Size Modulation of Enzymatically Cross-Linked Sodium Caseinate Nanoparticles via Ionic Strength Variation Affects the Properties of Acid-Induced Gels



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
2.3. Size Exclusion Chromatography
2.4. Acid-Induced Gel Formation
2.4.1. pH Monitoring during Acidification
2.4.2. Small Amplitude Oscillatory Shear Rheometry
2.4.3. Forced Syneresis
2.5. Statistical Analyses
3. Results and Discussion
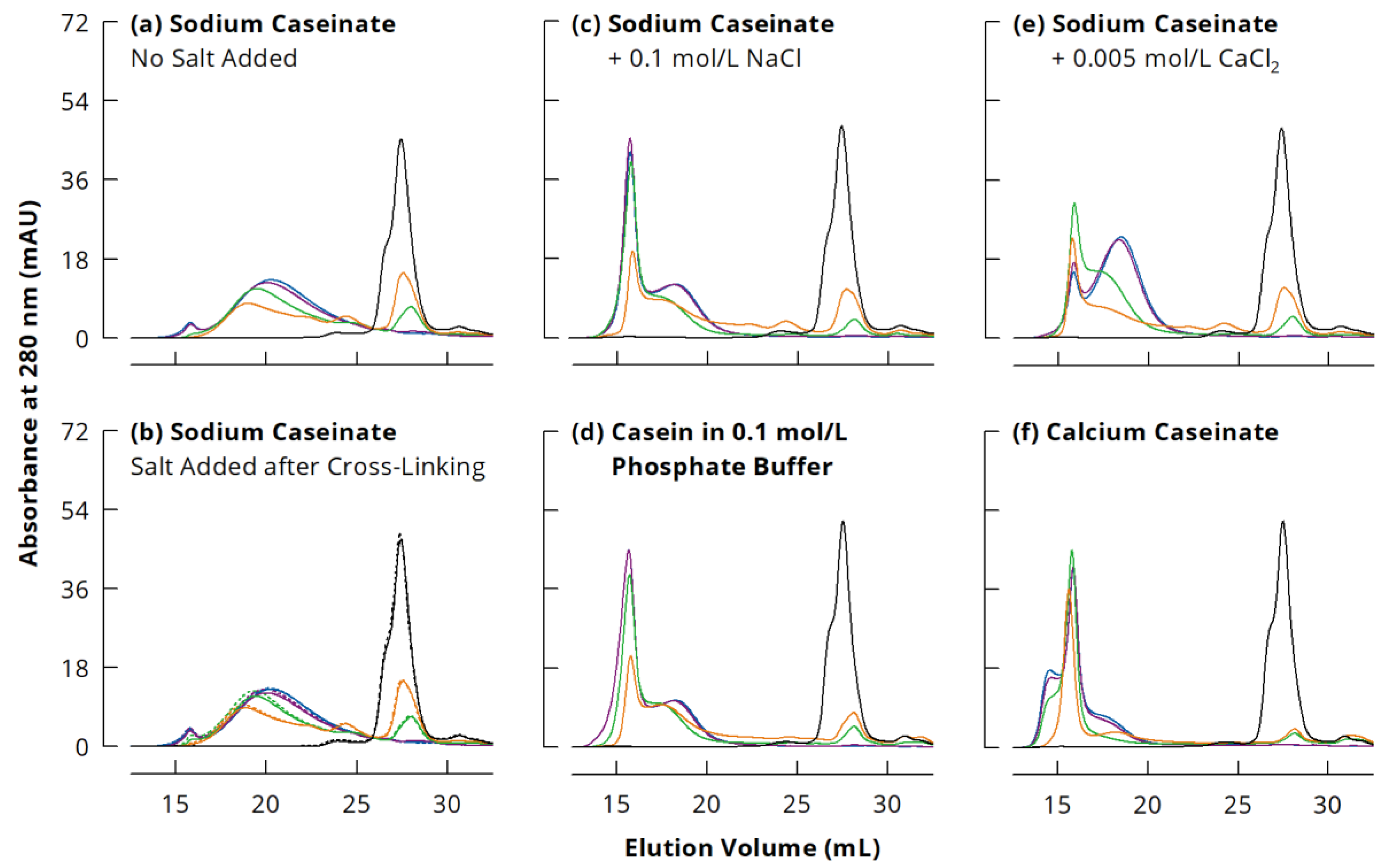
3.1. Size Modulation of Cross-Linked Sodium Caseinate Nanoparticles
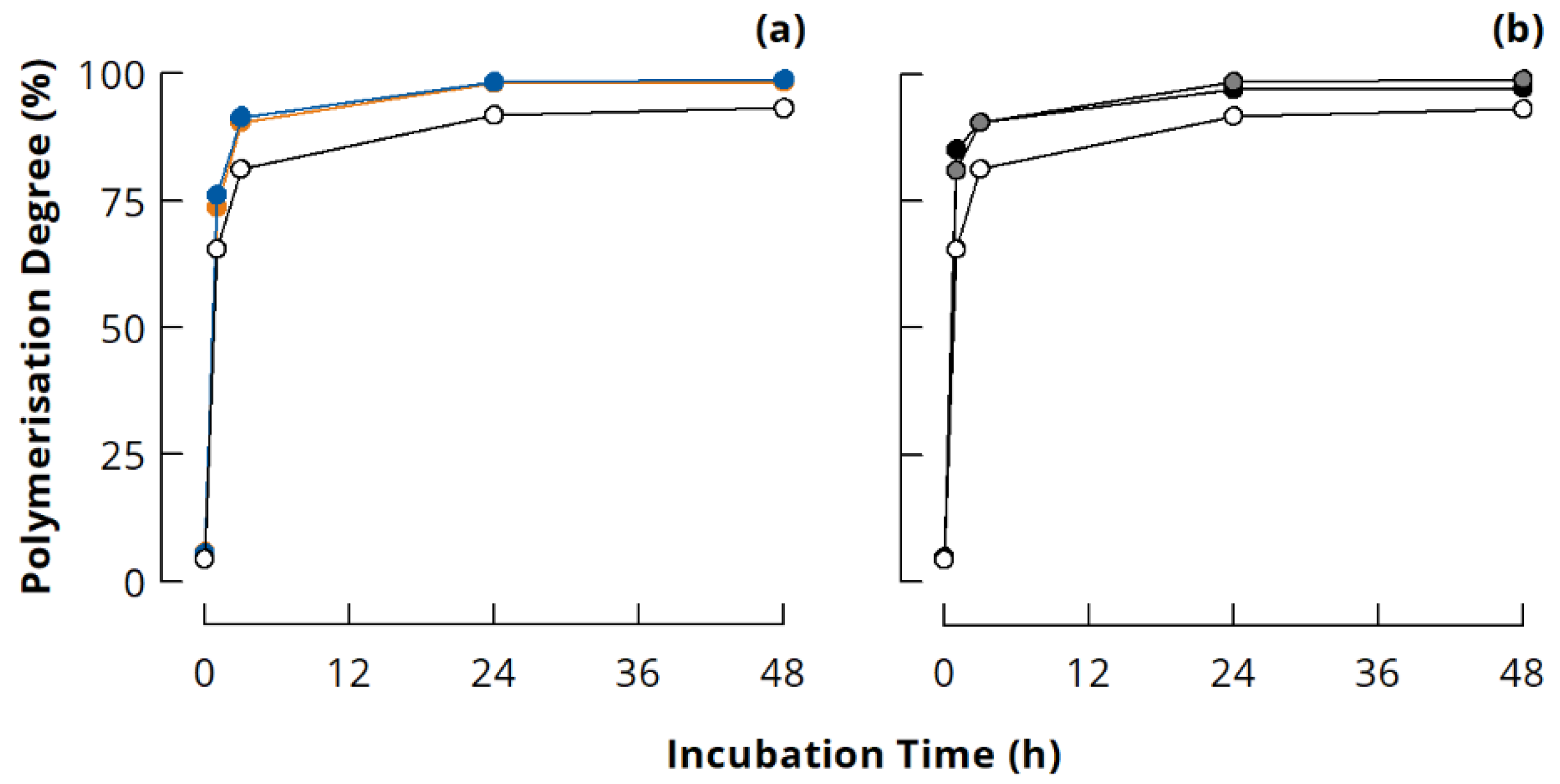
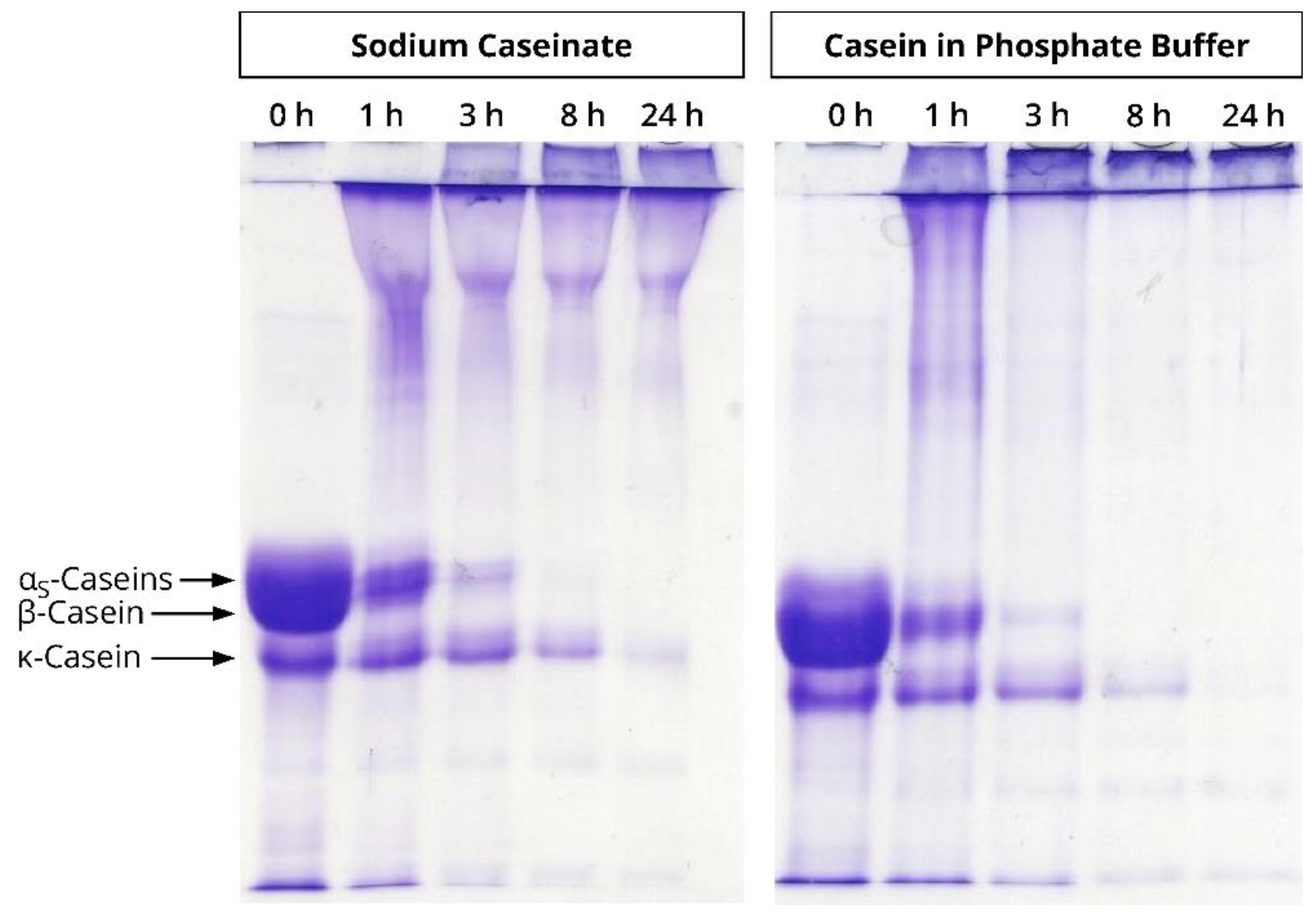
3.2. Quantitative Evaluation of Casein Polymerisation in Different Ionic Milieus
3.3. Gels from Enzymatically Cross-Linked Sodium Caseinate Nanoparticles with Different Sizes
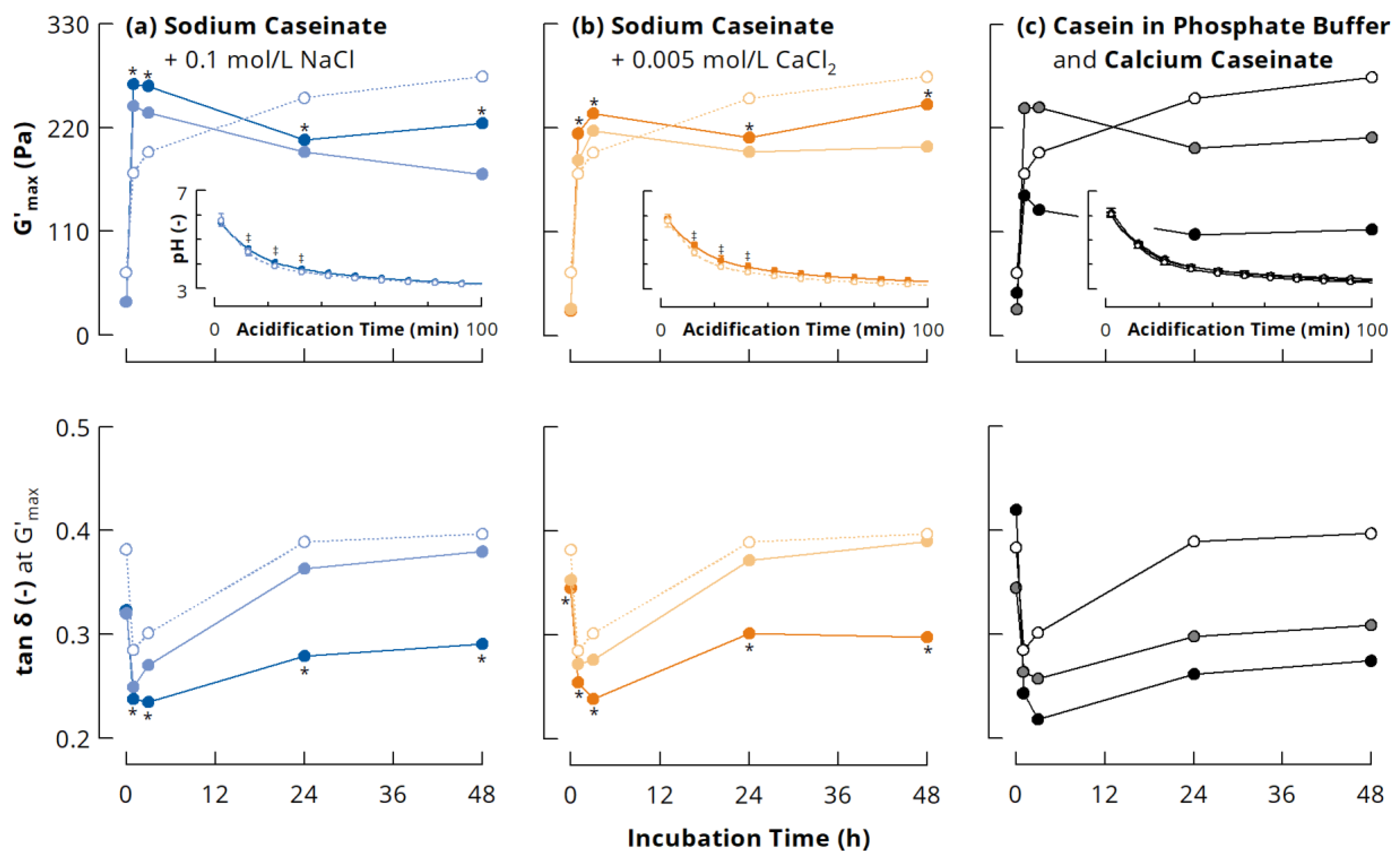
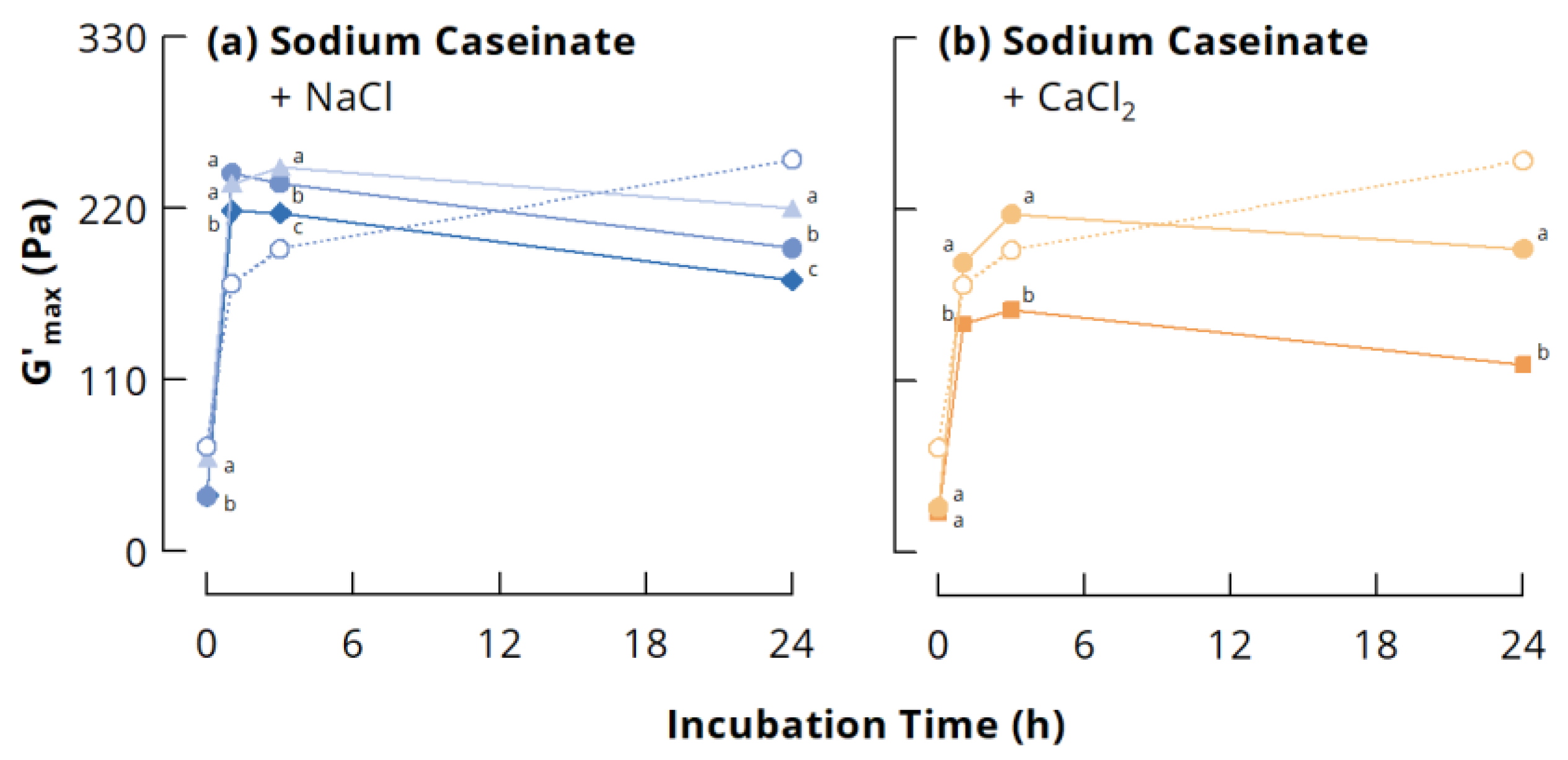
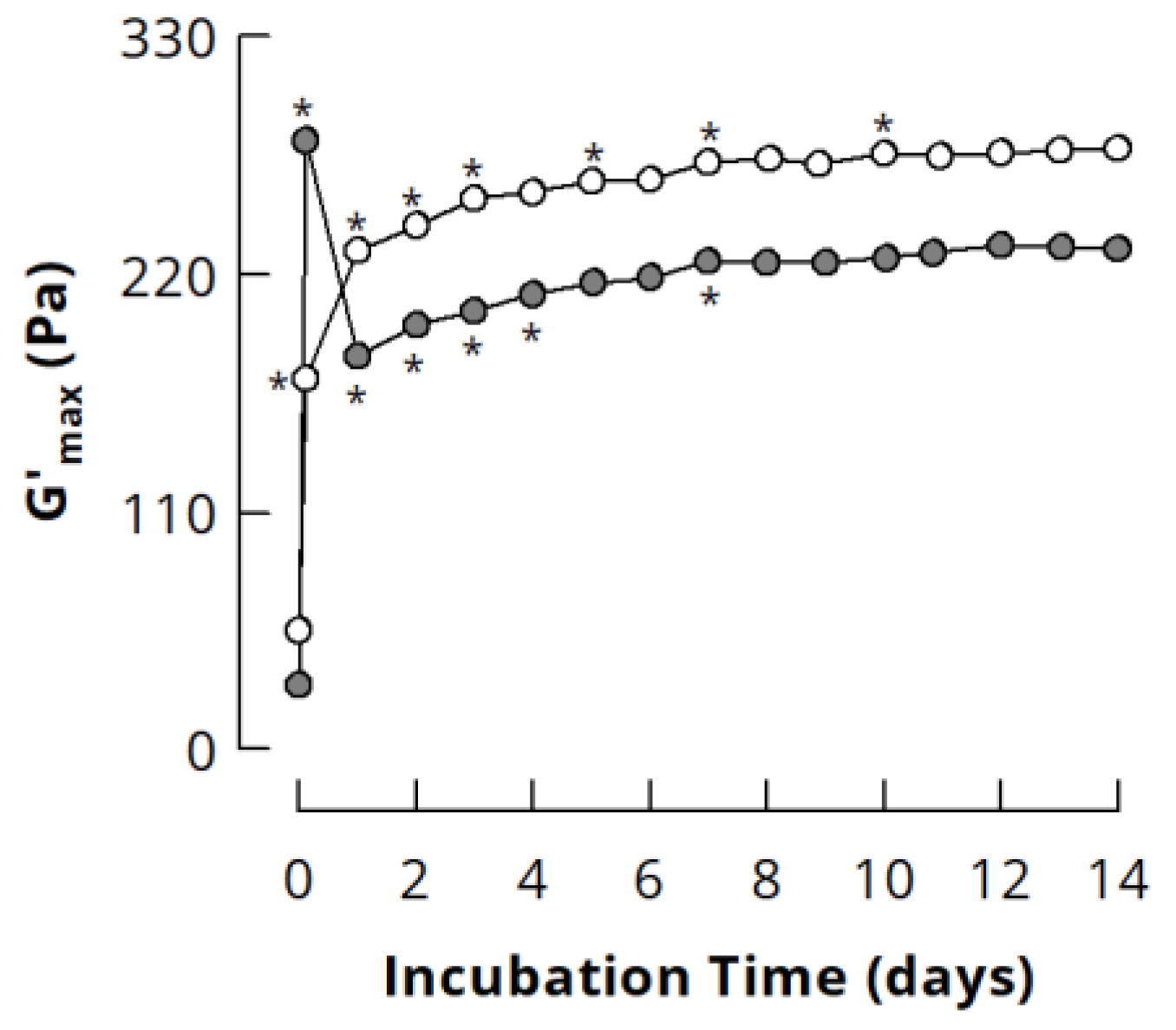
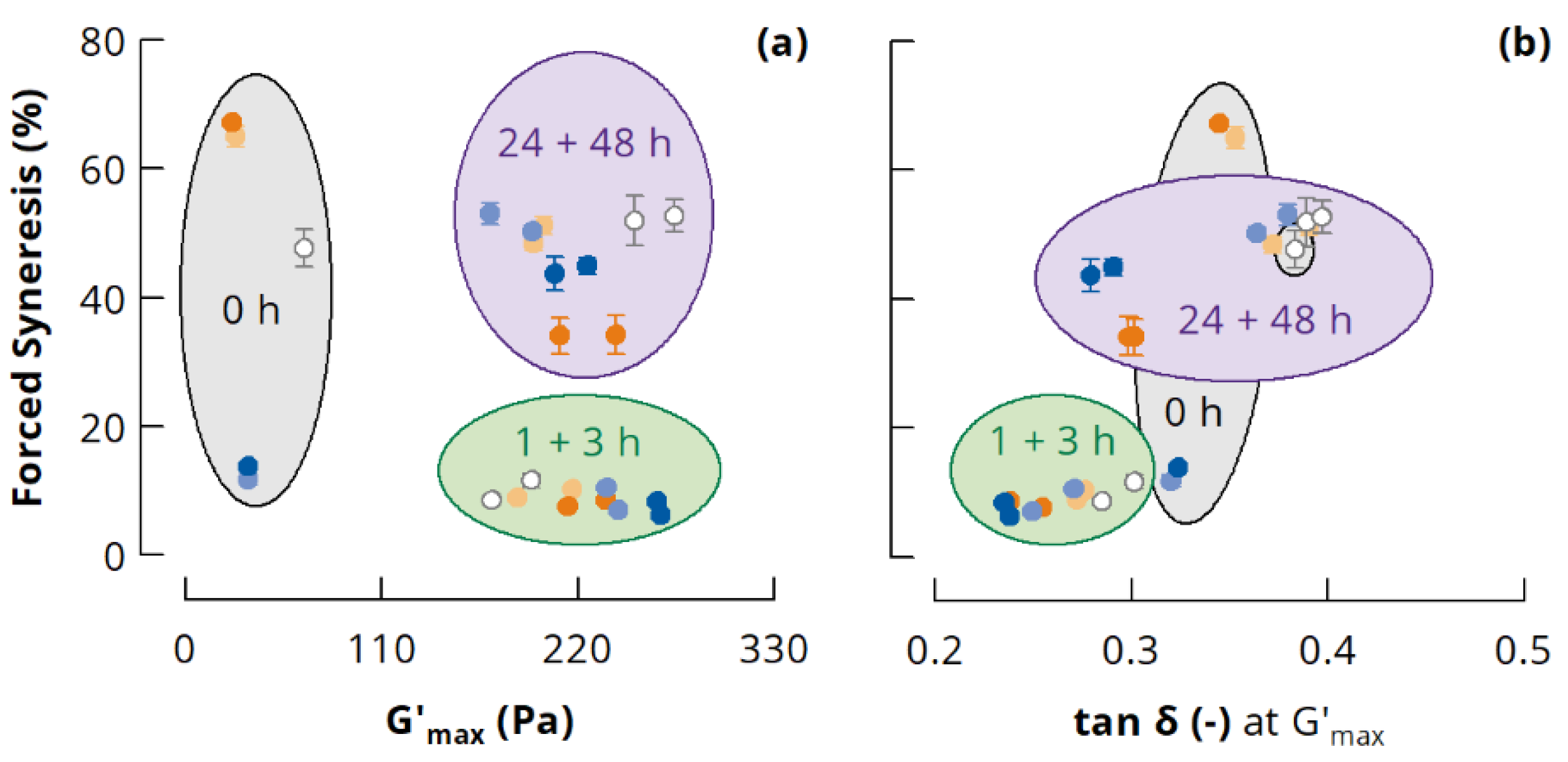
3.3.1. Small Deformation Rheology
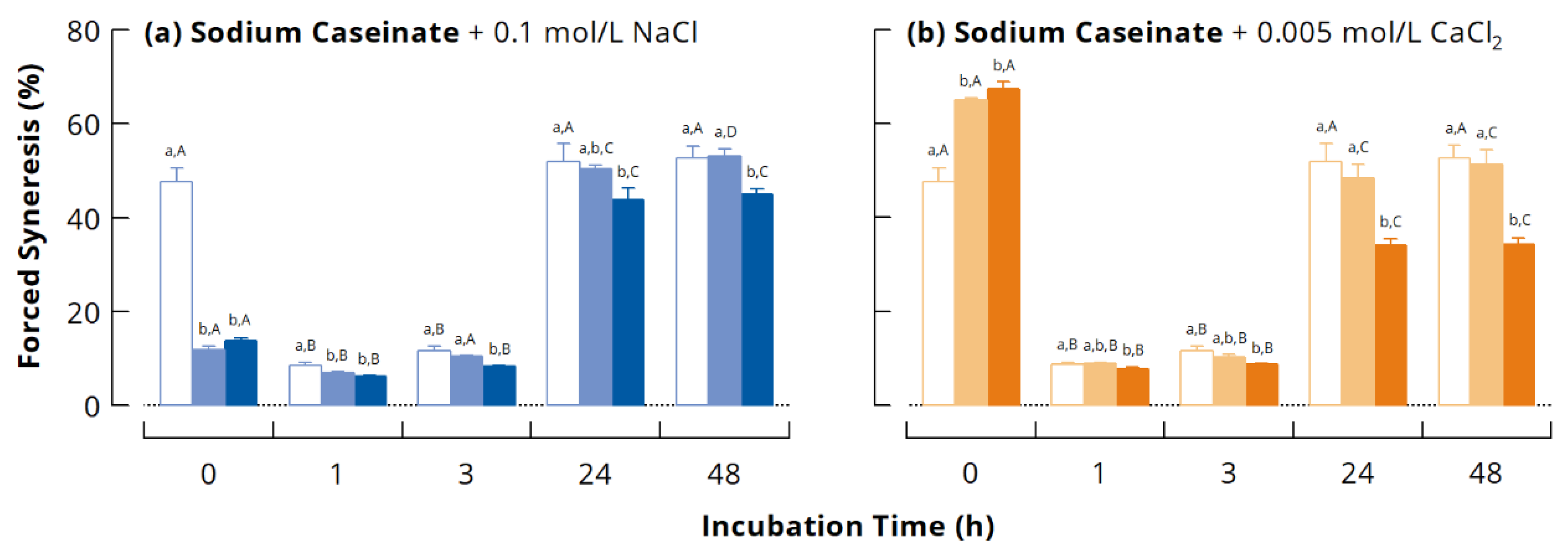
3.3.2. Forced Syneresis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Additional Experiments
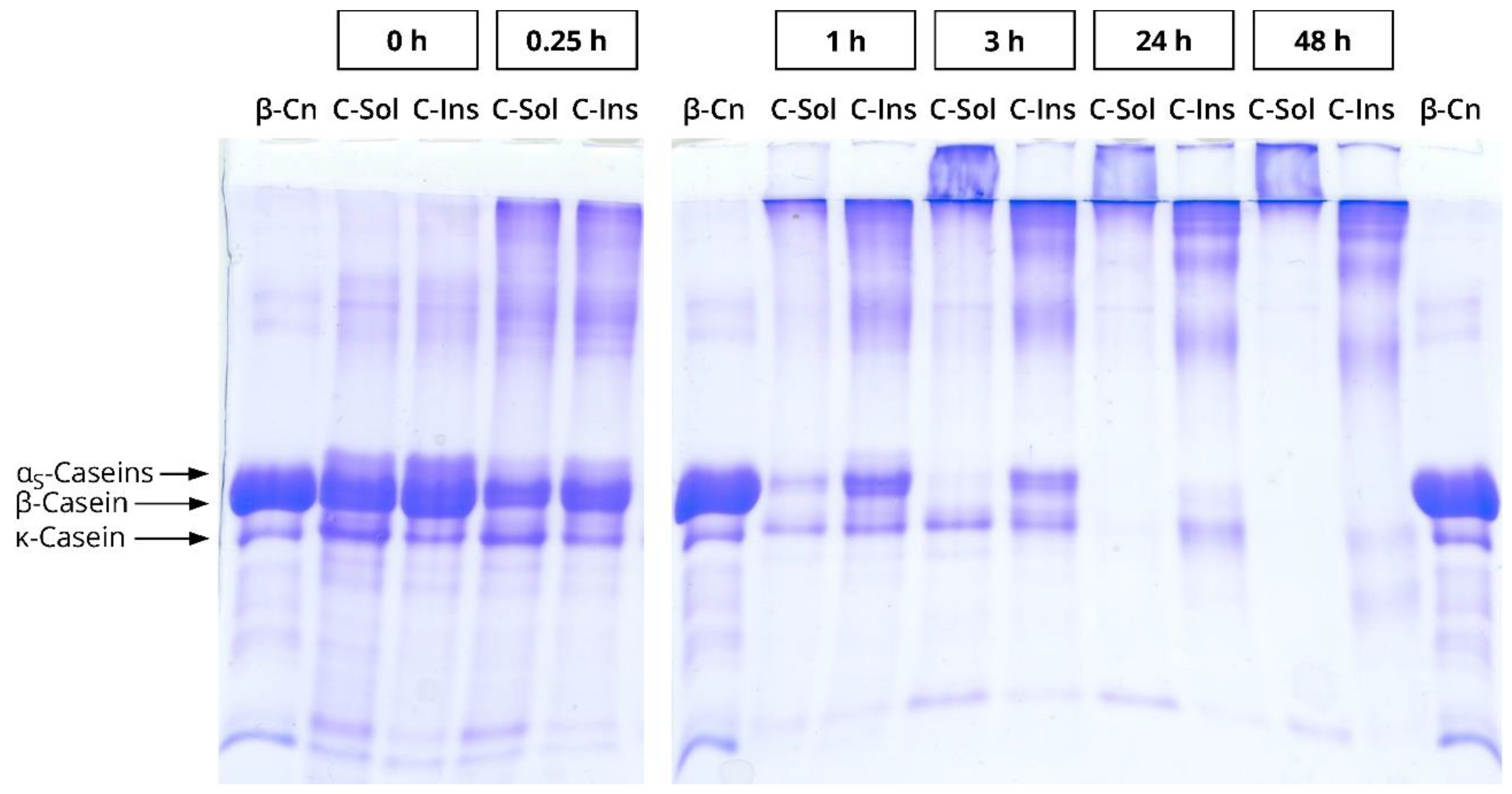
Appendix A.1. Preparation of Cold Soluble and Cold Insoluble Sodium Caseinate Fractions
Appendix A.2. Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis
Appendix B. Supplementary Figures


References
- Carr, A.; Golding, M. Functional Milk Proteins Production and Utilization: Casein-Based Ingredients. In Advanced Dairy Chemistry, 4th ed.; McSweeney, P.L.H., O’Mahony, J.A., Eds.; Springer: New York, NY, USA, 2016; Volume 1B: Proteins: Applied Aspects, pp. 35–66. ISBN 978-1-4939-2800-2. [Google Scholar] [CrossRef]
- O’Regan, J.; Mulvihill, D.M. Caseins and Caseinates, Industrial Production, Compositional Standards, Specifications, and Regulatory Aspects. In Encyclopedia of Dairy Sciences; Fuquay, J.W., Fox, P.F., McSweeney, P.L.H., Eds.; Academic Press: San Diego, CA, USA, 2011; Volume 3, pp. 855–863. ISBN 978-0-12-374407-4. [Google Scholar] [CrossRef]
