
Shallow-Water Species Diversity of Common Intertidal Zoantharians (Cnidaria: Hexacorallia: Zoantharia) along the Northeastern Coast of Trinidad, Southern Caribbean


Abstract

1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. Morphological Analyses and Specimen Collection
2.3. DNA Extraction, PCR and ITS 2
2.4. Phylogenetic Analysis
3. Results
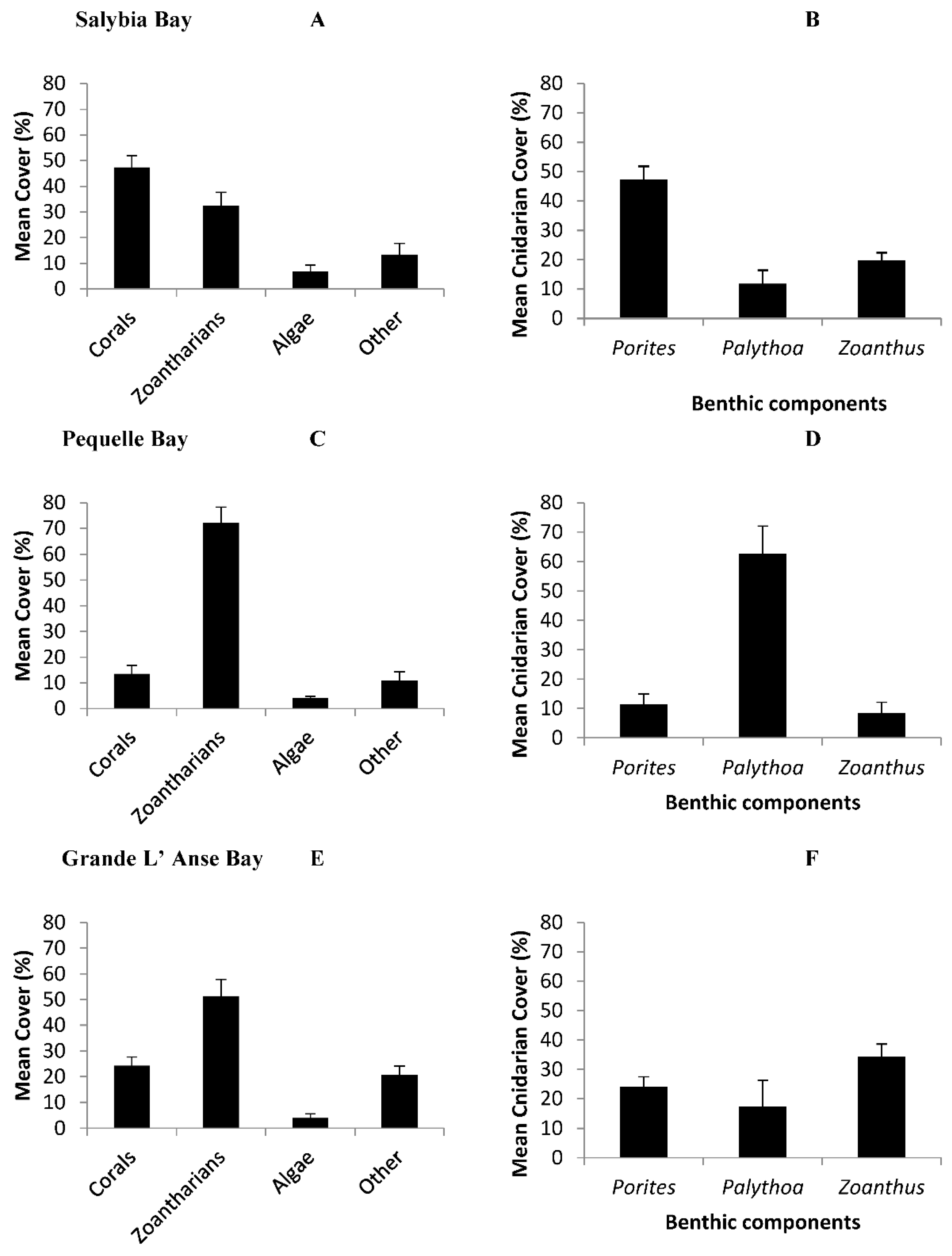
3.1. Zoantharian Distribution
3.2. Specimen Morphological and Molecular Analyses
4. Discussion
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karlson, R.H. Alternate competitive strategies in a periodically disturbed habitat. Bull. Mar. Sci. 1980, 30, 894–900. [Google Scholar]
- Irei, Y.; Nozawa, Y.; Reimer, J.D. Distribution patterns of five zoanthid species in Okinawa Island, Japan. Zool. Stud. 2011, 50, 426–433. [Google Scholar]
- Irei, Y.; Sinniger, F.; Reimer, J.D. Description of two azooxanthellate Palythoa species (Subclass Hexacorallia, Order Zoantharia) from Ryukyu Archipelago, southern Japan. Zookeys 2015, 478, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Reimer, J.D.; Shusuke, O.; Takishita, K.; Tsukahara, J.; Maruyama, T. Molecular evidence suggesting species in the zoanthid genera Palythoa and Protopalythoa (Anthozoa: Hexacorallia) are congeneric. Zool. Sci. 2006, 23, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Sinniger, F.; Montoya-Burgos, J.I.; Chevaldonné, P.; Pawlowski, J. Phylogeny of the order Zoantharia (Anthozoa, Hexacorallia) based on the mitochondrial ribosomal genes. Mar. Biol. 2005, 147, 1121–1128. [Google Scholar] [CrossRef]
- Karlson, R.H. Reproduction patterns on Zoanthus spp. from Discovery Bay, Jamaica. Proceedings 4th International Coral Reef Symposium. Manilla 1981, 2, 699–704. [Google Scholar]
- Sebens, P.S. Intertidal distribution of zoanthids on the Caribbean coast of Panama: Effects of predation and desiccation. Bull. Mar. Sci. 1982, 32, 316–335. [Google Scholar]
- Reimer, J.D.; Ono, S.; Fujiwara, Y.; Takishita, K.; Tsukahara, J. Reconsidering Zoanthus spp. diversity: Molecular evidence of conspecificity within four previously presumed species. Zool. Sci. 2004, 21, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Belford, S.G.; Phillip, D.A.T. Rapid assessment of a coral reef community in a marginal habitat in the southern Caribbean: S simple way to know what’s out there. Asian J. Biol. Sci. 2011, 4, 520–531. [Google Scholar] [CrossRef][Green Version]
- Belford, S.G.; Phillip, D.A.T. Intertidal distribution patterns of zoanthids compared to their scleractinian counterparts in the southern Caribbean. Int. J. Oceanogr. Mar. Ecol. Syst. 2012, 3, 67–75. [Google Scholar] [CrossRef][Green Version]
