The Correlation between Play Behavior, Serum Cortisol and Neutrophil-to-Lymphocyte Ratio in Welfare Assessment of Dairy Calves within the First Month of Life

 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods

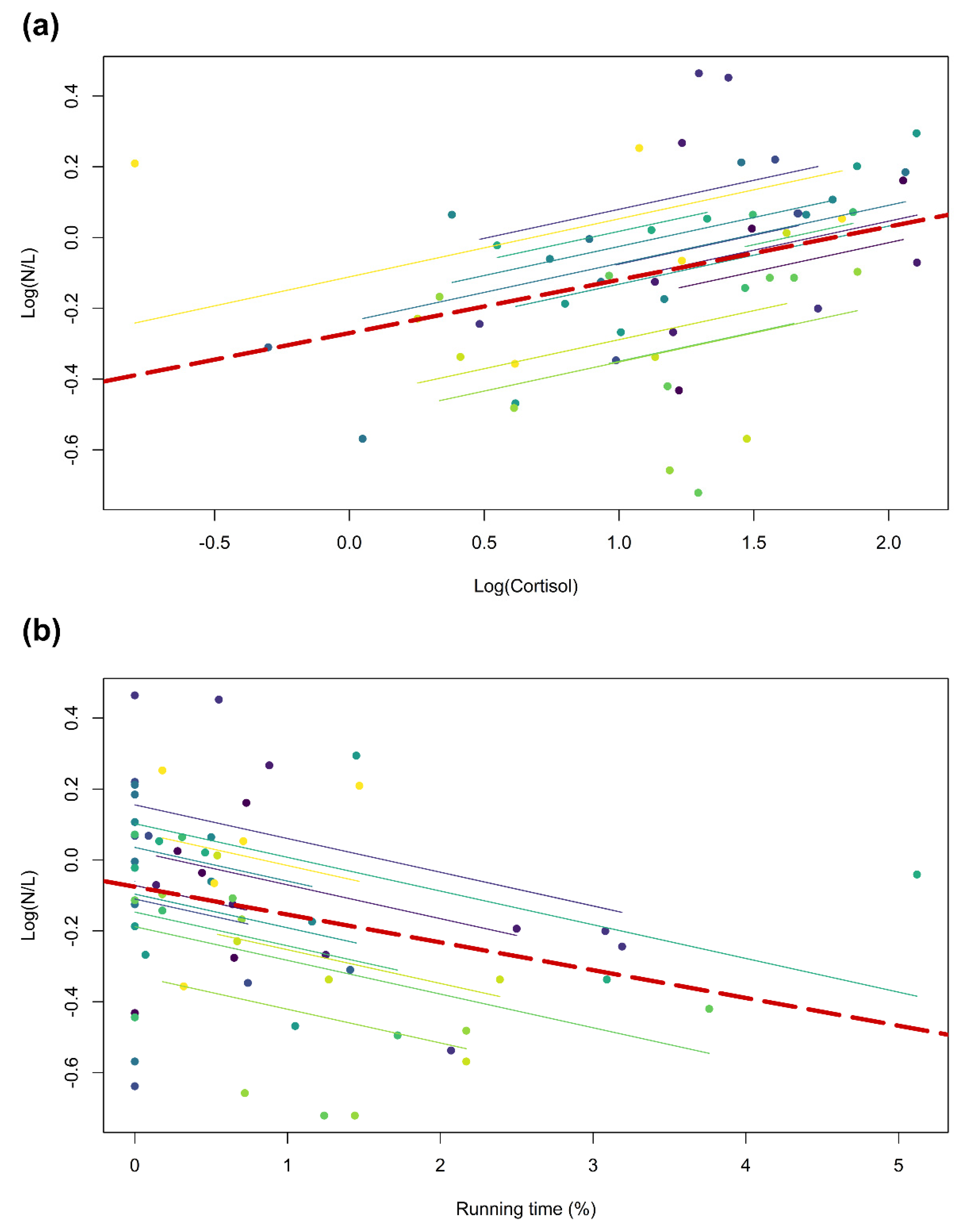{kind=link}
{kind=link}
| Behavior | Definition | Reference |
|---|---|---|
| Running (SE) | Any gait faster than a walk, including trot and gallop. | [32] |
| Leap (PE) | The two forelegs are lifted from the ground and stretched forwards, the forepart of the body is elevated and the calf moves forward. Movement upwards and forwards. During the last phase of the movement, the hind legs may be lifted from the ground. | [22] |
| Jump (PE) | The two forelegs are lifted from the ground, the forepart of the body is elevated. Movement upwards. During the last phase of the movement, the hind legs may be lifted from the ground. | [22] |
| Turn (PE) | The two forelegs are lifted from the ground and stretched forwards, and as the forepart of the body is elevated turned to one side, the calf moves sidewards. Movement upwards and sidewards. During the last phase of the movement, the hind legs may be lifted from the ground. | [22] |
| Buck (PE) | The body ascends from front to back, the top of the head is level with or lower than the shoulders and both hind hooves are lifted off the ground; legs may be kicked outwards. | [32] |
| Kick (PE) | One or both hind legs are lifted off the ground and extended outwards from the body. The calf can be stationary or moving. | [53,54] |
Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cozzi, G.; Gottardo, F.; Brscic, M.; Contiero, B.; Irrgang, N.; Knierim, U.; Pentelescu, O.; Windig, J.J.; Mirabito, L.; Kling Eveillard, F.; et al. Dehorning of cattle in the EU Member States: A quantitative survey of the current practices. Livest. Sci. 2015, 179, 4–11. [Google Scholar] [CrossRef]
- Tremetsberger, L.; Winckler, C. Effectiveness of animal health and welfare planning in dairy herds: A review. Anim. Welf. 2015, 24, 55–67. [Google Scholar] [CrossRef]
- Fraser, D.; Fraser, D. The role of the veterinarian in animal welfare. Animal welfare: Too much or too little? Abstracts of the 21st Symposium of the Nordic Committee for Veterinary Scientific Cooperation (NKVet), Vaerløse, Denmark, 24–25 September 2007. Acta Vet. Scand. 2008, 50. [Google Scholar] [CrossRef]
- Broom, D.M. Animal welfare: Concepts and measurement. J. Anim. Sci. 1991, 69, 4167–4175. [Google Scholar] [CrossRef] [PubMed]
- Broom, D.M. Indicators of poor welfare. Br. Vet. J. 1986, 142, 524–526. [Google Scholar] [CrossRef]
- Duncan, I.J.H. Welfare Is to Do with What Animals Feel. J. Agric. Environ. Ethics 1993, 6, 8–14. [Google Scholar]
- Rollin, B.E. Animal welfare, science, and value. J. Agric. Environ. Ethics 1993, 6, 44–50. [Google Scholar]
- Hemsworth, P.H.; Mellor, D.J.; Cronin, G.M.; Tilbrook, A.J. Scientific assessment of animal welfare. N. Z. Vet. J. 2015, 63, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Bekoff, M. Animal emotions: Exploring passionate natures. Bioscience 2006, 50, 861. [Google Scholar] [CrossRef]