- Huppertz, T.; Gazi, I.; Luyten, H.; Nieuwenhuijse, H.; Alting, A.; Schokker, E. Hydration of Casein Micelles and Caseinates: Implications for Casein Micelle Structure. Int. Dairy J. 2017, 74, 1–11. [Google Scholar] [CrossRef]
- Rohm, H.; Schmid, W. Influence of Dry-Matter Fortification on Flow Properties of Yoghurt. 1. Evaluation of Flow Curves. Milchwiss. Milk Sci. Int. 1993, 48, 556–560. [Google Scholar]
- Ünal, G.; Akalin, A.S. Influence of Fortification with Sodium-Calcium Caseinate and Whey Protein Concentrate on Microbiological, Textural and Sensory Properties of Set-Type Yoghurt. Int. J. Dairy Technol. 2013, 66, 264–272. [Google Scholar] [CrossRef]
- Lynch, A.G.; Mulvihill, D.M. Effect of Sodium Caseinate on the Stability of Cream Liqueurs. Int. J. Dairy Technol. 1997, 50, 1–7. [Google Scholar] [CrossRef]
- Singh, H. Aspects of Milk-Protein-Stabilised Emulsions. Food Hydrocoll. 2011, 25, 1938–1944. [Google Scholar] [CrossRef]
- Kizzie-Hayford, N.; Jaros, D.; Zahn, S.; Rohm, H. Effects of Protein Enrichment on the Microbiological, Physicochemical and Sensory Properties of Fermented Tiger Nut Milk. LWT Food Sci. Technol. 2016, 74, 319–324. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Chronakis, I.S. Crosslinking of Milk Proteins by Microbial Transglutaminase: Utilization in Functional Yogurt Products. Food Chem. 2018, 245, 620–632. [Google Scholar] [CrossRef] [PubMed]
- Romeih, E.; Walker, G. Recent Advances on Microbial Transglutaminase and Dairy Application. Trends Food Sci. Technol. 2017, 62, 133–140. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Koubaa, M.; Barba, F.J.; Greiner, R.; George, S.; Roohinejad, S. Recent Advances in the Application of Microbial Transglutaminase Crosslinking in Cheese and Ice Cream Products: A Review. Int. J. Biol. Macromol. 2018, 107, 2364–2374. [Google Scholar] [CrossRef] [PubMed]
- Raak, N.; Rohm, H.; Jaros, D. Enzymatic Protein Cross-Linking in Dairy Science and Technology. In Agents of Change: Enzymes in Milk and Dairy Products; Kelly, A., Larsen, L.B., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2021; pp. 417–445. ISBN 978-3-030-55481-1. [Google Scholar] [CrossRef]
