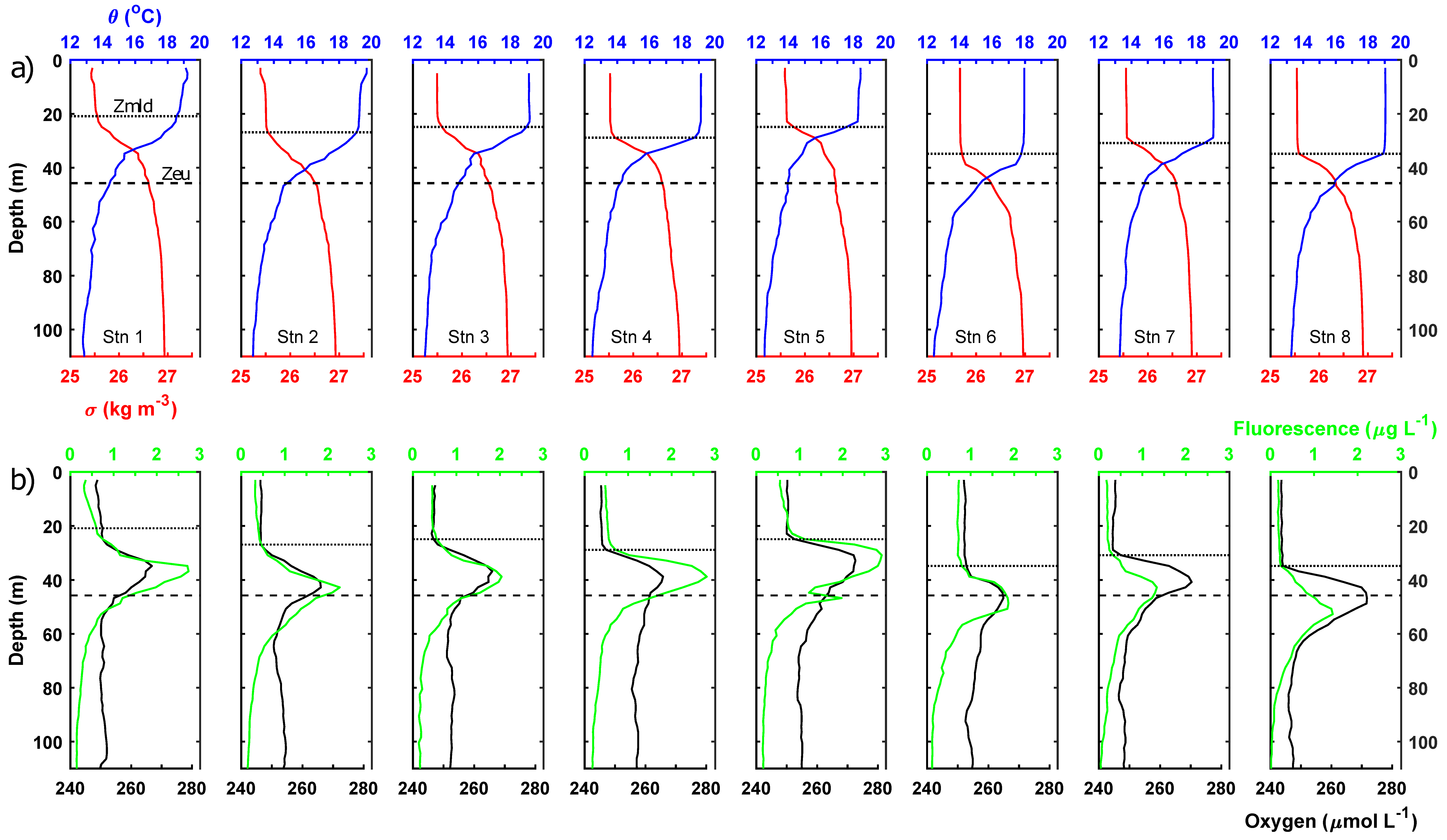
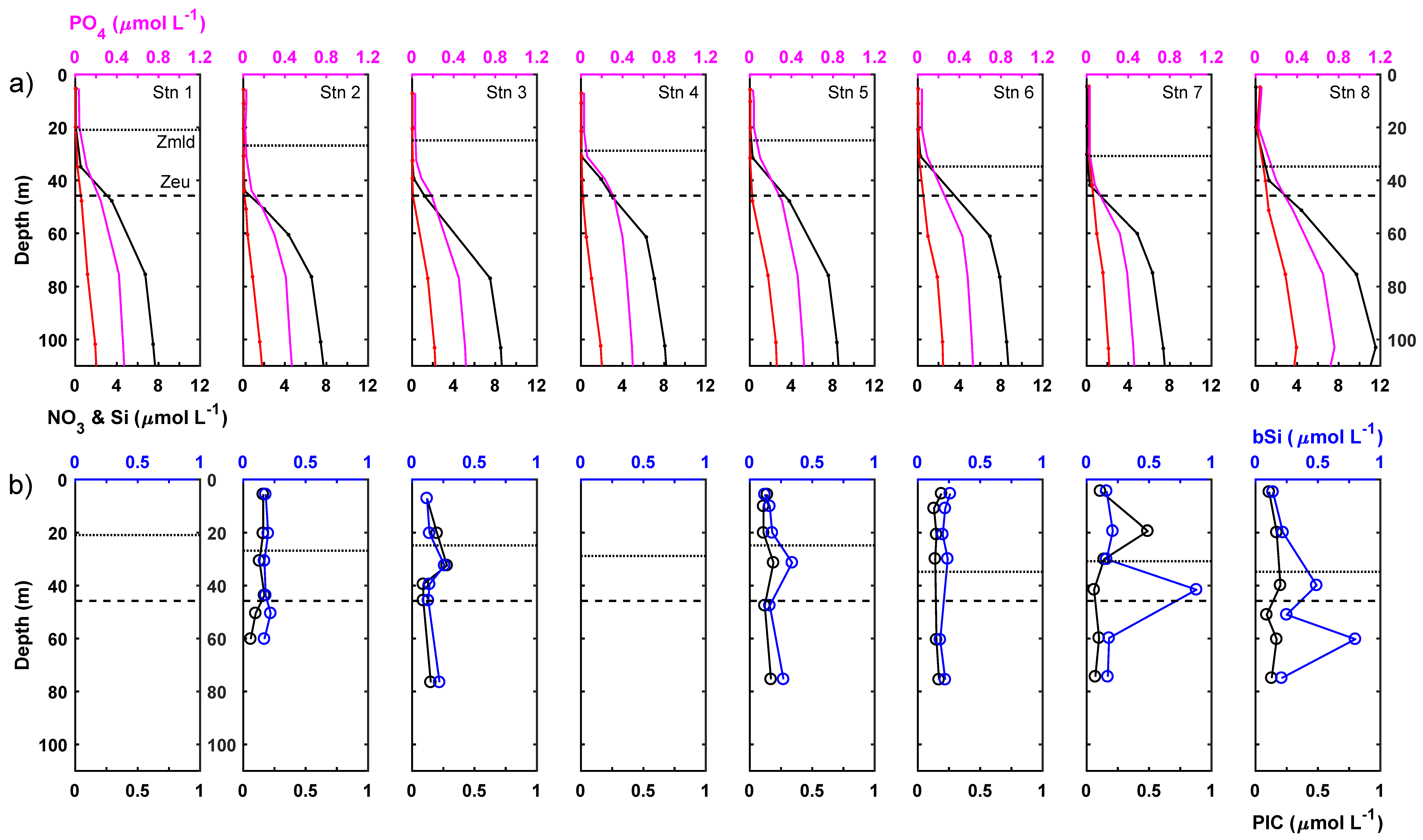
3.1. Mixed Layer Conditions
3.2. Lower Euphotic Zone
3.3. Sub-Euphotic Zone
3.4. Integrated Results
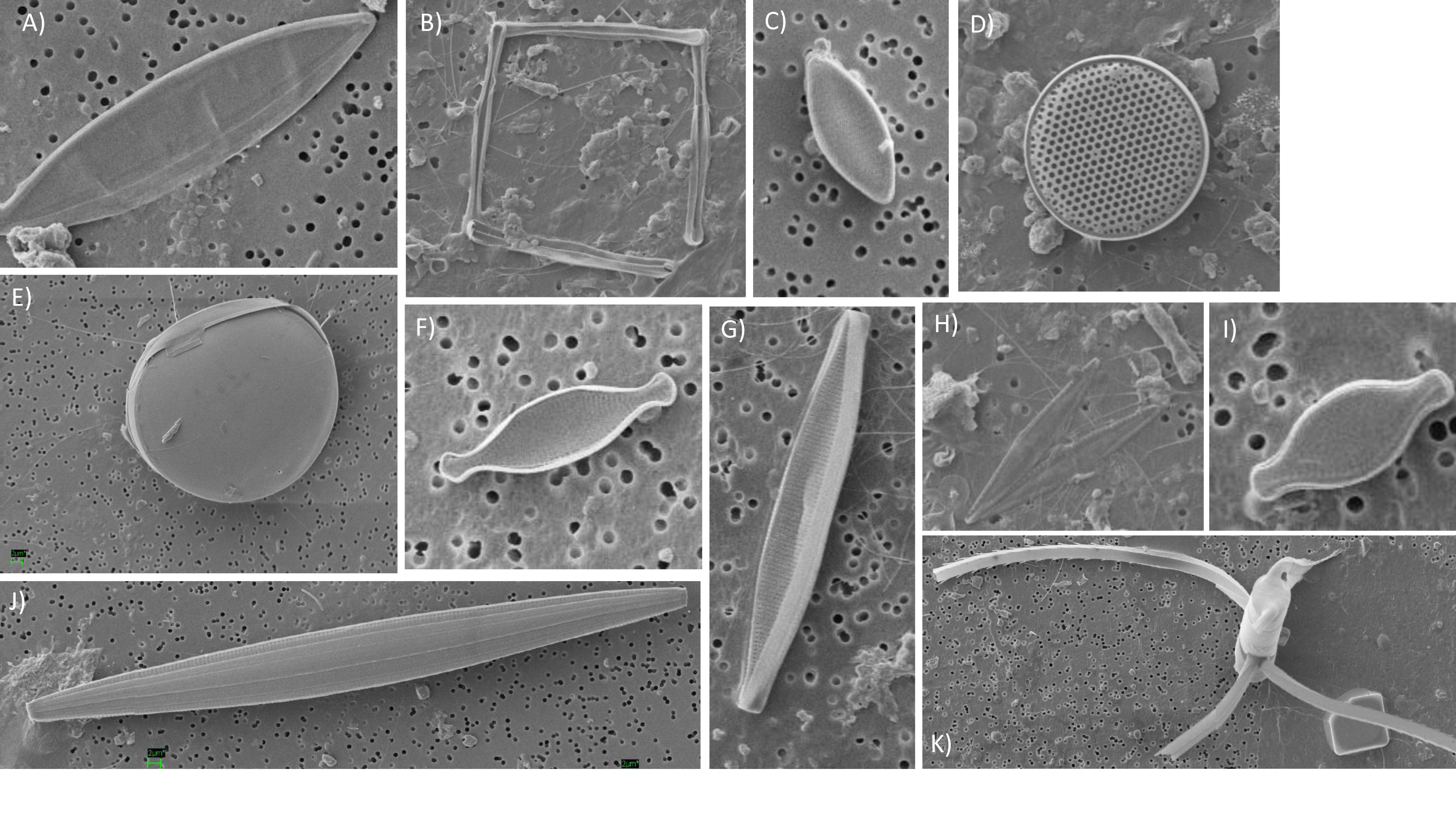
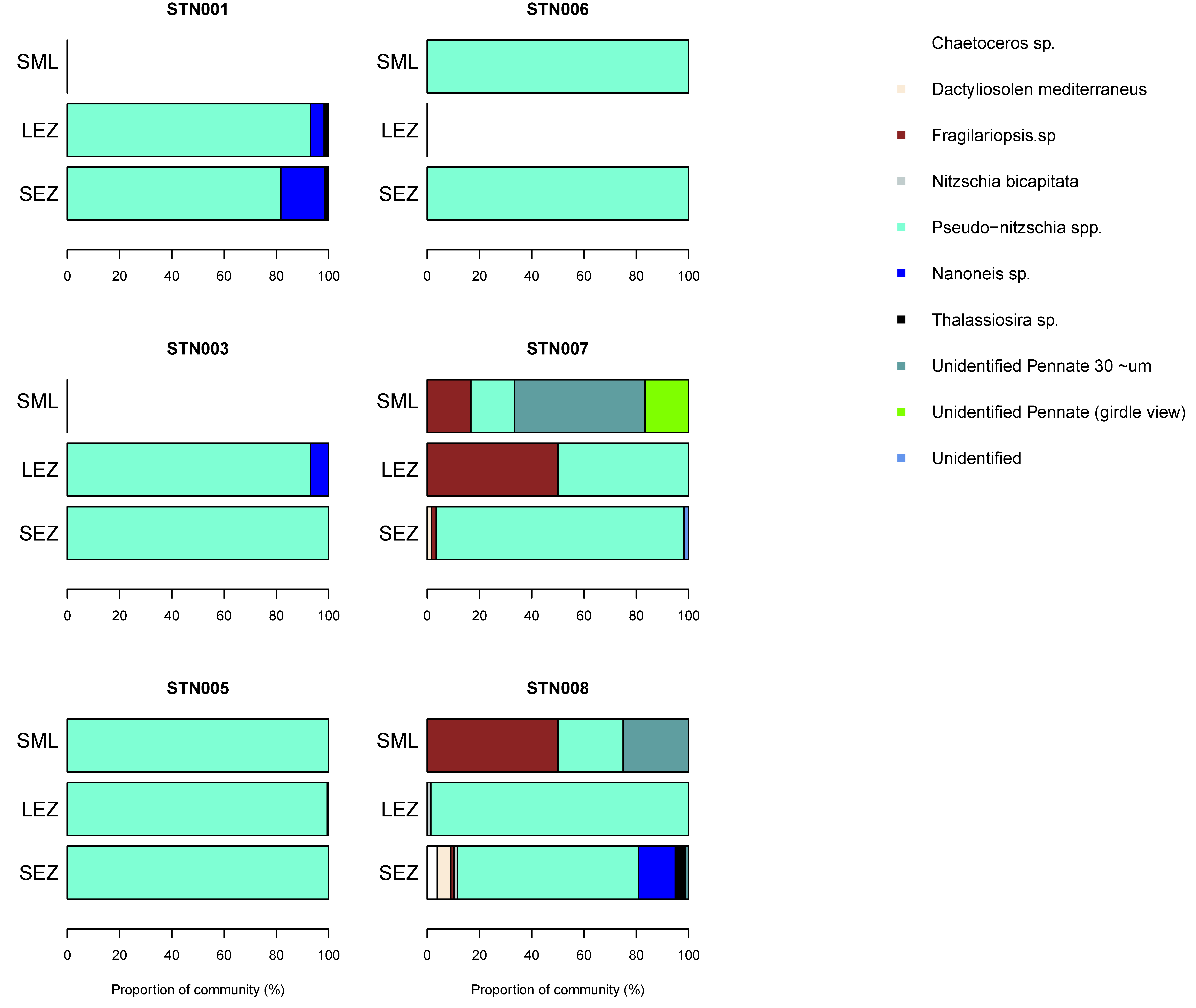
3.5. Phytoplankton Community
3.6. PIC Budget
3.7. bSi Budget
4. Discussion
Implications for Autumn Export Events
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klass, C.; Archer, D.E. Association of sinking organic matter with various types of mineral ballast in the deep sea: Implications for the rain ratio. Glob. Biogeochem. Cycles 2002, 16, 1116. [Google Scholar] [CrossRef]
- Passow, U.; De La Rocha, C.L. Accumulation of mineral ballast on organic aggregates. Glob. Biogeochem. Cycles 2006, 20, GB1013. [Google Scholar] [CrossRef] [Green Version]
- Sanders, R.; Morris, P.J.; Poulton, A.J.; Stinchcombe, M.C.; Charalampopoulou, A.; Lucas, M.I.; Thomalla, S.J. Does a ballast effect occur in the surface ocean? Geophys. Res. Lett. 2010, 37, L08602. [Google Scholar] [CrossRef] [Green Version]
- Le Moigne, F.A.C.; Sanders, R.J.; Villa-Alfageme, M.; Martin, A.P.; Pabortsava, K.; Planquette, H.; Morris, P.J.; Thomalla, S.J. On the proportion of ballast versus non-ballast associated carbon export in the surface ocean. Geophys. Res. Lett. 2012, 39, L15610. [Google Scholar] [CrossRef] [Green Version]
- Paasche, E.; Ostergren, I. The annual cycle of plankton diatom growth and silica production in the inner Oslofjord. Limnol. Oceanogr. 1980, 25, 481–494. [Google Scholar]
- Leblanc, K.; Leynaert, A.; Fernandez, I.C.; Rimmelin, P.; Moutin, T.; Raimbault, P.; Ras, J.; Queguiner, B. A seasonal study of diatom dynamics in the North Atlantic during the POMME experiment (2001): Evidence for Si limitation of the spring bloom. J. Geophys. Res. 2005, 110, C07S14. [Google Scholar] [CrossRef] [Green Version]
- Terrats, L.; Claustre, H.; Cornec, M.; Magin, A.; Neukermans, G. Detection of Coccolithophore Blooms with BioGeoChemical-Argo Floats. Geophys. Res. Lett. 2020, 47, e2020GL090559. [Google Scholar] [CrossRef]