- Gonyou, H.W. Why the study of animal behavior is associated with the animal welfare issue. J. Anim. Sci. 1994, 72, 2171–2177. [Google Scholar] [CrossRef]
- Jensen, M.B.; Kyhn, R. Play behaviour in group-housed dairy calves, the effect of space allowance. Appl. Anim. Behav. Sci. 2000, 67, 35–46. [Google Scholar] [CrossRef]
- Rushen, J.; Wright, R.; Johnsen, J.F.; Mejdell, C.M.; de Passillé, A.M. Reduced locomotor play behaviour of dairy calves following separation from the mother reflects their response to reduced energy intake. Appl. Anim. Behav. Sci. 2016, 177, 6–11. [Google Scholar] [CrossRef]
- Keeling, L.J.; Winckler, C.; Hintze, S.; Forkman, B. Towards a positive welfare protocol for cattle: A critical review of indicators and suggestion of how we might proceed. Front. Anim. Sci. 2021, 2, 1–19. [Google Scholar] [CrossRef]
- Fraser, D.; Duncan, I.J.H. “Pleasures”, “pains” and animal welfare: Toward a natural history of affect. Anim. Welf. 1998, 7, 383–396. [Google Scholar]
- Spinka, M.; Newberry, R.C.; Bekoff, M. Mammalian play: Training for the unexpected. Q. Rev. Biol. 2004, 76, 141–168. [Google Scholar] [CrossRef]
- Held, S.D.E.; Špinka, M. Animal play and animal welfare. Anim. Behav. 2011, 81, 891–899. [Google Scholar] [CrossRef]
- Dawkins, M.S. Battery hens name their price. Anim. Behav. 1983, 31, 1195–1205. [Google Scholar] [CrossRef]
- Lawrence, A. Consumer demand theory and the assessment of animal welfare. Anim. Behav. 1987, 35, 293–295. [Google Scholar] [CrossRef]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef]
- Ahloy-Dallaire, J.; Espinosa, J.; Mason, G. Play and optimal welfare: Does play indicate the presence of positive affective states? Behav. Process. 2018, 156, 3–15. [Google Scholar] [CrossRef]
- Burghardt, G.M. The Genesis of Animal Play: Testing the Limits; MIT Press: Cambridge, MA, USA, 2005; ISBN 0262025434. [Google Scholar]
- Jensen, M.B.; Vestergaard, K.S.; Krohn, C.C. Play behaviour in dairy calves kept in pens: The effect of social contact and space allowance. Appl. Anim. Behav. Sci. 1998, 56, 97–108. [Google Scholar] [CrossRef]
- Valníčková, B.; Stěhulová, I.; Šárová, R.; Špinka, M. The effect of age at separation from the dam and presence of social companions on play behavior and weight gain in dairy calves. J. Dairy Sci. 2015, 98, 5545–5556. [Google Scholar] [CrossRef]
- Hintze, S.; Maulbetsch, F.; Asher, L.; Winckler, C. Doing nothing and what it looks like: Inactivity in fattening cattle. PeerJ 2020, 8, e9395. [Google Scholar] [CrossRef]
- Lidfors, L.M. Behavioural effects of separating the dairy calf immediately or 4 days post-partum. Appl. Anim. Behav. Sci. 1996, 49, 269–283. [Google Scholar] [CrossRef]
- Guardini, G.; Bowen, J.; Mariti, C.; Fatjó, J.; Sighieri, C.; Gazzano, A. Influence of maternal care on behavioural development of domestic dogs (Canis familiaris) living in a home environment. Animals 2017, 7, 93. [Google Scholar] [CrossRef] [PubMed]