- Reimer, J.D.; Wee, H.B.; García-Hernández, J.E.; Hoeksema, B.W. Zoantharia (Anthozoa: Hexacorallia) abundance and associations with Porifera and Hydrozoa across a depth gradient on the west coast of Curaçao. Syst. Biodivers. 2018, 16, 820–830. [Google Scholar] [CrossRef]
- Belford, S.G.; Phillip, D.A.T.; Rutherford, M.G.; Schmidt, R.S.; Duncan, E.J. Biodiversity of coral reef communities in marginal environments along the north-eastern coast of Trinidad, southern Caribbean. Prog. Aqu. Farm. Mar. Biol. 2019, 2, 180017. [Google Scholar]
- Goreau, T.F. The ecology of Jamaican coral reefs I. Species composition and zonation. Ecology 1959, 40, 67–90. [Google Scholar] [CrossRef]
- Burnett, W.J.; Benzie, J.A.H.; Beardmore, J.A.; Ryland, J.S. Zoanthids (Anthozoa, Hexacorallia) from the Great Barrier Reef and Torres Straights, Australia: Systematics, evolution and a key to species. Coral Reefs 1997, 16, 55–68. [Google Scholar] [CrossRef]
- Ryland, J.S.; Lancaster, J.E. Revision of methods for separating species of Protopalythoa (Hexacorallia: Zoanthidea) in the tropical West Pacific. Invert. Syst. 2003, 17, 407–428. [Google Scholar] [CrossRef]
- López, C.; Reimer, J.D.; Brito, A.; Simón, D.; Clemente, S.; Hernández, M. Diversity of zoantharian species and their symbionts from Macaronesian and Cape Verde ecoregions demonstrates their widespread distribution in the Atlantic Ocean. Coral Reefs 2019, 38, 269–283. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.S. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbiont. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Burnett, W.J. Longitudinal variation in algal symbionts (zooxanthellae) from the Indian Ocean zoanthid Palythoa caesia. Mar. Ecol. Prog. Ser. 2002, 234, 105–109. [Google Scholar] [CrossRef]
- Reimer, J.D.; Shusuke, O.; Yasuo, F.; Junzo, T. Seasonal changes in morphological condition of symbiotic dinoflagellates (Symbiodinium spp.) in Zoanthus sansibaricus (Anthozoa: Hexacorallia) in Southern Japan. South Pac. Stud. 2007, 27, 2. [Google Scholar]
- Reimer, J.D.; Foord, C.; Irei, Y. Species diversity of shallow water zoanthids (Cnidaria: Anthozoa: Hexacorallia) in Florida. Hindawi Publ. Corp. J. Mar. Biol. 2012, 2012, 856079. [Google Scholar] [CrossRef]
- Pax, F. Studien an westindischen Actinien. In Ergebnisse einer Zoologischen nach Westindien von Prof. W. Kukenthal und Dr. R. Hartmeyer im Jahre, 1907; Spengel, J.W., Ed.; Zoologische Jahrbucher Supplement: Ann Arbor, MI, USA, 1910; Volume 11, pp. 157–330. [Google Scholar]
