Abstract
Exosomes are membrane-enveloped nanosized (30–150 nm) extracellular vesicles of endosomal origin produced by almost all cell types and encompass a multitude of functioning biomolecules. Exosomes have been considered crucial players of cell-to-cell communication in physiological and pathological conditions. Accumulating evidence suggests that exosomes can modulate the immune system by delivering a plethora of signals that can either stimulate or suppress immune responses, which have potential applications as immunotherapies for cancer and autoimmune diseases. Here, we discuss the current knowledge about the active biomolecular components of exosomes that contribute to exosomal function in modulating different immune cells and also how these immune cell-derived exosomes play critical roles in immune responses. We further discuss the translational potential of engineered exosomes as immunotherapeutic agents with their advantages over conventional nanocarriers for drug delivery and ongoing clinical trials.
1. Introduction
Cell to cell communications between neighboring and distant cells are fundamental for cell survival and also essential for maintaining physiological homeostasis. Furthermore, intercellular communication is crucial for the pathophysiology of several diseases, especially for tumorigenesis. In the past decade, with extensive research, exosomes emerged as novel mediators of intercellular communication in physiological and pathological conditions [1,2].
Exosomes are small endosomal origin extracellular vesicles (EVs) with a lipid bilayer, 30–150 nm, secreted by almost all cell types in the extracellular space such as blood, and travel to distant tissues [3,4,5]. According to the recently updated guidelines of the International Society for Extracellular Vesicles (ISEV) on minimal information for studies of extracellular vesicles (MISEV), the term “extracellular vesicle” or “EV” has now been agreed on as the consensus generic term for lipid bilayer-delimited particles released from the cell and cannot replicate [6]. They carry various biologically active macromolecules such as proteins, RNA, DNA, miRNAs and metabolites, which act as mediators in cell-to-cell communication [7,8,9]. Exosomes are formed by endocytosis of the endosome. In the process, there is endocytosis and scission of invaginated plasma membrane leading to the formation of endosomes. With the help of the Golgi complex, these newly formed vesicles turn from early endosomes to late endosomes [10,11,12,13]. The resulting endosome creates intraluminal vesicles (ILVs) in the lumen of the endosome through several invaginations of the membrane [14]. This process of invagination of the endosomal membrane in the lumen is controlled by endosomal sorting complexes required for transport (ESCRT) [15]. These proteins assist in sorting endosomal proteins and cell contents in ILVs, leading to the formation of mature endosomes called multivesicular bodies (MVBs) [16,17]. These MVBs then fuse with the cell membrane and later release their internal vesicles, i.e., exosomes, in extracellular space. Exosomes are differentiated based on their size, exosomal markers, RNA, and other special proteins [18].
The exosomes are involved in multiple different pathways affecting multiple systems. Research has shown their involvement in activating the immune system through different pathways. They include increasing cell activity and enhancing the release of natural killer cells, tumor necrosis cells, inducing macrophages [19,20,21,22,23]. Furthermore, they also play a role in signal transfer among numerous types of neurons, although the mechanism is still unclear [24,25]. They are also heavily involved in cell proliferation by carrying contents from parental cells and acting as stem cells mediators.
Given their abundance and biological properties, they can be utilized as a potential diagnostic and therapeutic tool. In this review, we summarize the various functional biomolecular components of exosomes and outline the exosome-mediated regulation of the immune system. Moreover, we also discuss the clinical implications of exosomes in immunotherapy with their advantages and ongoing clinical trials.
2. Isolation, Purification and Characterization of Exosomes
Exosomes have been isolated from a variety of bodily fluids including bile, blood, breast milk, urine, cerebrospinal fluid, and saliva as well as from in vitro culture media [26]. Although exosomes have enormous prospects for clinical use as therapeutic, diagnostic, and prognostic probes, the progress is somewhat subdued by our inadequate knowledge of the most efficient and reproducible approach for large-scale production of quality exo-somes in a short time. Several isolation techniques have been developed by exploiting a particular trait, such as the size, density and surface markers of exosomes which includes ultracentrifugation, density gradient centrifugation, ultrafiltration, an immunoaffinity capture-based method, microfluidics-based techniques and a polyethylene glycol (PEG)-based isolation method [27,28,29]. Up to the present, the most widely adopted and reliable method is ultracentrifugation, which involves a series of centrifugation steps to remove cells, large vesicles, debris and precipitate exosomes. But ultracentrifugation method is tedious, time-consuming that needs special equipment and unable to separate impurities like viruses, apoptotic bodies and proteins. Commercially available polymeric precipitation mixtures have been widely applied to isolate exosomes as it is a quick and high yielding technique, albeit it is inferior to ultracentrifugation because of co-isolation of contaminants and compatibility with downstream analysis. Although immunoaffinity capture-based methods have the potential to isolate selective subtypes of exosomes (e.g., CD9, CD63, CD81, pCAM), a substantial population is left out and its not applicable for larger samples. Microfluidics-based techniques are highly efficient, portable and cost-effective but they have a low sample capacity [30].