- O’Brien, C.J.; Peloquin, J.A.; Vogt, M.; Heinle, M.; Gruber, N.; Ajani, P.; Andruleit, H.; Arıstegui, J.; Beaufort, L.; Estrada, M.; et al. Global marine plankton functional type biomass distributions: Coccolithophores. Earth Syst. Sci. Data 2013, 5, 259–276. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, K.; Arıstegui, J.; Armand, L.; Assmy, P.; Beker, B.; Bode, A.; Breton, E.; Cornet, V.; Gibson, J.; Gosselin, M.-P.; et al. A global diatom database—abundance, biovolume and bimass in the world ocean. Earth Syst. Sci. Data 2012, 4, 149–165. [Google Scholar] [CrossRef] [Green Version]
- Francois, R.; Honjo, S.; Krishfield, R.; Manganini, S. Factors controlling the flux of organic carbon to the bathypelagic zone of the ocean. Glob. Biogeochem. Cycles 2002, 16, 1087. [Google Scholar] [CrossRef] [Green Version]
- Rings, A.; Lücke, A.; Schleser, G.H. A new method for the quantitative separation of diatom frustules from lake sediments. Limnol. Oceanogr. Methods 2004, 2, 25–34. [Google Scholar] [CrossRef]
- Hopkins, J.; Henson, S.A.; Painter, S.C.; Tyrrell, T.; Poulton, A.J. Phenological characteristics of global coccolithophore blooms. Glob. Biogeochem. Cycles 2015, 29, 239–253. [Google Scholar] [CrossRef] [Green Version]
- Kemp, A.E.S.; Villareal, T.A. The case of the diatoms and the muddled mandalas: Time to recognize diatom adaptations to stratified waters. Prog. Oceanogr. 2018, 167, 138–149. [Google Scholar] [CrossRef]
- Kemp, A.E.S.; Pike, J.; Pearce, R.B.; Lange, C.B. The “fall dump”—A new pespective on the role of a "shade flora" in the annual cycle of diatom production and export flux. Deep Sea Res. Part II 2000, 47, 2129–2154. [Google Scholar] [CrossRef]
- Caracciolo, M.; Beaigrand, G.; Helaouet, P.; Gevaert, F.; Edwards, M.; Lizon, F.; Kleparski, L.; Goberville, E. Annual phytoplankton succession results from niche-environment interaction. J. Plankton Res. 2020, 43, 85–102. [Google Scholar] [CrossRef]
- Brun, P.; Vogt, M.; Payne, M.R.; Gruber, N.; O’Brien, C.J.; Buitenhuis, E.T.; Le Quere, C.; Leblanc, K.; Luo, Y.-W. Ecological niches of open ocean phytoplankton taxa. Limnol. Oceanogr. 2015, 60, 1020–1038. [Google Scholar] [CrossRef] [Green Version]
- Boyd, P.W.; Strzepek, R.; Fu, F.; Hutchins, D.A. Environmental control of open-ocean phytoplankton groups: Now and in the future. Limnol. Oceanogr. 2010, 55, 1353–1376. [Google Scholar] [CrossRef]
