1. Introduction
New Zealand farmers traditionally milk their cows twice a day (TAD). However, since the late 1990s, the interest in once-a-day (OAD) milking has grown [
1] due to several benefits: reduced labour, managemen, and feed cost, improved labour efficiency, cow health and reproductive performance, and the creation of alternative employment options for farmers [
2,
3,
4]. However, OAD milking reduces milk production, which leads to economic uncertainty [
3]. Once-a-day milking also modifies the milk composition, with higher concentrations of fat and protein, and lower concentrations of lactose compared to TAD milking [
5,
6].
Milk coagulation properties (MCPs) are measures of milk processability for cheese making. Typical MCPs are rennet coagulation time (RCT, min), curd firming time or time taken to form 20 mm depth curd (K
20, min), and curd firmness 30 min after rennet addition (A
30, mm) using a Formagraph. The MCP methodology has been reviewed by Troch et al. [
7]. In cheese production, milk that aggregates quickly (low RCT) and forms a firm curd soon after the addition of the clotting enzyme (low K
20 and high A
30 values) is desirable.
Milk composition affects the yield and quality of dairy products [
8]. Milk coagulation properties are crucial factors and basic requirements for cheese making [
9]. Fat and protein are the most important milk solids that affect MCPs. Milk protein contributes to the functional properties of coagulation such as gel formation, curd formation, ion binding, and emulsifying ability [
8]. Casein is the most important protein component affecting the coagulation properties of milk. Generally, milk with a higher ratio of casein to whey protein is highly suitable for cheese-making [
10]. A higher concentration of calcium ions in milk also leads to good quality cheese, aggregating para-casein micelles by reducing the surface charges [
10,
11].
Selecting breeds that produce milk more suitable for manufacturing dairy products is one way to manipulate milk processing properties [
12]. Milk from Jersey cows has higher concentrations of fat and protein in both OAD and TAD milking frequencies [
12,
13]. There are studies in New Zealand reporting the effect of breed on milk composition and MCPs in cows milked with TAD [
12,
14] and international studies reporting the effect of gross composition on the coagulation characteristic of OAD milk [
15,
16,
17]. However, no studies have been conducted to investigate the effect of breed on the composition and coagulation properties of milk from cows milked OAD and TAD with different milking times. Studies have also reported differences in the composition of morning and afternoon milking [
18,
19], which could also affect the MCPs. The objective of this study was to investigate the effect of breed on the composition and coagulation properties of milk from cows milked OAD and TAD with different milking times.
2. Materials and Methods
2.1. Animal Management and Feeding
This study was conducted at the No. 1 Dairy farm and No. 4 Dairy farm of Massey University, Palmerston North during the 2020–2021 production season. The No. 1 Dairy farm is managed as a low input system with cows milked OAD, and the No. 4 Dairy farm is managed as a high input system with cows milked TAD. The details of farm and milking frequencies have previously been given in detail by Sanjayaranj et al. [
20]. The feed composition and chemical composition of the feed given in the production season are shown in
Table 1.
2.2. Milk Sampling
Milk samples were collected from 30 cows (11 Holstein-Friesian (F), 8 Holstein-Friesian × Jersey cross (F × J), and 11 Jersey (J)) from an OAD milking herd and 30 cows (16 F, 10 F × J, and 4 J) from a TAD milking herd during late lactation (229 ± 13 days in milk) in the 2020–2021 production season. The sampled cows from each No.1 Dairy farm and No.4 Dairy farm contained 15 first parity and 15 second parity cows and had no clinical mastitis or metabolic diseases. Morning milk samples were collected from the OAD herd, and morning and evening milk samples were collected from the TAD herd separately. Composite milk samples were collected using a Waikato milk flow meter. All samples were refrigerated at 0–4 °C without adding any preservatives and analysed within 2 days of sampling.