These techniques are not always mutually exclusive, and their combinations may be more reasonable for better yield [31]. The International Society for Extracellular Vesicles (ISEV) provided Minimal information for studies of extracellular vesicles 2018 (MISEV2018) guidelines based on evolution of the collective knowledge to guide and improve the field [6]. Many technical challenges need to be addressed in the development of exosome-based therapeutics. The most crucial factor in their therapeutic application is the large-scale production of high-quality exosomes in a shorter time. The tangential flow filtration (TFF) has been proposed as the ideal method for industrial-scale manufacturing of exosomes and is thought to have a superior yield and functionality of exosomes compared with those isolated by ultracentrifugation [32,33,34]. Although using TFF with appropriate pore sizes and parameters high-purity isolation of exosomes is feasible, delicate optimizations of TFF are indispensable to preserve the surface-associated functional proteins of exosomes [35].
Assessment of the physicochemical and biomolecular properties of exosomes, such as size, shape, surface charge, density, and porosity, molecular contents is fundamental to determine their biological function which warrants precise confirmation of these characteristics for future applications of the exosome [36]. Several methods have been utilized to characterize and validate these exosomes for both research and clinical purposes. These methods include western blot, flow cytometry, enzyme-linked immunosorbent assay (ELISA), polymerase chain reaction (PCR), nanoparticle tracking assay(NTA), electron microscopy, immunofluorescence, atomic force microscopy (AFM), dynamic light scattering (DLS), nucleic acid sequencing, mass spectrometry, single-photon emission computed tomography (SPECT), multispectral optical imaging, magnetic resonance imaging (MRI), tunable resistive pulse sensing (TRPS), etc. [36,37]. However, each of these techniques comes with its own drawbacks that must be taken into careful consideration for downstream application.
3. Biomolecular Components of Exosomes in Immunomodulation and Exosome-Mediated Regulation of Immune Cells
Biomolecular components of exosomes play crucial role in modulating immune system and regulating functions of immune cells. A summary of these molecules along with mechanisms and effects can be found in Figure 1 and Table 1.
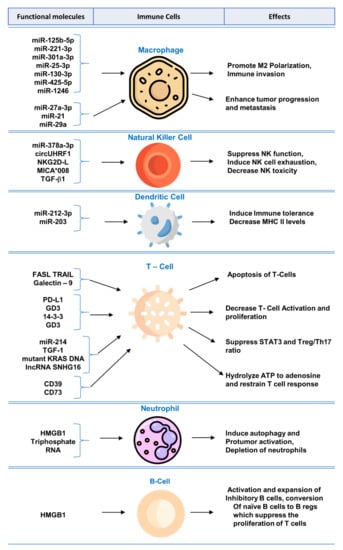
Figure 1.
Functional molecules in tumor-derived exosomes for immunomodulation and exosome-mediated regulation of immune cells.

Table 1.
Biomolecular components of EVs/exosomes for immunomodulation.
3.1. Exosomal Biomolecules Modulating T-Cell Function
Tumor-derived exosomes express multiple functional biomolecules that have been shown to impair T lymphocyte function, such as by induction of T cell apoptosis [38], inhibition of activation [46], or by suppression of function [44,45].