- Latasa, M.; Cabello, A.M.; Moran, X.A.G.; Massana, R.; Scharek, R. Distribution of phytoplankton groups within the deep chlorophyll maximum. Limnol. Oceanogr. 2017, 62, 665–685. [Google Scholar] [CrossRef] [Green Version]
- Painter, S.C.; Finlay, M.; Hemsley, V.S.; Martin, A.P. Seasonality, phytoplankton succession and the biogeochemical impacts of an autumn storm in the northeast Atlantic Ocean. Prog. Oceanogr. 2016, 142, 72–104. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, K.; Hare, C.E.; Feng, Y.; Berg, G.M.; DiTullio, G.R.; Neely, A.; Benner, I.; Sprengel, C.; Beck, A.; Sanudo-Wilhelmy, S.A.; et al. Distribution of calcifying and silicifying phytoplankton in relation to environmental and biogeochemical parameters during the late stages of the 2005 North East Atlantic Spring Bloom. Biogeosciences 2009, 6, 2155–2179. [Google Scholar]
- Gordon, H.R.; Bynton, G.C.; Balch, W.M.; Groom, S.B.; Harbour, D.S.; Smyth, T.J. Retrieval of coccolithophore calcite concentration from SeaWiFS imagery. Geophys. Res. Lett. 2001, 28, 1587–1590. [Google Scholar] [CrossRef]
- Balch, W.M.; Gordon, H.R.; Bowler, B.C.; Drapeau, D.T.; Booth, E.S. Calcium carbonate measurements in the surface global ocean based on Moderate-Resolution Imaging Spectroradiometer data. J. Geophys. Res. 2005, 110, C07001. [Google Scholar] [CrossRef]
- Friedland, K.D.; Record, N.R.; Asch, R.G.; Kristiansen, T.; Saba, V.S.; Drinkwater, K.F.; Henson, S.; Leaf, R.T.; Morse, R.E.; Johns, D.G.; et al. Seasonal phytoplankton blooms in the North. Atlantic linked to the overwintering strategies of copepods. Elem. Sci. Anthr. 2016, 4, 000099. [Google Scholar] [CrossRef]
- Martinez, E.; Antoine, D.; D’Ortenzio, F.; de Boyer Montegut, C. Phytoplankton spring and fall blooms in the North Atlantic in the 1980s and 2000s. J. Geophys. Res. 2011, 116, C11. [Google Scholar] [CrossRef] [Green Version]
- Lampitt, R.S.; Billet, D.S.M.; Martin, A.P. The sustained observatory over the Porcupine Abyssal Plain (PAP): Insights from time series observations and process studies. Deep. Sea Res. Part II 2010, 57, 1267–1271. [Google Scholar] [CrossRef]