2.3. Analysis of MCPs
The MCPs were measured using a Formagraph [
7,
21] (Foss Electric, DK-3400, Hillerød, Denmark) of the model type 20: 1 recorder + 1 service module with the capacity of 20 samples/hour. Each sample of 10 mL milk was heated to 35°C and mixed with 0.2 mL rennet solution. Rennet powder (1100 IMCU/g with 96% chymosin and 4% pepsin; prepared according to IDF standard 157:2007 by Renco, New Zealand) was freshly diluted with Milli-Q water to obtain a solution of 0.24% (wt/vol), with a final value of 0.0528 IMCU/mL before addition. Rennet coagulation time (RCT, min), curd firming time or time taken to form 20 mm depth curd (K
20, min), and curd firmness 30 min after rennet addition (A
30, mm) were measured for two replicates for each sample for 30 min. The Formagraph was used in this study due to the large number of samples, as it allows a minimum of 10 milk samples to coagulate in a maximum of 30 min.
The following model was developed to predict the average curd firmness over time for the different milking groups to allow the shape of the curves to be described. This model is a 2nd order plus dead time (SOPDT) model [
22] to get a better fit early in the response to improve the predictive capability of the model.
where
CFt is the curd firmness at time
t (mm);
CFp is the asymptotical potential value of curd firmness at an infinite time (mm);
τ1 and
τ2 are the time constants;
RCTeq is the rennet coagulation time equivalent to the
RCT in traditional Formagraph. This model was used to illustrate the different shapes of the Formagraph plots over time for the different milking groups.
2.4. Determination of Milk Composition
Concentrations of fat, protein, lactose, and total solids of all the samples were measured using a Milkoscan FT1 instrument (Foss Analytical, Hillerød, Denmark).
2.5. Statistical Analysis
Statistical analysis was performed using SAS version 9.4 (SAS Institute Inc., Cary, NC, USA). Least-squares means, and standard errors of the variables were obtained using the following linear model using the GLM procedure.
Yijkm is the observation for composition and coagulation traits for milking frequency-milking time i, breed group j, lactation number k, and cow m.
μ is the population mean.
MSi is the fixed effect of milking frequency-milking time i (i = OAD, TAD-morning, and TAD-afternoon).
Bj(MSi) is the fixed effect of breed j nested in milking frequency-milking time i (j = F, F × J, and J).
Lk is the fixed effect of lactation number (k = 1st and 2nd lactation).
β1 is the regression coefficient associated with the linear effect of deviation from the median calving date of cow m.
eijkm is the residual random error assumed with mean zero and variance .
A limitation of this study was the confounding effect of feed and MF. The feed provided by each farm was not identical during the experimental period. The cows milked TAD were fed slightly higher supplements than cows milked OAD. However, the genetic merit of the cows in the two farms was similar because there were a few sires that had progeny in both farms.
Partial correlations between the dependent variables in each milking frequency-milking time were performed by MANOVA using the GLM procedure of SAS adjusting for the fixed effects of breed and lactation number.
Above GLM model was used with the direct measurements of MCPs from Formagraph such as RCT, K20, and A30.
3. Results
Descriptive statistics of daily milk yield, milk composition, and MCPs of cows sampled in the study are presented in
Table 2.
The minimum value for A
30 was 0.01 mm. A very low value for A
30 was observed for 14% of F cows and 7% of F × J cows. F-values for factors affecting milk yield, milk composition traits, and MCPs are presented in
Table 3. The milking frequency-milking time explained the largest proportion of variation in most traits, except for lactose percentage, K
20, and A
30; for these traits milking frequency-milking time was non-significant. Breed nested in milking frequency-milking time explained the significant variation in all traits except for lactose percentage. For lactose percentage and A
30, the lactation number explained the majority of variations.
Records with poorly correlated samples were included when calculating the least-squares means to avoid bias in the results. The least-squares means and standard errors for the milk composition and coagulation properties for F, F × J, and J cows milked OAD or TAD in the morning and afternoon are shown in
Table 4. Milk from F × J cows milked TAD morning had significantly (
p < 0.05) higher fat concentrations compared to F cows. Jersey cows milked TAD in the afternoon produced milk with a higher concentration of fat compared to F cows. Similarly, Jersey cows milked OAD and TAD in the morning produced milk with a significantly higher concentration of protein compared to F cows. Milk from J cows coagulated significantly faster (lower RCT and K
20 means) and produced a significantly firmer curd (higher A
30 mean) compared to F cows in each milking frequency-milking time. The K
20 was significantly lower in J cows milked TAD in the morning and afternoon compared to F cows. Friesian × Jersey cows had intermediate values for most MCPs in each milking frequency-milking time, but most differences were not significant.