Oral squamous cell carcinoma-derived exosomes expressing FasL have been studied to show induction of apoptosis of T cells via receptor and mitochondrial pathways [40]. FasL-bearing exosomes also correlated with poor prognosis and nodal involvement [40]. Prostatic cancer cell line-derived exosomes carrying FasL showed dose-dependent apoptosis of CD8+ T cells and inhibited T cell proliferation in co-culture [38]. The addition of anti-FasL antibody prevented the apoptosis of CD8+ T cells by tumor-derived exosomes [38]. Melanosomes containing exosomes derived from melanoma cells have been studied to show FasL apoptosis of Jurkat cells, an immortalized line of T cells [39]. In another study, FasL and TNF-related apoptosis-inducing ligand (TRAIL) bearing microvesicles from colorectal cells showed induction of T cell apoptosis both in vivo and in vitro [41]. The study of membrane fragments derived from ovarian carcinoma suggested the association of FasL with apoptosis of T lymphocytes [42]. Interestingly, human B cell-derived lymphoblastoid cell lines (LCL) have no detectable FasL on cell membranes but induce apoptosis in CD4+ T cells by the production of MHCII+FasL+ exosome [43].
Metastatic melanoma has been shown to release exosomes carrying PD-L1 on its surface, upregulated by IFN-γ, which suppress CD8+ T Cells and promote tumor proliferation and malignant progression [44,45]. Circulating PD-L1+ exosomes contributed to poor outcome and immunosuppressive tumor microenvironment in head and neck squamous cell carcinomas (HNSCC) patients, whereas soluble PD-L1 was not associated with disease progression [46]. Similarly, in patients with NSCLC, exosomal PD-L1 has been linked to disease advancement but not soluble PD-L1 [65]. PD-L1+ exosomes derived from breast cancer cells have demonstrated transfer of PDL1 from PD-L1-positive to PD-L1-negative breast cancer cells, inhibiting T cell activation and T cell granzyme B secretion, further contributing to immunosuppressive tumor microenvironment. [47] Furthermore, inhibition of exosome PD-L1 secretion via pharmacologic and genetic measures exhibited enhancement of anti-tumor immunity in mice models [47].
14-3-3ζ, a family of phospho-serine binding proteins, overexpression has been linked to cancer progression and indicates poor prognosis and chemoresistance in multiple tumor types [66]. Hepatocellular carcinoma (HCC) cells have been shown to release exosomes carrying 14-3-3ζ which were transferred to T cells infiltrating the tumor microenvironment inhibiting their antitumor effects [49].
Colorectal cancer stem cells (CRCSCs) have shown exosome-mediated stem-like property expansion via exosomal miRNA-146a-5p (miR-146a) which promoted tumorigenicity and reduced the number of tumor-infiltrating CD8(+) T cells [52].
Moreover, tumor-derived exosomes have also shown to affect immunosuppressive Treg numbers in the tumor microenvironment [48]. Malignant effusion-derived exosomes, expressing TGF-β1 on the surface, co-cultured with Tregs showed maintenance of Treg numbers and suppressive functions [48]. It has been observed that tumor cells release microvesicles bearing miR-214 and other tumor-specific miRNAs which are taken up by peripheral CD4+ T cells resulting in downregulation of phosphatase and tensin homolog (PTEN) and promotion of Treg numbers in tumor microenvironment [53]. It has also been studied that delivery of anti- miR-214 antisense oligonucleotides (ASOs) via microvesicles inhibited Treg expansion and tumor proliferation [53].
Tumor-associated macrophage-derived exosomal transfer of miR-29a-3p and miR-21-5p have been implicated in generation of immune-suppressive microenvironment, via suppression of STAT3 and imbalance of Treg/Th17 ratio [67]. Melanoma cells have been observed to release exosomes bearing microRNAs, such as miR-690, which lead to activation of the mitochondrial apoptotic pathway in CD4+ T cells [68]. Breast cancer cells shed exosomes that transmit SNHG16 causing activation of the TGF-β1/SMAD5 pathway by sponging miR-16–5p leading to conversion of γδ1 T cells into the CD73+ immunosuppressive subtype [69]. Moreover, it has been hypothesized that stimulated T cells release exosomes containing miRNAs such as miR-21-5p, miR-16-5p, miR-155-5p, which modulate other T cells in paracrine environment [70]. MicroRNAs, such as miR-335, are transferred unidirectionally from activated T cells to antigen-presenting cells via exosomes [71].