- Macedo, J.A.; Fazani Cavallieri, A.L.; Lopes da Cunha, R.; Sato, H.H. The Effect of Transglutaminase from Streptomyces sp. CBMAI 837 on the Gelation of Acidified Sodium Caseinate. Int. Dairy J. 2010, 20, 673–679. [Google Scholar] [CrossRef]
- Myllärinen, P.; Buchert, J.; Autio, K. Effect of Transglutaminase on Rheological Properties and Microstructure of Chemically Acidified Sodium Caseinate Gels. Int. Dairy J. 2007, 17, 800–807. [Google Scholar] [CrossRef]
- Raak, N.; Rohm, H.; Jaros, D. Enzymatic Cross-Linking of Casein Facilitates Gel Structure Weakening Induced by Overacidification. Food Biophys. 2017, 12, 261–268. [Google Scholar] [CrossRef]
- Rohm, H.; Ullrich, F.; Schmidt, C.; Löbner, J.; Jaros, D. Gelation of Cross-Linked Casein under Small and Large Shear Strain. J. Texture Stud. 2014, 45, 130–137. [Google Scholar] [CrossRef]
- Kizzie-Hayford, N.; Jaros, D.; Rohm, H. Enrichment of Tiger Nut Milk with Microbial Transglutaminase Cross-Linked Protein Improves the Physico-Chemical Properties of the Fermented System. LWT Food Sci. Technol. 2017, 81, 226–232. [Google Scholar] [CrossRef]
- Chen, L.; Li, Y.; Han, J.; Yuan, D.; Lu, Z.; Zhang, L. Influence of Transglutaminase-Induced Modification of Milk Protein Concentrate (MPC) on Yoghurt Texture. Int. Dairy J. 2018, 78, 65–72. [Google Scholar] [CrossRef]
- Guyot, C.; Kulozik, U. Effect of Transglutaminase-Treated Milk Powders on the Properties of Skim Milk Yoghurt. Int. Dairy J. 2011, 21, 628–635. [Google Scholar] [CrossRef]
- Romeih, E.; Albadarin, A.B.; Olaleye, A.; Walker, G. Enzymatically Cross-Linked Skim Milk Powder: Enhanced Rheological and Functional Properties. Int. Dairy J. 2021, 113, 104885. [Google Scholar] [CrossRef]
- Zeeb, B.; Beicht, J.; Eisele, T.; Gibis, M.; Fischer, L.; Weiss, J. Transglutaminase-Induced Crosslinking of Sodium Caseinate Stabilized Oil Droplets in Oil-In-Water Emulsions. Food Res. Int. 2013, 54, 1712–1721. [Google Scholar] [CrossRef]
- Raak, N.; Rohm, H.; Jaros, D. Enzymatically Cross-Linked Sodium Caseinate as Techno-Functional Ingredient in Acid-Induced Milk Gels. Food Bioproc. Technol. 2020, 13, 1857–1865. [Google Scholar] [CrossRef]