- Orihuela, A.; Mota-Rojas, D.; Strappini, A.; Serrapica, F.; Braghieri, A.; Mora-Medina, P.; Napolitano, F. Neurophysiological mechanisms of cow–calf bonding in buffalo and other farm animals. Animals 2021, 11, 1968. [Google Scholar] [CrossRef]
- Orihuela, A.; Galina, C.S. Effects of separation of cows and calves on reproductive performance and animal welfare in tropical beef cattle. Animals 2019, 9, 223. [Google Scholar] [CrossRef] [PubMed]
- Weary, D.M.; Chua, B. Effects of early separation on the dairy cow and calf. Appl. Anim. Behav. Sci. 2000, 69, 177–188. [Google Scholar] [CrossRef]
- Flower, F.C.; Weary, D.M. Effects of early separation on the dairy cow and calf: 2. Separation at 1 day and 2 weeks after birth. Appl. Anim. Behav. Sci. 2001, 70, 275–284. [Google Scholar] [CrossRef]
- Mintline, E.M.; Wood, S.L.; de Passillé, A.M.; Rushen, J.; Tucker, C.B. Assessing calf play behavior in an arena test. Appl. Anim. Behav. Sci. 2012, 141, 101–107. [Google Scholar] [CrossRef]
- Mintline, E.M.; Stewart, M.; Rogers, A.R.; Cox, N.R.; Verkerk, G.A.; Stookey, J.M.; Webster, J.R.; Tucker, C.B. Play behavior as an indicator of animal welfare: Disbudding in dairy calves. Appl. Anim. Behav. Sci. 2013, 144, 22–30. [Google Scholar] [CrossRef]
- Spencer, R.L.; Deak, T. A users guide to HPA axis research. Physiol. Behav. 2017, 178, 43–65. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Kim, E.; Choi, M.H. Technical and clinical aspects of cortisol as a biochemical marker of chronic stress. BMB Rep. 2015, 48, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Gross, W.B.; Siegel, H.S. Evaluation of the Heterophil/Lymphocyte Ratio as a Measure of Stress in Chickens. Avian Dis. 1983, 27, 972. [Google Scholar] [CrossRef]
- Swan, M.P.; Hickman, D.L. Evaluation of the neutrophil-lymphocyte ratio as a measure of distress in rats. Lab Anim. 2014, 43, 276–282. [Google Scholar] [CrossRef]
- Kent, J.E.; Ewbank, R. The effect of road transportation on the blood constituents and behaviour of calves. II. One to three weeks old. Br. Vet. J. 1986, 142, 131–140. [Google Scholar] [CrossRef]
- Weber, P.S.D.; Toelboell, T.; Chang, L.-C.; Tirrell, J.D.; Saama, P.M.; Smith, G.W.; Burton, J.L. Mechanisms of glucocorticoid-induced down-regulation of neutrophil L-selectin in cattle: Evidence for effects at the gene-expression level and primarily on blood neutrophils. J. Leukoc. Biol. 2004, 75, 815–827. [Google Scholar] [CrossRef]
- Lomborg, S.R.; Nielsen, L.R.; Heegaard, P.M.H.; Jacobsen, S. Acute phase proteins in cattle after exposure to complex stress. Vet. Res. Commun. 2008, 32, 575–582. [Google Scholar] [CrossRef]
- Da Costa, M.C.; da Flaiban, K.K.M.C.; Coneglian, M.M.; Dognani, R.; Vettorato, E.D.; Balarin, M.R.S.; Lisbôa, J.A.N. Metabolismo oxidativo dos neutrófilos de bezerros das raças Nelore e Limousin nos primeiros quatro meses de vida. Pesqui. Vet. Bras. 2008, 28, 431–436. [Google Scholar] [CrossRef]