3.2. Exosomal Biomolecules Modulating NK Cell Function
NK cells are an important component of the innate immune system; they identify MHC-I-lacking tumor cells, bind to stress-induced ligands on tumor cells, and become activated to kill tumor cells [72]. Tumor-derived exosomes have been shown to stimulate [21] or inhibit [57] the activity of NK cells. Inhibition of NK cell activation via tumor-derived exosomes leads to the escape of tumor cells from NK cell immune surveillance [59]. Tumor-derived exosomes have been shown to inhibit NK cells by blocking their activation via IL-2 [73]. Tumor-derived exosomes carrying MICA*008 molecules target NK cells and downregulate surface NKG2D receptors and minimize NK cytotoxicity leading to immune evasion [54]. Mesothelioma-derived exosomes expressing TGFβ1 and NKG2DL impair NK cell activation by downregulating surface NKG2D on NK cells [57]. Similar mode of impairment of NK cell function via immunosuppressive exosomes derived from leukemia/lymphoma T- and B-cell lines have been studied [58]. Exosomes isolated from pancreatic ductal adenocarcinoma expressing TGF-β1 demonstrated impairment of NK cell function by diminishing expression of NKG2D, CD107a, TNF-α, and INF-γ, also showed to impair glucose uptake ability by NK cells [59]. TGF-β1 inhibits the activation and function of NK cells through the TGFβ-Smad2/3 signaling pathway [59].
On the contrary, tumor-derived exosomes have also shown to stimulate NK cell function in some studies [55]. Multiple myeloma cell-derived, HSP70 bearing exosomes stimulate IFNγ production by NK cells via TLR2/HSP70 activation of NF-κB pathway [55]. IFN-γ drives anti-tumor immunity but at the same time upregulates expression of PD-L1 on tumor and myeloid cells resulting in immune tolerance [74]. Additionally, melanoma cells overexpressing HSP70 have shown to release HSP70-positive exosomes which activated NK cells, which then preferentially kill NKG2D ligand or MICA/B expressing tumor cells in mice studies [56]. Furthermore, exosomes expressing HSP70/Bag4, derived from pancreatic and colon cancer cell lines, have demonstrated enhancement of migratory capacity and stimulation of cytolytic activity of NK cells [21].
3.3. Exosomal Biomolecules Responsible for the Polarization of Macrophages
HCC derived exosomes, expressing miR-146-5p, lead to formation of M2-polarized tumor-associated macrophages, which inhibited the expression of IFN-γ and TNF-α while upregulating inhibitory receptors of such as PD-1 and CTLA-4 in T cells resulting in T cell exhaustion. Transcription factor Sal-like protein-4 (SALL4) was essential for regulating miR-146a-5p in HCC exosomes. Blocking the SALL4/miR-146a-5p interaction delayed HCC progression in mice model [51]. Polarization of M0 macrophage to M2 macrophage via exosomes released from tumor cells has also been observed in lung cancer [75].
3.4. Exosomal Biomolecules Modulating B-Cell Function
Studies have shown that exosomes play a role in altering B cell function [76]. Exosomes derived from HCC, exhibiting high mobility group box-1 (HMGB1), have been shown to increase proliferation of TIM-1+ regulatory B cells, which expressed IL-10 and suppressed CD8+ T cell proliferation leading to immune tolerance. Exosomal HMGB1 promoted TIM-1+ Breg cell proliferation via Toll-like receptor (TLR) 2/4 and mitogen-activated protein kinase (MAPK) signaling pathways [60]. In the tumor microenvironment, Bregs target tumor-infiltrating immune cells and interrupt antitumor immunity [77].
3.5. Exosomal Biomolecules Modulating Dendritic Cell (DC) Function
Tumor-derived exosomes, expressing IL-6, have been shown to suppress differentiation of bone marrow precursors into immature dendritic cells [61]. Ding G et al. reported that pancreatic tumors shed exosomes expressing miRNA-212-3p, which inhibit RFXAP expression and downregulate MHC-II expression on dendritic cells leading to immune tolerance [62]. In another study on Epstein-Barr virus (EBV)-associated gastric carcinoma (EBVaGC), it has been observed that EBV infected epithelial cells shed exosomes that inhibit dendritic cell maturation [78]. On the other hand, exosomes derived from squamous cell carcinoma, treated by 5-aminolevulinic acid photodynamic therapy, are able to stimulate DC maturation and promote antitumor immune response [79].