- Jaros, D.; Partschefeld, C.; Henle, T.; Rohm, H. Transglutaminase in Dairy Products: Chemistry, Physics, Applications. J. Texture Stud. 2006, 37, 113–155. [Google Scholar] [CrossRef]
- Abbate, R.A.; Raak, N.; Boye, S.; Janke, A.; Rohm, H.; Jaros, D.; Lederer, A. Asymmetric Flow Field Flow Fractionation for the Investigation of Caseins Cross-linked by Microbial Transglutaminase. Food Hydrocoll. 2019, 92, 117–124. [Google Scholar] [CrossRef]
- Hannß, M.; Abbate, R.A.; Mitzenheim, E.; Alkhalaf, M.; Boehm, W.; Lederer, A.; Henle, T. Association of Enzymatically and Non-Enzymatically Functionalized Caseins Analyzed by Size-Exclusion Chromatography and Light Scattering Techniques. J. Agric. Food Chem. 2020, 68, 2773–2782. [Google Scholar] [CrossRef]
- Raak, N.; Abbate, R.A.; Alkhalaf, M.; Lederer, A.; Rohm, H.; Jaros, D. Concentration-Triggered Liquid-To-Solid Transition of Sodium Caseinate Suspensions as a Function of Temperature and Enzymatic Cross-Linking. Food Hydrocoll. 2020, 101, 105464. [Google Scholar] [CrossRef]
- HadjSadok, A.; Pitkowski, A.; Nicolai, T.; Benyahia, L.; Moulai-Mostefa, N. Characterisation of Sodium Caseinate as a Function of Ionic Strength, pH and Temperature Using Static and Dynamic Light Scattering. Food Hydrocoll. 2008, 22, 1460–1466. [Google Scholar] [CrossRef]
- Smialowska, A.; Matia-Merino, L.; Ingham, B.; Carr, A.J. Effect of Calcium on the Aggregation Behaviour of Caseinates. Coll. Surf. A 2017, 522, 113–123. [Google Scholar] [CrossRef]
- Raak, N.; Brehm, L.; Abbate, R.A.; Henle, T.; Lederer, A.; Rohm, H.; Jaros, D. Self-association of casein studied using enzymatic cross-linking at different temperatures. Food Biosci. 2019, 28, 89–98. [Google Scholar] [CrossRef]
- Raak, N.; Brehm, L.; Leidner, R.; Henle, T.; Rohm, H.; Jaros, D. Hydrolysis by Indigenous Plasmin: Consequences for Enzymatic Cross-Linking and Acid-Induced Gel Formation of Non-Micellar Casein. Food Biophys. 2020, 15, 32–41. [Google Scholar] [CrossRef]