- Benesi, F.J.; Teixeira, C.M.C.; Leal, M.L.R.; Lisboa, J.A.N.; Mirandola, R.M.S.; Shecaira, C.L.; Gomes, V. Leukograms of healthy Holstein calves within the first month of life. Pesqui. Vet. Bras. 2012, 32, 352–356. [Google Scholar] [CrossRef][Green Version]
- Council of European Union. Council Directive 98/58/EC of 20 July 1998 concerning the protection of animals kept for farming purposes. Off. J. Eur. Commun. 1998, 221, 23–27. [Google Scholar]
- Stilwell, G.; Carvalho, R.C.; Carolino, N.; Lima, M.S.; Broom, D.M. Effect of hot-iron disbudding on behaviour and plasma cortisol of calves sedated with xylazine. Res. Vet. Sci. 2010, 88, 188–193. [Google Scholar] [CrossRef]
- Kupczyński, R.; Budny, A.; Śpitalniak, K.; Tracz, E. Dehorning of calves—Methods of pain and stress alleviation—A review. Ann. Anim. Sci. 2014, 14, 231–243. [Google Scholar] [CrossRef]
- Gottardo, F.; Nalon, E.; Contiero, B.; Normando, S.; Dalvit, P.; Cozzi, G. The dehorning of dairy calves: Practices and opinions of 639 farmers. J. Dairy Sci. 2011, 94, 5724–5734. [Google Scholar] [CrossRef]
- Munksgaard, L.; Simonsen, H.B. Behavioral and pituitary adrenal-axis responses of dairy cows to social isolation and deprivation of lying down. J. Anim. Sci. 1996, 74, 769. [Google Scholar] [CrossRef]
- Turini, L.; Conte, G.; Bonelli, F.; Sgorbini, M.; Madrigali, A.; Mele, M. The relationship between colostrum quality, passive transfer of immunity and birth and weaning weight in neonatal calves. Livest. Sci. 2020, 238, 104033. [Google Scholar] [CrossRef]
- Ogi, A.; Mariti, C.; Baragli, P.; Sergi, V.; Gazzano, A. Effects of stroking on salivary oxytocin and cortisol in guide dogs: Preliminary results. Animals 2020, 10, 708. [Google Scholar] [CrossRef] [PubMed]
- Tatara, M.R.; Krupski, W.; Tymczyna, B.; Łuszczewska-sierakowska, I. Biochemical bone metabolism markers and morphometric, densitometric and biomechanical properties of femur and tibia in female and gonadectomised male Polish Landrace pigs. J. Pre-Clin. Clin. Res. 2012, 6, 14–19. [Google Scholar]
- Ekiz, B.; Ergul Ekiz, E.; Kocak, O.; Yalcintan, H.; Yilmaz, A. Effect of pre-slaughter management regarding transportation and time in lairage on certain stress parameters, carcass and meat quality characteristics in Kivircik lambs. Meat Sci. 2012, 90, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Hausman, N.L.; Javed, N.; Bednar, M.K.; Guell, M.; Schaller, E.; Nevill, R.E.; Kahng, S. Interobserver agreement: A preliminary investigation into how much is enough? J. Appl. Behav. Anal. 2021, 9999, 1–12. [Google Scholar] [CrossRef]
- Sutherland, M.A.; Worth, G.M.; Schütz, K.E.; Stewart, M. Rearing substrate and space allowance influences locomotor play behaviour of dairy calves in an arena test. Appl. Anim. Behav. Sci. 2014, 154, 8–14. [Google Scholar] [CrossRef]
- Sutherland, M.A.; Worth, G.M.; Cameron, C.; Ross, C.M.; Rapp, D. Health, physiology, and behavior of dairy calves reared on 4 different substrates. J. Dairy Sci. 2017, 100, 2148–2156. [Google Scholar] [CrossRef] [PubMed]