Pancreatic cancer cell derived exosomes bearing miR-203 have demonstrated diminished expression of toll-like receptor (TLR) and cytokines such as tumor necrosis factor-α (TNF-α) and interleukin-12 (IL-12) in DC [80]. It has been observed that regulatory T cells modulate DC function via exosomal miR-150-5p and miR-142-3p, which lead to formation of a tolerogenic phenotype with increased IL-10 and decreased IL-6 following LPS stimulation [81]. T cells release genomic and mitochondrial DNA within extracellular vesicles that prime DC to induce antiviral responses via the cGAS/STING cytosolic DNA-sensing pathway and expression of IRF3-dependent interferon regulated genes [82]. Furthermore, cancer cells treated with topotecan have shown to release DNA containing exosomes that activate DCs via STING signaling leading to antitumor immunity [83]. Similarly, irradiated breast cancer cells have been observed to transfer dsDNA to DCs and stimulate DC upregulation of costimulatory molecules and STING-dependent activation of IFN-I [84].
3.6. Exosomal Biomolecules Modulating Myeloid-Derived Suppressor Cell (MDSC) Function
Myeloid-derived suppressor cells inhibit T cell response in the tumor microenvironment [85]. It has been reported that gastric cancer cells release exosomes bearing miR-107 that cause activation and expansion of MDSCs [63]. In another study, glioma cells were shown to deliver hypoxia-inducible miR-10a and miR-21 via exosomes which caused activation and differentiation of MDSC leading to immunosuppressive effects [64].
4. Tumor Cell-Derived Exosomes in Immunotherapy
Tumor cells have emerged as a potential source for emitting exosomes [86]. Furthermore, it was found that tumor cells derive more exosomes than normal cells in the human body [87]. Tumor-derived exosomes (TEXs) act as a double-edged sword as they can play important role in cancer progression and metastasis by decreasing cytotoxicity and maintaining immunosuppressive tumor microenvironment as well as can improvise anti-tumor immune response by carrying tumor antigens and several heat shock proteins (HSPs) such as HSP70, HSP90 [88].
TEXs can be utilized as tumor vaccines based on their immunological properties. Various studies have revealed TEX as a potential source for cancer vaccines [89]. TEXs express various parental antigens similar to their parent tumor cells. As they influence both surrounding and distant organs, they can be used as an antigenic source for cancer vaccines [90]. Innate immune cells such as dendritic cells, macrophages when exposed to TEXs can be altered into pro-inflammatory cells to enhance the production of pro-inflammatory cytokines such as IL-6, IL-12, Interferon-γ as well as decrease anti-inflammatory cytokines such as IL-10 [90]. Since both neoantigen and adjuvant properties must be included in cancer vaccines to stimulate an immune response, TEXs having both these features are considered a potent source for formulating appropriate cancer vaccines. Therefore, clinical trials tried to use TEXs from malignant ascites to cure malignant pleural effusion [NCT02657460, NCT01854866] [90].
Tumor-associated antigens expressed on the surface of TEXs are similar to the parent tumor cell or via recipient APCs stimulating anti-tumor immune response [91]. Other immunostimulatory molecules such as CD80, OX40, OX40L, CD70, MHC and various tumor-associated antigens are expressed on the TEXs which also strengthen anti-tumor response [92]. DCs can take up and process tumor-associated antigens expressed on TEXs to trigger stronger immune responses [90].
TEXs contain HSP-70, HSP-90, MHC-1 molecules which act as trigger for enhancing immune response [88]. HSP70 derived from TEXs activates natural killer cells that lead to cancer cell lysis via granzyme B [93]. The anti-tumor response of TEXs is more effective than irradiated tumor cells, apoptotic bodies or lysate of cancer cells. Rao et al. demonstrated that hepatocellular carcinoma cell-derived exosomes elicit a more powerful DC-mediated immune response than lysate both in vitro and in vivo [94]. They have also induced better cancer vaccination effect than tumor cell lysates [95].
Zitvogel et al. in 2001 first demonstrated that TEXs induce cancer-specific T-cell immunity [96]. Moreover, exosomal EGFRvIII and transforming growth factor-β (TGF-β) together with other tumor antigens trigger immune responses mediated mainly by T and B cells [93]. For non-small cell lung cancer, TEXs derived exosomal proteins, specifically, TIM-3 and Galectin-9 might be useful biomarkers [97]. CD8 cytotoxic T cells undergo dysfunction and exhaustion during cancer progression due to immunosuppressive mechanisms. PD-L1 and CTL-associated antigen 4 (CTLA-4) have emerged as checkpoint receptors that are aimed at removing exhaustion of CD8 T cells [98]. Therefore, TEXs PD-l has emerged as a new therapeutic target to fight the resistance against current antibiotic approaches [99]. PD-L1 blockade to c-Met overexpressing cancer cells is an effective method to treat gastric cancer and other malignancies [100].