In cows milked OAD, all three coagulation properties showed no significant correlation with fat concentration (
Table 5). However, the correlation between K
20 and protein percentage was negative (
p < 0.05). The correlation between A
30 and lactose percentage was positive (
p < 0.05). The results also show a strong correlation coefficient among MCPs, with RCT being positively and significantly correlated with K
20 and negatively and significantly correlated with A
30.
In cows milked TAD in the morning, RCT was negatively (
p < 0.05) correlated with fat concentration (
Table 6). The time taken to form a 20 mm curd width after rennet addition (K
20) was significantly and negatively correlated with the concentrations of fat and protein, whereas A
30 was significantly and positively correlated with fat and protein concentrations. In cows milked TAD in the morning and afternoon, the correlation between RCT and K
20 was positive (
p < 0.05) and the correlation between RCT and A
30 was strongly negative (
p < 0.05).
The partial correlation coefficients between coagulation properties and milk components of cows milked in the study are presented in
Table 7. Curd firming time (K
20) was significantly and negatively correlated with the concentrations of fat and protein whereas A
30 was significantly and positively correlated with fat and protein concentrations.
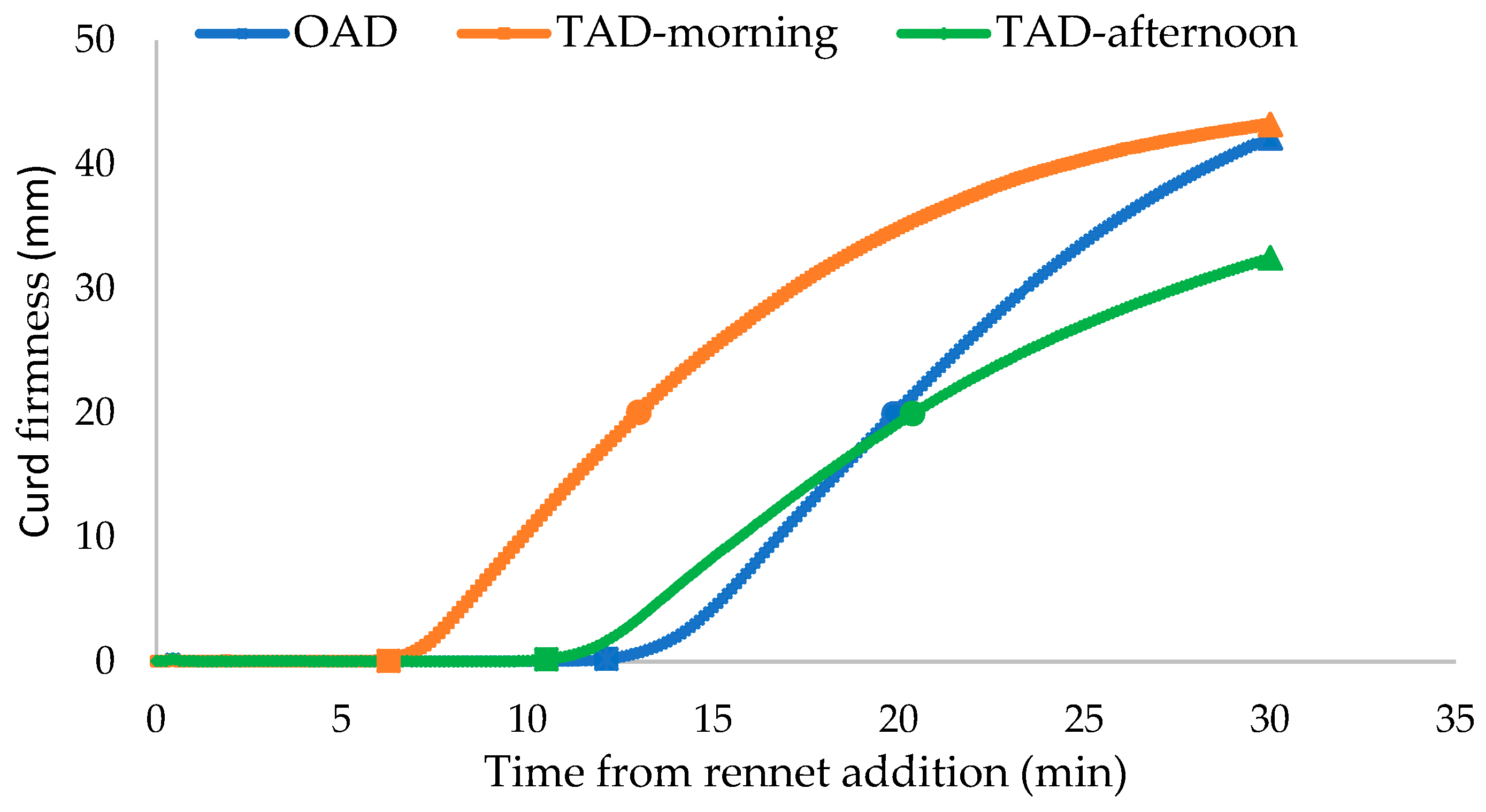
Figure 1 shows the differences between the curd firmness over time from the rennet addition of milk from cows milked OAD and cows milked TAD in the morning and afternoon. The curves were obtained by averaging the curd firmness as measured by the Formagraph. Twice-a-day morning milk coagulated faster than TAD afternoon milk and OAD milk.
Figure 1.
The pattern of curd firming after rennet addition of milk from cows milked once a day (OAD) and twice a day (TAD) (morning and afternoon) in the late lactation in the 2020–2021 production season.