- Raak, N.; Schöne, C.; Rohm, H.; Jaros, D. Acid-Induced Gelation of Enzymatically Cross-Linked Caseinate in Different Ionic Milieus. Food Hydrocoll. 2019, 86, 43–49. [Google Scholar] [CrossRef]
- International Dairy Federation. Caseins and Caseinates—Determination of Protein Content (Reference Method); IDF Standard 92: Brussels, Belgium, 1979. [Google Scholar]
- Folk, J.E.; Cole, P.W. Structural Requirements of Specific Substrates for Guinea Pig Liver Transglutaminase. J. Biol. Chem. 1965, 240, 2951–2960. [Google Scholar] [CrossRef]
- Folk, J.E.; Cole, P.W. Mechanism of Action of Guinea Pig Liver Transglutaminase. I. Purification and Properties of the Enzyme: Identification of a Functional Cysteine Essential for Activity. J. Biol. Chem. 1966, 241, 5518–5525. [Google Scholar] [CrossRef]
- Jaros, D.; Jacob, M.; Otto, C.; Rohm, H. Excessive Cross-Linking of Caseins by Microbial Transglutaminase and Its Impact on Physical Properties of Acidified Milk Gels. Int. Dairy J. 2010, 20, 321–327. [Google Scholar] [CrossRef]
- Raak, N.; Abbate, R.A.; Lederer, A.; Rohm, H.; Jaros, D. Size Separation Techniques for the Characterisation of Cross-Linked Casein: A Review of Methods and Their Applications. Separations 2018, 5, 14. [Google Scholar] [CrossRef]
- Macierzanka, A.; Bordron, F.; Rigby, N.M.; Mills, E.N.C.; Lille, M.; Poutanen, K.; Mackie, A.R. Transglutaminase Cross-Linking Kinetics of Sodium Caseinate is Changed after Emulsification. Food Hydrocoll. 2011, 25, 843–850. [Google Scholar] [CrossRef]
- Bönisch, M.P.; Lauber, S.; Kulozik, U. Improvement of Enzymatic Cross-Linking of Casein Micelles with Transglutaminase by Glutathione Addition. Int. Dairy J. 2007, 17, 3–11. [Google Scholar] [CrossRef]
- Kizzie-Hayford, N.; Raak, N.; Jaros, D.; Rohm, H. Addition of Crude Tiger Nut Protein Extract Affects Stiffness of Enzymatically Cross-Linked Dairy Proteins. Int. J. Food Sci. Technol. 2018, 53, 1865–1870. [Google Scholar] [CrossRef]
- Moeckel, U.; Duerasch, A.; Weiz, A.; Ruck, M.; Henle, T. Glycation Reactions of Casein Micelles. J. Agric. Food Chem. 2016, 64, 2953–2961. [Google Scholar] [CrossRef] [PubMed]