- Hartman, S.E.; Bett, B.J.; Durden, J.M.; Henson, S.A.; Iversen, M.; Jeffreys, R.M.; Horton, T.; Lampitt, R.; Gates, A.R. Enduring science: Three decades of observing the Northeast Atlantic from the Porcupine Abyssal Plain Sustained Observatory (PAP-SO). Prog. Oceanogr. 2021, 191, 102508. [Google Scholar] [CrossRef]
- Buckingham, C.E.; Naveira-Garabato, A.C.; Thompson, A.F.; Brannigan, L.; Lazar, A.; Marshall, D.P.; Nurser, A.J.G.; Damerell, G.; Heywood, K.J.; Belcher, S.E. Seasonality of submesoscale flows in the ocean surface boundary layer. Geophys. Res. Lett. 2016, 43, 2118–2126. [Google Scholar] [CrossRef] [Green Version]
- Damerell, G.M.; Heywood, K.J.; Calvert, D.; Grant, A.L.M.; Bell, M.J.; Belcher, S.E. A comparison of five surface mixed layer models with a year of observations in the North Atlantic. Prog. Oceanogr. 2020, 187, 102316. [Google Scholar] [CrossRef]
- Naveira-Garabato, A. Cruise Report: RRS James Cook Cruise 090, 30 August–17 September 2013. Ocean Surface Mixing, Ocean Sub-Mesoscale Interaction Study (OSMOSIS); National Oceanography Centre: Southampton, UK, 2013; 111p. [Google Scholar]
- de Boyer Montegut, C.; Madec, G.; Fischer, A.S.; Lazar, A.; Ludicone, D. Mixed layer depth over the global ocean: An. examination of profile data and a profile-based climatology. J. Geophys. Res. 2004, 109, C12003. [Google Scholar] [CrossRef]
- Lee, Z.-P.; Du, K.-P.; Arnone, R. A model for the diffuse attenuation coefficient of downwelling irradiance. J. Geophys. Res. 2005, 110. [Google Scholar] [CrossRef]
- Kirk, J.T.O. Light and Photosynthesis in Aquatic Ecosystem, 3rd ed.; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Hydes, D.J.; Aoyama, M.; Aminot, A.; Bakker, K.; Becker, S.; Coverly, S.; Daniel, A.; Dickson, A.G.; Grosso, O.; Kerouel, R.; et al. Determination of dissolved nutrients (N, P, Si) in seawater with high precision and inter-comparability using gas-segmented continuous flow analysers. In The GO-Ship Repeat Hydrography Manual: A Collection of Expert Reports and Guidelines; Hood, E.M., Sabine, C.L., Sloyan, B.M., Eds.; IOCCP Report No. 14, ICPO Publication Series No. 134; ICPO: Paris, France, 2010; pp. 1–87. Available online: http://www.ioccp.org/images/06Nutrients/Hydes_et_al_Nutrients.pdf (accessed on 16 July 2021).