Figure 1.
The pattern of curd firming after rennet addition of milk from cows milked once a day (OAD) and twice a day (TAD) (morning and afternoon) in the late lactation in the 2020–2021 production season.
However, as per the model developed, the asymptotical potential value of curd firmness at an infinite time was higher for OAD milk (53.05 mm) compared to TAD-morning milk (47.69 mm). The asymptotical potential value of curd firmness for TAD-afternoon milk was 43.50 mm.
4. Discussion
The present study evaluated the effect of breed (F, F × J, and J cows) and milking frequency-milking time (OAD and TAD in the morning and afternoon in late lactation) on milk composition and coagulation parameters. It has been previously identified that breed significantly affects MCPs [
23,
24]. However, little information is available regarding the effects of the breed with the interaction of milking frequency on MCPs.
Jersey cows were superior in the concentrations of fat and protein compared to F cows in each milk milking frequency-milking time. These results are in agreement with Auldist et al. [
12] and Lopez-Villalobos et al. [
25] in cows milked TAD and Tong et al. [
3] and Clark et al. [
6] in cows milked OAD. Higher concentrations of fat and protein could be a reason for faster K
20 and greater curd firmness (A
30) in J cows compared to F cows within the milking frequency-milking time. Similar results were reported by Auldist et al. [
12], Poulson et al. [
26] and Stocco et al. [
27]. Jersey cows were superior for MCPs compared to F cows in each milking frequency-milking time, with F × J cows being intermediate in most cases. A higher concentration of fat in F × J cows milked OAD would be due to the heterosis effect in which the progeny performs better than the average performance of the parents.