5. Immune Cell-Derived Exosomes in Immunotherapy
The immune system has an innate and an adaptive constituent, each having a distinct role and function. There are a variety of cells that take part in the innate and adaptive immunity. Recent studies have demonstrated exosomes are involved in the intercellular communication in the immune system as well as playing a role in immune modulation [101,102]. Exosomes released from immune cells can contain molecules that are essential for immune response initiation, antigen presentation along with intercellular exchange of membrane or cytosolic components without even the cell being in proximity to each other [103]. Moreover, based on the proteins packed in the exosomes, their target receptors and type of immune cell activation can vary (Figure 2). Recent studies have demonstrated antitumor effect of DC-derived tumor cells on top of their role in immunity [104,105].
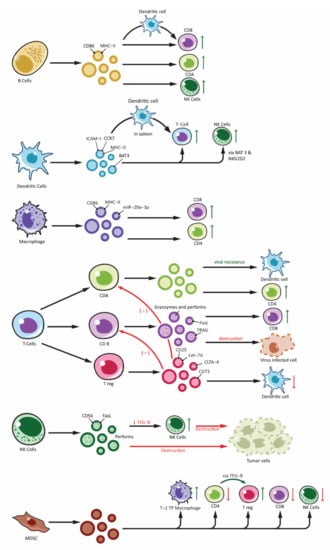
Figure 2.
Effects of immune cell-derived exosomes on cellular components of immune system. Showing exosomes produced from various immune cells: B-lymphocytes (B cells), T-lymphocytes (T Cells along with subdivisions of CD4+, CD8+ and Treg), Natural Killer cells (NK-cells), Dendritic cells and other antigen presenting cells (APCs) and Myeloid Derived Suppressor Cells (MDSC). Immune cell derived exosomes can affect various other cells participating in the immune responses. Exosomes can stimulate immune response by: peptide-MHC complex presentation to T cells, antigen transfer to other dendritic cells, activation of T cells, B cells and NK cells. Similarly, inhibitory function is derived from exosomes from Treg and NK cells and this include overall inhibition of the adaptive arm of immune response as well as destruction of tumor cells by NK cell. Exosomes from MDSC also plays an important role in immune inhibition mainly by activating Treg and Type-2 Tumor promoting phenotype macrophages (T-2 TP macrophage).
5.1. Dendritic Cells-Derived Exosomes
DCs are called the “master regulators” of the immune response as they commence all antigen-specific immune responses. They can efficiently process and present antigen, initiating the activation and proliferation of the T cell population [106]. The antigen is presented to T cell causing their subsequent activation via direct association between MHC II molecule on DC and TCR on T Cell. Another costimulatory signal is provided during this interaction (B7/CD28, LFA-1/ICAM-1 and ICAM2, CD2/LFA-3) [107]. This is called an immune synapse. Research has shown that exosomes released from activated DC contain MHCII, ICAM-1 and co-stimulatory molecules which can subsequently activate T cells albeit the process being less efficient [108,109]. However, exosomes from mature DCs may transfer MHC-peptide complex to other APCs which amplifies the T cell activation [110]. Moreover, it has been seen that exosome packed with miRNA from peripheral DC home to lymphoid tissue (e.g., spleen) which can then be taken up by immature cells residing there and activating them, thus augmenting the immune response. Research suggests that this migration of exosomes is due to presence of CCR7 molecules on their membrane [111]. Some studies have also shown that exosomes from dendritic cells can also activate NK cells [112]. This process is mediated via interaction of IL-15α and of NKG2DL on the exosome surface to their corresponding receptors on NK cells [113]. Elashiry at el. demonstrated that DC-derived exosomes loaded with molecular cargo (TGFβ1 and IL10) to modulate Th17/Treg balance is an effective immunotherapeutic approach to regulate degenerative bone disease in vivo [114]. DC-derived exosomes harbor unique proteins that protect them from clearance and attack by the complement system and help in binding to tissue integrins.