- Hagel, L. Gel Filtration: Size Exclusion Chromatography. In Protein Purification: Principles, High Resolution Methods, and Applications, 3rd ed.; Janson, J.-C., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 51–91. ISBN 978-0-470-93993-2. [Google Scholar] [CrossRef]
- Menéndez, O.; Schwarzenbolz, U.; Rohm, H.; Henle, T. Casein Gelation under Simultaneous Action of Transglutaminase and Glucono-δ-Lactone. Food Nahr. 2004, 48, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Raak, N.; Gehrisch, S.; Rohm, H.; Jaros, D. Rotational Thromboelastometry for Characterising Acid-Induced Gelation of Cross-Linked Casein. Food Biophys. 2015, 10, 25–29. [Google Scholar] [CrossRef]
- Lucey, J.A.; Srinivasan, M.; Singh, H.; Munro, P.A. Characterization of Commercial and Experimental Sodium Caseinates by Multiangle Laser Light Scattering and Size-Exclusion Chromatography. J. Agric. Food Chem. 2000, 48, 1610–1616. [Google Scholar] [CrossRef] [PubMed]
- Parker, E.A.; Donato, L.; Dalgleish, D.G. Effects of Added Sodium Caseinate on the Formation of Particles in Heated Milk. J. Agric. Food Chem. 2005, 53, 8265–8272. [Google Scholar] [CrossRef]
- Jaros, D.; Schwarzenbolz, U.; Raak, N.; Löbner, J.; Henle, T.; Rohm, H. Cross-Linking with Microbial Transglutaminase: Relationship between Polymerisation Degree and Stiffness of Acid Casein Gels. Int. Dairy J. 2014, 38, 174–178. [Google Scholar] [CrossRef]
- Jaros, D.; Schwarzenbolz, U.; Raak, N.; Löbner, J.; Henle, T.; Rohm, H. Corrigendum to “Cross-Linking with Microbial Transglutaminase: Relationship between Polymerisation Degree and Stiffness of Acid Casein Gels” [Int Dairy J 38 (2014) 174–178]. Int. Dairy J. 2014, 39, 345–347. [Google Scholar] [CrossRef]
- Lucey, J.A.; van Vliet, T.; Grolle, K.; Geurts, T.; Walstra, P. Properties of Acid Casein Gels Made by Acidification with Glucono-δ-Lactone. 1. Rheological Properties. Int. Dairy J. 1997, 7, 381–388. [Google Scholar] [CrossRef]
- Matia-Merino, L.; Lau, K.; Dickinson, E. Effects of Low-Methoxyl Amidated Pectin and Ionic Calcium on Rheology and Microstructure of Acid-Induced Sodium Caseinate Gels. Food Hydrocoll. 2004, 18, 271–281. [Google Scholar] [CrossRef]