The fat concentration of milk is mainly associated with curd firmness as in the present study. Huppertz and Kelly [
28] reported that milk flocculation occurs due to the collision and aggregation of fat globules and Stocco et al. [
29] reported that this could affect the RCT. The role of fat in milk is considered as an inert filler between the gel network formed by the casein micelles and contributes to the curd structure [
29,
30,
31]. More accurately, the ratio of casein: fat is more important for the coagulum strength than the concentration of fat on its own [
10]. A higher concentration of protein means a higher concentration of casein as 80% of the protein in milk is casein. Milk with a higher concentration of casein leads to the formation of curd with higher firmness [
14,
32]. Of the different casein fractions, κ-casein is the fraction that initiates the process of cheese-making by interacting with chymosin. When the concentration of κ-casein is higher, the κ-casein hydrolysed by the rennet enzyme would be higher, which leads to the aggregation of more hydrolysed casein and results in a firmer curd. The results of this study suggest that the differences in the MCPs of breeds could be mainly due to the variation in the concentration of fat and protein between the breeds. Auldist et al. [
12] reported that higher curd firmness in milk from J cows was obvious compared to F cows, in milk from individual cows, in bulk milk, and in milk standardised for protein: fat. However, this difference disappeared when bulk milk was standardised for constant solid concentration. Similarly, Stocco et al. [
27] reported, that the differences between breeds for MCPs were mainly due to the variation in the genetic factors controlling the production of milk fat, protein, and lactose. This study suggests the same.
In this study correlations between milk composition and MCPs and correlations among MCPs were obtained. When considering MCPs, strong and negative correlations between RCT and A
30 were reported, which was in agreement with Okigbo et al. [
33]. Lower RCT means the presence of a higher concentration of protein, especially casein, to form the three-dimensional network and the gel faster which leads to firmer curd. Rennet coagulation time also positively correlated with K
20, meaning when RCT is lower the formation of a three-dimensional network and gel is faster, therefore the time taken to form 20 mm width curd (A
20) will be lower. When the concentrations of fat and protein were higher, K
20 was faster due to the availability of more hydrolysed casein aggregation and trapped fat inside the gel network. However, all these processes depend on the substrate-to-enzyme ratio.
The correlation coefficients between MCPs and the concentrations of fat and protein were weak to moderate in each milking frequency-milking time. RCT was not related to concentrations of fat or protein except in cows milked TAD in the morning, with higher fat associated with lower RCT. It seems the relationship between K20, and milk composition depends on the milking frequency- milking time because K20 was correlated with the concentration of protein in OAD milk, correlated with fat in TAD afternoon milk, and correlated with both the concentrations of fat and protein in TAD morning milk and in all the milk samples regardless of milking frequency and milking time. Similarly, the higher the concentration of either fat or protein or both, the greater the A30, except for cows milked OAD. This suggests the composition has little effect on MCPs, but they were influenced by milking frequency-milking time.
Similar results were reported by Auldist et al. [
12] in cows milked TAD, in which RCT was not correlated with any of the milk components, and the correlation coefficients for K
20 and A
30 were weak. Stocco et al. [
29] reported that an increase in the fat concentration in goat milk shortened the RCT and curd firming time and affected the A
45 and A
60 in a cubic trend. They also reported that higher protein concentration increased the RCT and that a higher concentration of casein raised the curd firmness. However, the authors noted that the correlations explained only a small proportion of the variation in the MCPs. De Marchi et al. [
23] reported that the concentrations of fat and protein did not control significant variation in MCPs as the MCPs were better in the Rendena breed than Brown Swiss, although the Brown Swiss breed has better milk quality traits, particularly higher fat and protein concentrations. These results suggest that the composition of milk is not a useful indicator of MCPs of milk from cows milked OAD and TAD in the morning and afternoon. This is also supported by the weak to moderate correlation coefficients obtained in this study. It seems the correlation coefficients depended on milking frequency-milking time as the values of the correlation coefficients between milk composition and MCPs were not constant across milking frequency-milking time combinations. Keffored et al. [
34] and Auldist et al. [
12] reported that MCPs are controlled by a more complex combination of parameters than the composition, breed, and protein polymorphism.