- Welschmeyer, N.A. Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and phaeopigments. Limnol. Oceanogr. 1994, 39, 1985–1992. [Google Scholar] [CrossRef]
- Ragueneau, O.; Treguer, P. Determination of biogenic silica in coastal waters: Applicability and limits of the alkaline digestion method. Marine Chem. 1994, 45, 43–51. [Google Scholar] [CrossRef]
- Green, D.R.H.; Cooper, M.J.; German, C.R.; Wilson, P.A. Optimization of an inductively coupled plasma—Optical emission spectrometry method for the rapid determination of high-precision Mg/Ca and Sr/Ca in foraminiferal calcite. Geochem. Geophys. Geosyst. 2003, 4, 8404. [Google Scholar] [CrossRef]
- Cros, L.; Fortuno, J.-M. Atlas of Northwestern Mediterranean coccolithophores. Sci. Mar. 2002, 66, 1–186. [Google Scholar] [CrossRef] [Green Version]
- Young, J.; Geisen, M.; Cros, L.; Kleijne, A.; Sprengel, C.; Probert, I.; Østergaard, J. A guide to extant coccolithophore taxonomy. J. Nannoplankton Res. Spec. Issue 2003, 1, 1–125. [Google Scholar]
- Nannotax3. Available online: www.mikrotax.org/Nannotax3/ (accessed on 16 July 2021).
- Charalampopoulou, A.; Poulton, A.J.; Tyrrell, T.; Lucas, M.I. Irradiance and pH affect coccolithophore community composition on a transect between the North Sea and the Arctic Ocean. Mar. Ecol. Prog. Ser. 2011, 431, 25–43. [Google Scholar] [CrossRef] [Green Version]
- Poulton, A.J.; Holligan, P.M.; Charalampopoulou, A.; Adey, T.R. Coccolithophore ecology in the tropical and subtropical Atlantic Ocean: New perspectives from the Atlantic meridional transect (AMT) programme. Prog. Oceanogr. 2017, 158, 150–170. [Google Scholar] [CrossRef]
- Hasle, G.R.; Syvertsen, E.E. Marine diatoms. In Identifying Marine Phytoplankton; Tomas, C.R., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 5–385. [Google Scholar]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990; p. 747. [Google Scholar]
- Steidinger, K.A.; Tangen, K. Dinoflagellates. In Identifying Marine Phytoplankton; Tomas, C.R., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 387–584. [Google Scholar]
- Daniels, C.J.; Poulton, A.J.; Young, J.R.; Esposito, M.; Humphreys, M.P.; Ribas-Ribas, M.; Tynan, E.; Tyrrell, T. Species-specific calcite production reveals Coccolithus pelagicus as the key calcifier in the Arctic Ocean. Mar. Ecol. Prog. Ser. 2016, 555, 29–47. [Google Scholar] [CrossRef] [Green Version]
- Young, J.R.; Ziveri, P. Calculation of coccolith volume and its use in calibration of carbonate flux estimates. Deep Sea Res. Part II 2000, 47, 1679–1700. [Google Scholar] [CrossRef]
- Yang, T.-N.; Wei, K.-Y. How many coccoliths are there in a coccosphere of the extant coccolithophorids? A compilation. J. Nannoplankton Res. 2003, 25, 7–15. [Google Scholar]
- Brzezinski, M.A. The Si:C:N ratio of marine diatoms: Interspecific variability and the effect of some environmental variables. J. Phycol. 1985, 21, 347–357. [Google Scholar] [CrossRef]
- Conley, D.J.; Kilham, S.S.; Theriot, E. Differences in silica content between marine and freshwater diatoms. Limnol. Oceanogr. 1989, 34, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Boutorh, J.; Moriceau, B.; Gallinari, M.; Ragueneau, O.; Bucciarelli, E. Effect of trace metal-limited growth on the postmortem dissolution of the marine diatom Pseudo-nitzschia delicatissima. Glob. Biogeochem. Cycles 2016, 30, 57–69. [Google Scholar] [CrossRef]