- Bakdash, J.Z.; Marusich, L.R. Repeated Measures Correlation. Front. Psychol. 2017, 8, 456. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Graf, B.; Senn, M. Behavioural and physiological responses of calves to dehorning by heat cauterization with or without local anaesthesia. Appl. Anim. Behav. Sci. 1999, 62, 153–171. [Google Scholar] [CrossRef]
- Faulkner, P.M.; Weary, D.M. Reducing pain after dehorning in dairy calves. J. Dairy Sci. 2000, 83, 2037–2041. [Google Scholar] [CrossRef]
- Caray, D.; de Boyer des Roches, A.; Frouja, S.; Andanson, S.; Veissier, I. Hot-iron disbudding: Stress responses and behavior of 1- and 4-week-old calves receiving anti-inflammatory analgesia without or with sedation using xylazine. Livest. Sci. 2015, 179, 22–28. [Google Scholar] [CrossRef]
- Stafford, K.J.; Mellor, D.J. Dehorning and disbudding distress and its alleviation in calves. Vet. J. 2005, 169, 337–349. [Google Scholar] [CrossRef]
- Stilwell, G.; de Carvalho, R.C.; Lima, M.S.; Broom, D.M. Effect of caustic paste disbudding, using local anaesthesia with and without analgesia, on behaviour and cortisol of calves. Appl. Anim. Behav. Sci. 2009, 116, 35–44. [Google Scholar] [CrossRef]
- Morisse, J.P.; Cotte, J.P.; Huonnic, D. Effect of dehorning on behaviour and plasma cortisol responses in young calves. Appl. Anim. Behav. Sci. 1995, 43, 239–247. [Google Scholar] [CrossRef]
- Panivivat, R.; Kegley, E.B.; Pennington, J.A.; Kellogg, D.W.; Krumpelman, S.L. Growth performance and health of dairy calves bedded with different types of materials. J. Dairy Sci. 2004, 87, 3736–3745. [Google Scholar] [CrossRef]
- Higginbotham, G.E.; Stull, C.L. Performance and health of dairy calves using different types of commercial housing. Prof. Anim. Sci. 1997, 13, 18–23. [Google Scholar] [CrossRef]
- Coleman, D.A.; Moss, B.R.; McCaskey, T.A. Supplemental shade for dairy calves reared in commercial calf hutches in a southern climate. J. Dairy Sci. 1996, 79, 2038–2043. [Google Scholar] [CrossRef]
- Hart, K.A. The use of cortisol for the objective assessment of stress in animals: Pros and cons. Vet. J. 2012, 192, 137–139. [Google Scholar] [CrossRef]
- Almoosavi, S.M.M.S.; Ghoorchi, T.; Naserian, A.A.; Ramezanpor, S.S.; Ghaffari, M.H. Long-term impacts of late-gestation maternal heat stress on growth performance, blood hormones and metabolites of newborn calves independent of maternal reduced feed intake. Domest. Anim. Endocrinol. 2020, 72, 106433. [Google Scholar] [CrossRef] [PubMed]
- Veissier, I.; de la Fe, A.R.; Pradel, P. Nonnutritive oral activities and stress responses of veal calves in relation to feeding and housing conditions. Appl. Anim. Behav. Sci. 1998, 57, 35–49. [Google Scholar] [CrossRef]
- Bonelli, F.; Rota, A.; Aurich, C.; Ille, N.; Camillo, F.; Panzani, D.; Sgorbini, M. Determination of salivary cortisol in donkey stallions. J. Equine Vet. Sci. 2019, 77, 68–71. [Google Scholar] [CrossRef]