- Sinniger, F.; Reimer, J.; Pawlowski, J. The Parazoanthidae (Hexacorallia: Zoantharia) DNA taxonomy: Description of two new genera. Mar. Biodivers. 2010, 40, 57–70. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Trench, R. Biogeography of two species of Symbiodinium (Freudenthal) inhabiting the intertidal sea anemone Anthopleura elegantissima (Brandt). Biol. Bull. 2000, 199, 126–134. [Google Scholar] [CrossRef] [PubMed]
- LaJeunesse, T.C. Diversity and community structure of symbiotic dinoflagellates from Caribbean coral reefs. Mar. Biol. 2002, 141, 387–400. [Google Scholar]
- LaJeunesse, T.C.; Thornhill, D.J. Improved resolution of reef-coral endosymbiotic dinoflagellate (Symbiodinium) species diversity, ecology, and evolution through psbA non-coding region genotyping. PLoS ONE 2011, 6, e29013. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Strecher, G.; Li, M.; Knyaz, C.; Tamura, K. Molecular evolutionary genetic analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, E.F.; Soares, M.D.; Bezerra, L.E.; Matthews-Cascon, H. Distribution patterns of zoanthids (Cnidaria: Zoantharia) on a tropical reef. Mar. Biol. Res. 2015, 11, 584–592. [Google Scholar] [CrossRef]
- Belford, S.G. Spatial abundance and colour morphotype densities of the rock boring sea urchin (Echinometra lucunter) at two different habitats. Thalassas 2020, 36, 157–164. [Google Scholar] [CrossRef]
- López, C.; Freitas, R.; Magileviciute, E.; Ratão, S.S.; Brehmer, P.; Reimer, J.D. Report of a Zoanthus zone from the Cabo Verde islands (Central eastern Atlantic). Thalassas 2018, 34, 409–413. [Google Scholar] [CrossRef]
- Ong, C.W.; Reimer, J.D.; Todd, P.A. Morphological plastic responses to shading in the zoanthids Zoanthus sansibaricus and Palythoa tuberculosa. Mar. Biol. 2013, 160, 1053–1064. [Google Scholar] [CrossRef]
- Sinniger, F.; Reimer, J.D.; Pawlowski, J. Potential of DNA sequences to identify zoanthids (Cnidaria: Zoantharia). Zool. Sci. 2008, 25, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- LaJeunesse, T.C.; Loh, W.; Van Woesik, R.; Hoegh-Guldberg, O.; Schmidt, G.; Fitt, W. Low symbiont diversity in southern Great Barrier Reef corals, relative to those of the Caribbean. Am. Soc. Limnol. Oceanogr. 2003, 48, 2046–2054. [Google Scholar] [CrossRef]
- Kumara, S.; Zacharia, P.U.; Sreenath, K.R.; Kripa, V.; George, G. GIS based mapping of zoanthids along Saurashtra coast, Gujarat, India. J. Mar. Biol. Assoc. India 2017, 59, 19–25. [Google Scholar]
- Hughes, T.P.; Baird, A.H.; Bellwood, D.R.; Card, M.; Connolly, S.R.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.B.C.; Kleypas, J.; et al. Climate change human impacts and the resilience of coral reefs. Science 2003, 301, 929–933. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample # | Collection Site α | Disk Color | Tentacle Color | Tentacle Count (3 Polyps Per Colony) | Disk Diameter (mm) | COI I.D. (GenBank Accession #) |