- Hillebrand, H.; Durselen, C.-D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculations for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Hemsley, V.S.; Smyth, T.J.; Martin, A.P.; Frajka-Williams, E.; Thompson, A.F.; Damerell, G.; Painter, S.C. Estimating oceanic primary production using vertical irradiance and chlorophyll profiles from ocean gliders in the North Atlantic. Environ. Sci. Technol. 2015, 49, 11612–11621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longhurst, A. Seasonal cycles of pelagic production and consumption. Prog. Oceanogr. 1995, 36, 77–167. [Google Scholar] [CrossRef]
- Koeve, W. Wintertime nutrients in the North Atlantic—New approaches and implications for new production estimates. Mar. Chem. 2001, 74, 245–260. [Google Scholar] [CrossRef]
- Hartman, S.E.; Larkin, K.E.; Lampitt, R.S.; Lankhorst, M.; Hydes, D.J. Seasonal and inter-annual biogeochemical variations in the Porcupine Abyssal Plain 2003–2005 associated with winter mixing and surface circulation. Deep Sea Res. Part II 2010, 57, 1303–1312. [Google Scholar] [CrossRef]
- Hartman, S.E.; Jiang, Z.-P.; Turk, D.; Lampitt, R.S.; Frigstad, H.; Ostle, C.; Schuster, U. Biogeochemical variations at the Porcupine Abyssal Plain Sustained Observatory (PAP-SO) in the northeast Atlantic Ocean, from weekly to inter-annual time scales. Biogeosciences 2015, 12, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Boyer, T.P.; Antonov, J.I.; Baranova, O.K.; Coleman, C.; Garcia, H.E.; Grodsky, A.; Johnson, D.R.; Locarnini, R.A.; Mishonov, A.V.; O’Brien, T.D.; et al. World Ocean Database 2013; NOAA Atlas NESDIS 72; Levitus, S., Mishonov, A., Eds.; National Oceanographic Data Center: Silver Spring, MD, USA, 2013; p. 209. [Google Scholar]
- Poulton, A.J.; Charalampopoulou, A.; Young, J.R.; Tarran, G.A.; Lucas, M.I.; Quartly, G.D. Coccolithophore dynamics in non-bloom conditions during late summer in the central Iceland Basin (July–August 2007). Limnol. Oceanogr. 2010, 55, 1601–1613. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.H.; Harbour, D.S.; Harris, R.P.; Burkill, P.H.; Edwards, E.S. Seasonal succession in the pelagic ecosystem of the North Atlantic and the utilization of nitrogen. J. Plankton Res. 1993, 15, 875–891. [Google Scholar] [CrossRef]
- Continuous Plankton Recorder Survey Team. Continuous Plankton Records: Plankton atlas of the North Atlantic Ocean. Mar. Ecol. Prog. Ser. Suppl. 2004, 11–75. [Google Scholar]
- Fehling, J.; Davidson, K.; Bolch, C.; Tett, P. Seasonality of Pseudo-nitzschia spp. (Bacillariophyceae) in western Scottish waters. Mar. Ecol. Prog. Ser. 2006, 323, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, A.; Harrison, P.J. Coupled changes in the cell morphology and the elemental (C, N, and Si) composition of the pennate diatom Pseudo-nitzschia due to iron deficiency. Limnol. Oceanogr. 2007, 52, 2270–2284. [Google Scholar] [CrossRef]
- Leblanc, K.B.; Queguiner, F.; Diaz, V.; Cornet, M.; Michel-Rodriguez, X.; Durrieu de Madron, C.; Bowler, S.; Malviya, M.; Thyssen, G.; Gregori, M.; et al. Nanoplanktonic diatoms are globally overlooked but play a role in spring blooms and carbon export. Nat. Commun. 2018, 9, 953. [Google Scholar] [CrossRef] [PubMed]
- Savidge, G.; Turner, D.R.; Burkill, P.H.; Watson, A.J.; Angel, M.V.; Pingree, R.D.; Leach, H.; Richards, K.J. The BOFS 1990 spring bloom experiment: Temporal evolution and spatial variability of the hydrogrpahic field. Prog. Oceanogr. 1992, 29, 235–281. [Google Scholar] [CrossRef]
- Savidge, G.; Boyd, P.; Pomroy, A.; Harbour, D.; Joint, I. Phytoplankton production and biomass estimates in the northeast Atlantic Ocean, May–June 1990. Deep Sea Res. I 1995, 42, 599–617. [Google Scholar] [CrossRef]