- Raak, N.; Rohm, H.; Jaros, D. Cross-linking with Microbial Transglutaminase: Isopeptide Bonds and Polymer Size as Drivers for Acid Casein Gel Stiffness. Int. Dairy J. 2017, 66, 49–55. [Google Scholar] [CrossRef]
- Saricay, Y.; Wierenga, P.A.; de Vries, R. Rheological Properties of Dispersions of Enzymatically Cross-Linked Apo-α-Lactalbumin. Food Hydrocoll. 2016, 56, 344–351. [Google Scholar] [CrossRef]
- Raak, N. Cross-Linking with Microbial Transglutaminase: Drivers of Structure-Function-Interrelations in Acid Caseinate Gels. Ph.D. Thesis, Technische Universität Dresden, Dresden, Germany, 8 July 2019. Available online: https://nbn-resolving.org/urn:nbn:de:bsz:14-qucosa2-361633 (accessed on 12 September 2020).
- Hannß, M.; Böhm, W.; Drichel, S.; Henle, T. Acid-Induced Gelation of Enzymatically and Nonenzymatically Cross-Linked Caseins—Texture Properties, and Microstructural Insights. J. Agric. Food Chem. 2020, 68, 13970–13981. [Google Scholar] [CrossRef] [PubMed]
- Moschakis, T.; Murray, B.S.; Dickinson, E. On the Kinetics of Acid Sodium Caseinate Gelation Using Particle Tracking to Probe the Microrheology. J. Coll. Interface Sci. 2010, 345, 278–285. [Google Scholar] [CrossRef]
- Lucey, J.A.; van Vliet, T.; Grolle, K.; Geurts, T.; Walstra, P. Properties of Acid Casein Gels Made by Acidification with Glucono-δ-Lactone. 2. Syneresis, Permeability and Microstructural Properties. Int. Dairy J. 1997, 7, 389–397. [Google Scholar] [CrossRef]
- Ercili-Cura, D.; Lille, M.; Legland, D.; Gaucel, S.; Poutanen, K.; Partanen, R.; Lantto, R. Structural Mechanisms Leading to Improved Water Retention in Acid Milk Gels by Use of Transglutaminase. Food Hydrocoll. 2013, 30, 419–427. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Type | Ionic Milieu Prior to Cross-Linking 1 | Cross-Linking Conditions | Salt Added After Cross-Linking | Approx. Ionic Strength 2 |
|---|---|---|---|---|
| Sodium Caseinate | 0.022 M NaOH | 40 °C, 3 U/gProtein, 0–48 h | - | 0.015 M |
| 30 °C, 3 U/gProtein, 0–14 days | - | 0.015 M | ||