|---|---|---|---|---|---|---|
| P1-br-GA | GA | Brown | Brown | 26 | 11.0 | P. caribaeorum (MZ150796) |
| P2-br-GA | GA | Brown | Brown | 36 | 10.8 | P. caribaeorum (MZ150797) |
| P3-br-SB | SB | Brown | Brown | 37 | 10.5 | P. caribaeorum (MZ150798) |
| P4-gr-GA | GA | Green | Brown | 46 | 13.8 | P. grandiflora (MZ150799) |
| P5-gr-GA | GA | Green | Brown | 45 | 12.6 | P. grandiflora (MZ150800) |
| P6-br-GA | GA | Brown | Brown | 33 | 10.7 | P. caribaeorum (MZ150801) |
| P7-br-WSB | WSB | Brown | Brown | 26 | 11.0 | P. caribaeorum (MZ147090) |
| P8-br-WSB | WSB | Brown | Brown | 26 | 11.0 | P. caribaeorum (MZ147091) |
| Z1-gr-StB | StB | Green/Blue | Green | 41 | 10.2 | Z. sociatus (MZ147096) |
| Z2-gr-StB | StB | Green/Blue | Green | 42 | 10.3 | Z. sociatus (MZ147097) |
| Z3-gr-WSB | WSB | Green/Blue | Green | 42 | 10.3 | Z. sociatus (MZ150806) |
| Z4-gr-WSB | WSB | Green/Blue | Green | 42 | 10.3 | Z. sociatus (MZ150802) |
| Z5-gr-SB | SB | Green | Green | 42 | 7.5 | Z. pulchellus (MZ150803) |
| Z6-gr-SB | SB | Green | Green | 42 | 6.9 | Z. pulchellus (MZ156026) |
| Z7-br-gr-SB | SB | Green/Blue | Green | 42 | 7.6 | Z. sociatus (MZ150807) |
| Z8-org-SB | SB | Green | Green | 46 | 10.2 | Z. pulchellus (MZ150805) |
| Z9-org-GA | GA | Orange | Brown | 46 | 10.3 | Z. pulchellus (MZ150804) |
| Z10-gr-TLH | TLH | Green | Green | 42 | 10.3 | Z. pulchellus (MZ147092) |
| Z13-grey-GA | SB | Grey | Green | 44 | 5.2 | Z. pulchellus (MZ147093) |
| Z14-org-GA | SB | Orange | Brown | 45 | 10.2 | Z. pulchellus (MZ147094) |
| Z15-blu-TLH | TLH | Green/Blue | Green | 42 | 10.3 | Z. sociatus (MZ147095) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belford, S. Shallow-Water Species Diversity of Common Intertidal Zoantharians (Cnidaria: Hexacorallia: Zoantharia) along the Northeastern Coast of Trinidad, Southern Caribbean. Oceans 2021, 2, 477-488. https://doi.org/10.3390/oceans2030027
Belford S. Shallow-Water Species Diversity of Common Intertidal Zoantharians (Cnidaria: Hexacorallia: Zoantharia) along the Northeastern Coast of Trinidad, Southern Caribbean. Oceans. 2021; 2(3):477-488. https://doi.org/10.3390/oceans2030027
Chicago/Turabian StyleBelford, Stanton. 2021. "Shallow-Water Species Diversity of Common Intertidal Zoantharians (Cnidaria: Hexacorallia: Zoantharia) along the Northeastern Coast of Trinidad, Southern Caribbean" Oceans 2, no. 3: 477-488. https://doi.org/10.3390/oceans2030027
APA StyleBelford, S. (2021). Shallow-Water Species Diversity of Common Intertidal Zoantharians (Cnidaria: Hexacorallia: Zoantharia) along the Northeastern Coast of Trinidad, Southern Caribbean. Oceans, 2(3), 477-488. https://doi.org/10.3390/oceans2030027