- Nienow, J.A.; Snyder, R.A.; Jeffrey, W.H.; Wise, S. Fine structure and ecology of Nanoneis longta in the northeastern Gulf of Mexico with a revised definition of the species. Diatom Res. 2016, 32, 43–58. [Google Scholar] [CrossRef]
- Villar, E.; Ferrant, G.K.; Follows, M.; Garczarek, L.; Speich, S.; Audic, A.; Bittner, L.; Blanke, B.; Brum, J.R.; Brunet, C.; et al. Environmental characteristics of Agulhas rings affect interocean plankton transport. Science 2015, 348, 1261447. Available online: http://www.igs.cnrs-mrs.fr/Tara_Agulhas/#FigW9 (accessed on 20 May 2021). [CrossRef] [Green Version]
- Norris, R.E. A new planktonic diatom, Nanoneis hasleae gen. et sp. nov. Nor. J. Bot. 1973, 20, 321–325. [Google Scholar]
- Lampitt, R.S.; Salter, I.; de Cuevas, B.A.; Hartman, S.; Larkin, K.E.; Pebody, C.A. Long-term variability of downward particle flux in the deep northeast Atlantic: Causes and trends. Deep Sea Res. Part II 2010, 57, 1346–1361. [Google Scholar] [CrossRef]
- Horton, T.; Thurston, M.H.; Vlierboom, R.; Gutteridge, Z.; Pebodu, C.A.; Gates, A.R.; Bett, B.J. Are abyssal scavenging amphipod assemblages linked to climate cycles? Prog. Oceanogr. 2020, 184, 102318. [Google Scholar] [CrossRef]
- Alldredge, A. Particle Aggregation Dynamics. In Encyclopedia of Ocean Sciences, 2nd ed.; Academic Press: Cambridge, MA, USA, 2001; pp. 330–337. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
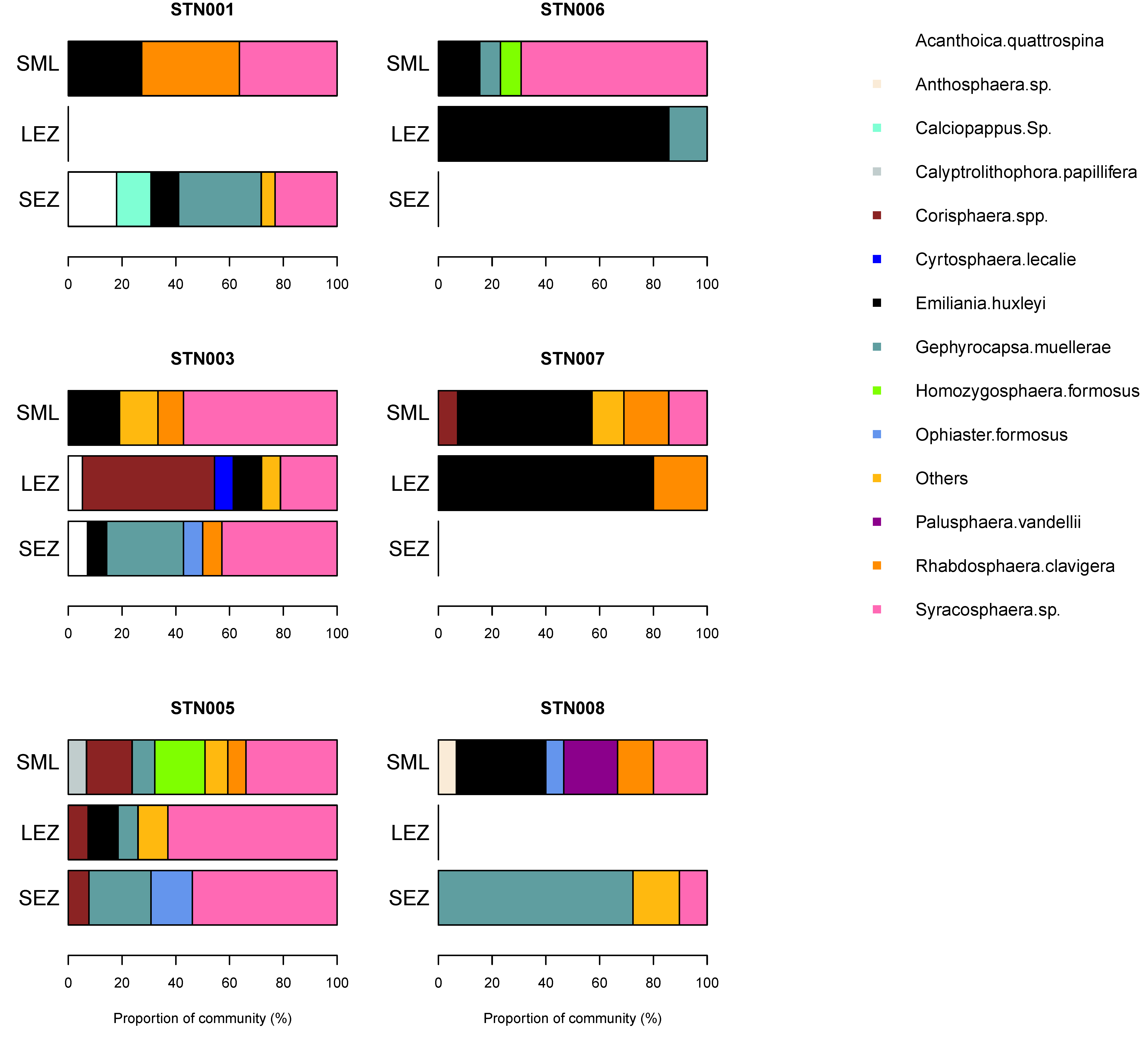{kind=link}
| Species | Coccolith Calcite (pmol) | Coccoliths per Cell | Cellular Calcite (pmol) | Source |
|---|---|---|---|---|
| Emiliania huxleyi | 0.024 | 22 | 0.52 | [45] |
| Syracosphaera spp. | 0.012 | 35 | 0.42 | [45] |
| Gephyrocapsa mullerae | 0.080 | 20 | 1.60 | [46,47] |
| Rhabdosphaera clavigera | 0.67 | 20 | 13.49 | [46,47] |
| Station | Date/Time | Latitude (° N) | Longitude (° W) | Mixed Layer Depth (m) | Temperature (°C) | Salinity (PSS-78) | Total Chl-a (mg/m3) (% <10 μm) | NO3− (μmol L−1) | PO43− (μmol L−1) | Si (μmol L−1) | bSi (μmol L−1) | PIC (μmol L−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 03/09/2013 15:49 | 48.6802 | 16.1907 | 20.9 | 18.96 | 35.59 | 0.58 (88.4) | 0.04 | 0.04 | 0.02 | - | - |
| 2 | 03/09/2013 19:34 | 48.6802 | 16.1907 | 26.8 | 19.35 | 35.71 | 0.41 (91.6) | 0.03 | 0.03 | 0.05 | 0.19 | 0.16 |
| 3 | 04/09/2013 13:01 | 48.6827 | 16.1590 | 24.9 | 19.12 | 35.66 | 0.67 (90.9) | 0.05 | 0.03 | 0.02 | 0.13 | 0.16 |
| 4 | 04/09/2013 21:39 | 48.6685 | 16.2330 | 28.8 | 19.11 | 35.70 | 0.63 (-) | 0.05 | 0.03 | 0.03 | - | - |
| 5 | 06/09/2013 18:03 | 48.6465 | 16.2605 | 24.9 | 18.33 | 35.55 | 0.73 (85.5) | 0.04 | 0.04 | 0.04 | 0.16 | 0.12 |
| 6 | 08/09/2013 18:38 | 48.7102 | 16.2028 | 34.8 | 17.94 | 35.50 | 0.95 (85.5) | 0.09 | 0.05 | 0.07 | 0.23 | 0.15 |
| 7 | 12/09/2013 15:27 | 48.6282 | 16.2760 | 30.8 | 19.01 | 35.71 | 0.52 (94.4) | 0.04 | 0.03 | 0.23 | 0.17 | 0.25 |
| 8 | 13/09/2013 09:29 | 48.6325 | 16.2728 | 34.8 | 19.03 | 35.70 | 0.51 (92.7) | 0.04 | 0.05 | 0.30 | 0.18 | 0.14 |
| Mean ± S.D. | 28.3 ± 5.0 | 18.9 ± 0.5 | 35.64 ± 0.08 | 0.63 ± 0.17 (89.9 ± 3.5) | 0.05 ± 0.02 | 0.04 ± 0.01 | 0.09 ± 0.11 | 0.15 ± 0.07 | 0.14 ± 0.07 |
| Station | Depth of SCM (m) | Light Level % | Total Chl-a (mg m−3) (% <10 μm) | NO2 (μmol L−1) | NO3 (μmol L−1) | PO4 (μmol L−1) | Si (μmol L−1) | bSi (μmol L−1) | PIC (μmol L−1) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 35.0 | 2.9 | 2.85 (91.1) | 0.1 | 0.52 | 0.11 | 0.21 | - | - |
| 2 | 43.7 | 1.2 | 1.30 (93.8) | <LOD | 0.08 | 0.06 | 0.03 | 0.17 | 0.15 |