One of the important factors affecting MCPs is the genetic variant of κ-casein which can be A or B. The B variant is associated with a higher concentration of protein, and a higher proportion of κ-casein in the total casein and leads to improved MCPs [
35,
36,
37]. In New Zealand and across the world, the A variant of κ-casein is predominant in F cows and the B variant is predominant in J cows [
38]. The superiority of J cows for the MCPs could be due to the presence of the B variant for κ-casein as the B allele has greater MCPs [
10]. Gai et al. [
39] reported that the C variant of α-casein, B and A1 variants of β-casein, and B variants of β-lactoglobulin are superior for MCPs. Similarly, Walsh et al. [
40] and Mayer et al. [
41] reported that both the κ-casein BB genotype and β-lactoglobulin BB genotype have greater MCPs. In addition, the calcium concentration of milk is also crucial for better MCPs. Adding Ca
2+ speeds up the RCT by increasing the aggregation of renneted micelles [
42]. Davis et al. [
43] reported that J cow milk contains a higher Ca: protein ratio. However, Auldist et al. [
12] did not observe this. In the present study, the concentrations of each casein fraction and calcium were not studied. Milk coagulation is a process that seems dependent on milk composition, mineral composition of milk, genetic variants, and maybe a combination of all factors.
Based on the Formagraph results, the TAD morning milk coagulated faster and formed firmer curd at around 30 min compared to the other two milk, but the asymptotical potential value of curd firmness was lower compared to OAD milk. Faster rennet coagulation could be explained by the substrate: enzyme ratio. The ratio of κ-casein: rennet would be lower in TAD morning milk and the proportion of cleaved substrate rose faster to form firm curd. On the other hand, OAD milk has a higher concentration of casein [
16,
17] and therefore, a higher κ-casein: rennet ratio, which led to the proportion of cleaved substrate initially rising more slowly (i.e., longer RCT) but the formation of a more extensive network in a given sufficient time and hence a higher asymptotical potential value than the TAD-morning milk. Stocco et al. [
27] reported that in goat milk, where the concentration of rennet was not adjusted to the concentration of protein, the RCT was longer due to the lower rennet-to-casein ratio. The reason for this is the need for more time to form sufficient hydrolysis of κ-casein to induce the aggregation of para-casein micelles [
44]. Milk from the OAD herd had higher fat and protein concentration compared to TAD morning and afternoon milk. As adding a constant amount of rennet gives a higher substrate: enzyme ratio for higher protein milk, some of the observed results may indicate that the MCPs, as measured by the Formagraph, are being affected by the substrate: enzyme ratio. Therefore, further studies should be conducted while keeping the substrate: enzyme ratio constant.
Low values for A
30 could be due to the poor coagulation ability of milk. De Marchi et al. [
23] reported F cows showed the highest percentage of poorly coagulated samples compared to Simmental and Rendena cows in Italy. Tyriseva et al. [
45] found that noncoagulating milk from F cows was genetically related to each other and suggested that these poor MCPs could be linked to genetic factors. Chiofaloa et al. [
46] proposed that the reason for lower MCPs in F cows could be a higher concentration of plasmin content, which causes proteolysis of casein and partially reduce the casein: true protein ratio. The limitation of using the current Formagraph method is that the time of the coagulation process is set to 30 min. This causes failure in measuring the RCT and K
20 values [
27]. The use of traditional Formagraph test time could introduce a bias in comparing milk [
27] from different breeds and different milking frequency-milking times. The Formagraph is a useful tool to measure the MCPs for 30 min, and models were developed to predict the curd firmness beyond 30 min using the information from the Formagraph [
47].
The results of the present study were obtained in the late lactation of the milking season. Lucey [
48] reported that milk from late lactation is less suitable for producing high-quality dairy products. This could be due to higher plasmin and plasmin activity which cause the deterioration of MCPs by the proteolysis of casein in milk [
49]. Lucy [
48] reported that OAD milking during late lactation would produce milk with higher plasminogen and plasmin activity. This suggests that milk from cows milked OAD in late lactation is less suitable for cheese making. However, Bastian et al. [
50] reported that plasmin activity did not influence the clotting time. In contrast, Tyriseva et al. [
45] and Vacca et al. [
51] reported better MCPs in late lactation milk. It is also important to note that the stage of lactation has a strong connection with the quality of diet. Therefore, the quality and processability of late lactation milk are also decided by the diet given to cows. The study could be repeated using cows in early and mid-lactation with controlled feed to find the suitability of milk for processing of the different breeds and milking frequency.



 ,
, 

{kind=link}