- Burnett, T.A.; Madureira, A.M.L.; Silper, B.F.; Tahmasbi, A.; Nadalin, A.; Veira, D.M.; Cerri, R.L.A. Relationship of concentrations of cortisol in hair with health, biomarkers in blood, and reproductive status in dairy cows. J. Dairy Sci. 2015, 98, 4414–4426. [Google Scholar] [CrossRef]
- Comin, A.; Prandi, A.; Peric, T.; Corazzin, M.; Dovier, S.; Bovolenta, S. Hair cortisol levels in dairy cows from winter housing to summer highland grazing. Livest. Sci. 2011, 138, 69–73. [Google Scholar] [CrossRef]
- Comin, A.; Peric, T.; Corazzin, M.; Veronesi, M.C.; Meloni, T.; Zufferli, V.; Cornacchia, G.; Prandi, A. Hair cortisol as a marker of hypothalamic-pituitary-adrenal axis activation in Friesian dairy cows clinically or physiologically compromised. Livest. Sci. 2013, 152, 36–41. [Google Scholar] [CrossRef]
- Davis, A.K.; Maney, D.L.; Maerz, J.C. The use of leukocyte profiles to measure stress in vertebrates: A review for ecologists. Funct. Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]


| Response Variable | Family Fit | Contrast | Ratio | 95% CI | t-Value |
| Cortisol | Tweedie | D3/D10 | 1.89 | 1.07–3.33 | 3.24 |
| D3/D17 | 3.49 | 1.90–6.39 | 5.94 | ||
| D3/D24 | 5.22 | 2.61–10.45 | 6.88 | ||
| D3/D31 | 6.33 | 3.12–12.85 | 7.52 | ||
| D10/D17 | 1.84 | 0.98–3.47 | 2.80 | ||
| D10/D24 | 2.76 | 1.35–5.65 | 4.10 | ||
| D10/D31 | 3.35 | 1.62–6.95 | 4.78 | ||
| D17/D31 | 1.82 | 0.85–3.89 | 2.26 | ||
| N/L | Gaussian | D3/D17 | 1.93 | 0.93–2.93 | 2.85 |
| D3/D24 | 3.04 | 1.24–4.83 | 4.83 | ||
| D3/D31 | 3.04 | 1.23–4.84 | 4.84 | ||
| D10/D17 | 1.95 | 0.94–2.96 | 2.96 | ||
| D10/D24 | 3.10 | 1.26–4.93 | 4.93 | ||
| D10/D31 | 3.10 | 1.24–4.95 | 4.95 | ||
| Running time | Tweedie-zero inflated | D3/D24 | 0.33 | 0.12–0.90 | −3.16 |
| D3/D31 | 0.38 | 0.35–1.04 | −2.72 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogi, A.; Campera, M.; Ienco, S.; Bonelli, F.; Mariti, C.; Gazzano, A. The Correlation between Play Behavior, Serum Cortisol and Neutrophil-to-Lymphocyte Ratio in Welfare Assessment of Dairy Calves within the First Month of Life. Dairy 2022, 3, 1-11. https://doi.org/10.3390/dairy3010001
Ogi A, Campera M, Ienco S, Bonelli F, Mariti C, Gazzano A. The Correlation between Play Behavior, Serum Cortisol and Neutrophil-to-Lymphocyte Ratio in Welfare Assessment of Dairy Calves within the First Month of Life. Dairy. 2022; 3(1):1-11. https://doi.org/10.3390/dairy3010001
Chicago/Turabian StyleOgi, Asahi, Marco Campera, Sara Ienco, Francesca Bonelli, Chiara Mariti, and Angelo Gazzano. 2022. "The Correlation between Play Behavior, Serum Cortisol and Neutrophil-to-Lymphocyte Ratio in Welfare Assessment of Dairy Calves within the First Month of Life" Dairy 3, no. 1: 1-11. https://doi.org/10.3390/dairy3010001
APA StyleOgi, A., Campera, M., Ienco, S., Bonelli, F., Mariti, C., & Gazzano, A. (2022). The Correlation between Play Behavior, Serum Cortisol and Neutrophil-to-Lymphocyte Ratio in Welfare Assessment of Dairy Calves within the First Month of Life. Dairy, 3(1), 1-11. https://doi.org/10.3390/dairy3010001