| 3 | 39.5 | 1.9 | 1.41 (93.9) | 0.1 | 0.46 | 0.11 | 0.06 | 0.17 | 0.15 |
| 4 | 39.3 | 1.9 | 2.01 (-) | 0.1 | 1.00 | 0.15 | 0.09 | - | - |
| 5 | 31.3 | 4.3 | 2.63 (90.4) | <LOD | 0.28 | 0.10 | 0.03 | 0.34 | 0.19 |
| 6 a | 29.9 | 4.9 | 1.43 (85.5) | 0.01 | 0.26 | 0.09 | 0.11 | 0.24 | 0.14 |
| 7 | 41.6 | 1.5 | 0.91 (95.8) | 0.01 | 0.17 | 0.06 | 0.39 | 0.52 | 0.10 |
| 8 | 39.9 | 1.8 | 1.49 (94.1) | 0.24 | 1.31 | 0.20 | 0.96 | 0.49 | 0.20 |
| Mean ± S.D. | 37.5 ± 4.9 | 2.6 ± 1.4 | 1.75 ± 0.68 (92.1 ± 3.4) | 0.09 ± 0.09 | 0.51 ± 0.43 | 0.11 ± 0.05 | 0.23 ± 0.32 | 0.32 ± 0.15 | 0.16 ± 0.04 |
| Station | Depth of PNM (m) | Light Level % | Total Chl-a (mg m−3) (% <10 μm) | NO2 (μmol L−1) | NO3 (μmol L−1) | PO4 (μmol L−1) | Si (μmol L−1) | bSi (μmol L−1) | PIC (μmol L−1) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 47.9 | 0.8 | 1.8 (94.7) | 0.71 | 3.57 | 0.25 | 0.61 | - | - |
| 2 | 60.2 | 0.2 | 1.1 (95.1) | 0.84 | 4.35 | 0.3 | 0.42 | 0.20 | 0.08 |
| 3 | 45.6 | 1.0 | 1.3 (94.4) | 0.31 | 1.2 | 0.19 | 0.09 | 0.22 | 0.15 |
| 4 | 61.1 | 0.2 | 0.3 (-) | 0.61 | 6.28 | 0.4 | 0.52 | - | - |
| 5 | 47.5 | 0.8 | 1.1 (93.6) | 0.8 | 3.8 | 0.31 | 0.26 | 0.22 | 0.15 |
| 6 a | - | - | - | - | - | - | - | - | - |
| 7 | 59.8 | 0.2 | 0.8 (-) | 0.31 | 4.87 | 0.32 | 0.98 | 0.18 | 0.09 |
| 8 | 51.0 | 0.6 | 1.3 (98.0) | 0.82 | 4.42 | 0.36 | 1.27 | 0.42 | 0.13 |
| Mean ± S.D. | 53.3 ± 6.8 | 0.6 ± 0.3 | 1.11 ± 0.44 (95.2 ± 1.7) | 0.63 ± 0.23 | 4.07 ± 1.54 | 0.30 ± 0.07 | 0.59 ± 0.41 | 0.25 ± 0.10 | 0.12 ± 0.03 |
| Station | Integration Depth (m) | Integrated NO3− (mmol m−2) | Integrated PO43− (mmol m−2) | Integrated Si (mmol m−2) | Integrated bSi (mmol Si m−2) | Integrated PIC (mmol C m−2) | Integrated Total Chl-a (mg m2) | Integrated <10 μm Chl-a (mg m2) | % of Total Chl-a in <10 μm Fraction | Integrated >10 μm Chl-a (mg m2) | % of Total Chl-a in >10 μm Fraction |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 50 | 39.87 | 4.71 | 8.87 | - | 65.4 | 59.7 | 91.2 | 5.8 | 8.8 | |
| 2 | 50 | 8.01 | 2.33 | 2.48 | 9.16 | 7.64 | 42.1 | 39.3 | 93.6 | 2.7 | 6.4 |
| 3 | 50 | 13.21 | 3.26 | 2.20 | 7.85 | 8.06 | 44.8 | 41.6 | 92.9 | 3.2 | 7.1 |
| 4 | 50 | 39.32 | 5.42 | 3.59 | - | 58.1 | - | - | - | - | |
| 5 | 50 | 44.66 | 5.56 | 4.37 | 10.40 | 7.09 | 67.4 | 60.2 | 89.4 | 7.1 | 10.6 |
| 6 | 50 | 47.50 | 5.26 | 9.24 | 11.35 | 7.51 | 48.8 | 42.0 | 85.9 | 6.9 | 14.1 |
| 7 | 50 | 14.05 | 2.64 | 16.52 | 17.40 | 10.09 | 38.4 | 35.9 | 93.6 | 2.5 | 6.4 |
| 8 | 50 | 40.94 | 5.96 | 28.61 | 14.24 | 7.98 | 45.0 | 42.4 | 94.2 | 2.6 | 5.8 |
| Mean ± S.D. | 30.95 ± 16.20 | 4.39 ± 1.43 | 9.48 ± 9.09 | 11.73 ± 3.52 | 8.06 ± 1.06 | 51.2 ± 11.0 | 45.9 ± 9.9 | 92 ± 3 | 4.4 ± 2.1 | 8 ± 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Painter, S.C. Minor Contribution by Biomineralizing Phytoplankton to Surface Ocean Biomineral Pools in the Late Stratified Period. Oceans 2021, 2, 489-508. https://doi.org/10.3390/oceans2030028
Painter SC. Minor Contribution by Biomineralizing Phytoplankton to Surface Ocean Biomineral Pools in the Late Stratified Period. Oceans. 2021; 2(3):489-508. https://doi.org/10.3390/oceans2030028
Chicago/Turabian StylePainter, Stuart C. 2021. "Minor Contribution by Biomineralizing Phytoplankton to Surface Ocean Biomineral Pools in the Late Stratified Period" Oceans 2, no. 3: 489-508. https://doi.org/10.3390/oceans2030028
APA StylePainter, S. C. (2021). Minor Contribution by Biomineralizing Phytoplankton to Surface Ocean Biomineral Pools in the Late Stratified Period. Oceans, 2(3), 489-508. https://doi.org/10.3390/oceans2030028