| 40 °C, 3 U/gProtein, 0–24 h | 0.050 M NaCl | 0.065 M | ||
| 40 °C, 3 U/gProtein, 0–48 h | 0.100 M NaCl | 0.115 M | ||
| 40 °C, 3 U/gProtein, 0–24 h | 0.150 M NaCl | 0.165 M | ||
| 40 °C, 3 U/gProtein, 0–48 h | 0.005 M CaCl2 | 0.030 M | ||
| 40 °C, 3 U/gProtein, 0–24 h | 0.015 M CaCl2 | 0.060 M | ||
| 0.022 M NaOH 0.1 M NaCl | 40 °C, 3 U/gProtein, 0–48 h | - | 0.115 M | |
| 0.022 M NaOH 0.005 M CaCl2 | 40 °C, 3 U/gProtein, 0–48 h | - | 0.030 M | |
| Casein in Phosphate Buffer | 0.07 M NaH2PO4 0.03 M Na2HPO4 (pH 6.6) | 40 °C, 3 U/gProtein, 0–48 h | - | 0.165 M |
| 30 °C, 3 U/gProtein, 0–14 days | - | 0.165 M | ||
| Calcium Caseinate | 0.013 M Ca(OH)2 | 40 °C, 3 U/gProtein, 0–48 h | - | 0.030 M |
| Study | Elution Buffer | First Eluting Particle Species | Second Eluting Particle Species | Casein Monomers |
|---|---|---|---|---|
| [36] | Denaturing and reducing | ~7–9 | ~9–13 | ~15–17 |
| [44] | Native | ~6–8.5 | ~8.5–12 | ~12–17 |
| [25] | Native | ~7.5–10 | ~10–15 | ~15–20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raak, N.; Leonhardt, L.; Rohm, H.; Jaros, D. Size Modulation of Enzymatically Cross-Linked Sodium Caseinate Nanoparticles via Ionic Strength Variation Affects the Properties of Acid-Induced Gels. Dairy 2021, 2, 148-164. https://doi.org/10.3390/dairy2010014
Raak N, Leonhardt L, Rohm H, Jaros D. Size Modulation of Enzymatically Cross-Linked Sodium Caseinate Nanoparticles via Ionic Strength Variation Affects the Properties of Acid-Induced Gels. Dairy. 2021; 2(1):148-164. https://doi.org/10.3390/dairy2010014
Chicago/Turabian StyleRaak, Norbert, Lars Leonhardt, Harald Rohm, and Doris Jaros. 2021. "Size Modulation of Enzymatically Cross-Linked Sodium Caseinate Nanoparticles via Ionic Strength Variation Affects the Properties of Acid-Induced Gels" Dairy 2, no. 1: 148-164. https://doi.org/10.3390/dairy2010014
APA StyleRaak, N., Leonhardt, L., Rohm, H., & Jaros, D. (2021). Size Modulation of Enzymatically Cross-Linked Sodium Caseinate Nanoparticles via Ionic Strength Variation Affects the Properties of Acid-Induced Gels. Dairy, 2(1), 148-164. https://doi.org/10.3390/dairy2010014