Similarly, macrophage, which is another APC, also produce exosomes with molecules like MHC-I, MHC-II, and costimulatory molecules like CD86 which can cause activate both CD4 and CD8+ T cells [115]. These findings solidify the importance of APC-derived exosomes in direct and indirect activation of the immune system.
5.2. B Cells-Derived Exosomes
B lymphocytes are an integral part of the adaptive component of immune system, responsible for antibody formation, antigen presentation as well as secreting an array of cytokines [116]. Similar to other APCs, B lymphocytes also release exosomes that contain abundant MHCII and CD86. Exosomes released from B lymphocytes play a vital role in the activation and downstream maintenance of the primed CD4+ T cell population. Studies have shown that activated T cell stimulates release of exosomes from B lymphocytes which in turn stimulates T cell proliferation and differentiation [105,117]. This process suggests their implication in propagating physiologic and pathologic immune response [118]. Interestingly, exosomes from B cell may also have MHCII and FasL which can bring about apoptosis of CD4+ T cells by association of Fas and FasL [119]. In a study, it was demonstrated that splenic langerin+, CD8α+ DCs can even activate CD8+ cytotoxic T cells by cross-presenting B cell-derived exosomal Antigen. This process was independent of host BCR expression and circulating Ab in contrast to exosomes derived from DC which required BCR presence [120]. However, this activation of cytotoxic CD8+ T cell by B lymphocyte-derived exosome still required CD4+, CD8+ and NK cells to be present [121]. B cell-derived exosome has also been found to contain integrins and ICAM -1 which facilitates its movement into tissue and thus delivers cargo loaded with necessary “ammunition” to proinflammatory cells at the site of inflammation [122]. Another fascinating discovery is the fixation of C3 fragments on EVs released from B cells. This causes enhanced antigen presentation and thus promotes better T cell response even when the level of antigen load is suboptimal [123].
5.3. T Cells-Derived Exosomes
T cell links the innate and adaptive components of the immune system [124]. T cells are mainly divided into 3 subsets, Helper T cell (CD4+), Cytotoxic T cell (CD8+) and regulatory T cells (Treg) based on their CD molecules on their surface and function. Exosomes derived from T cells also play a vital role in immune modulation. These exosomes typically have CD63 and CD81 as their transmembrane protein amongst other molecules, may also contain miRNA and even DNA [125]. They also contain FasL and APO ligand which can induce apoptosis in target cells [126].
It has been noted that genomic and mitochondrial DNA can be transferred from the T cell to DCs when they form immune synapses, causing the induction of antiviral response and making them resistant to viral infection [82]. Exosomes derived from CD4+ cells can cause reactivation of resting CD4+ cells [127].
CD8+ lymphocytes are cytotoxic T cells that destroy infected cells. They recognize foreign peptides on MHC class I molecules on the surface of target cells and eradicate them by either releasing granzymes and perforins or inducing apoptosis via FasL or TNF-related apoptosis-inducing ligand (TRAIL) expression [128]. Exosomes released from CD8+ lymphocytes have also been found to contain granzymes and perforins. It has been seen that exosomes are released from cytotoxic T lymphocytes (CTL) under the influence of IL-12 and the exosomes in turn can activate inactive CTL [129].
Treg cells which are another type of CD4+ cells can suppress the immune system. They exert their suppressive function on almost all cell types of immune system. T regs secrete more exosomes than any other type of T cell and they contain CD25, CD73 and CTLA4. Of these, CD73 seems to play a vital role in the suppression of immunity by producing adenosine while it is still unclear whether CD25 and CTLA4 play any part in immune suppression [130]. Exosomes from Treg cells also contain miRNA and when these exosomes were taken up by Th1 cells and DCs it caused inhibition of cytokine release of both these cells [81,131].
5.4. MDSC-Derived Exosomes
MDSC is a diverse group of immune cells that are derived from the myeloid cell lineage. These cells are immature neutrophils and monocytes, formed during pathologic activation and mainly suppresses the immune system by releasing a medley of cytokines [132,133]. Exosomes released from these cells have been implicated in immunosuppression as well as tumor growth and metastasis. Proteins present in the exosomes including S100A8 and S100A9 cause migration and accumulation in the tumor microenvironment, exerting a local immunosuppressive effect [134]. Another protein called High Mobility Group Box Protein 1 has been shown to promote the generation of MDSC from myeloid progenitor cells as well as mediating the secretion of IL-10 from MDSC which downregulates L-selectins and thus reduce T-cell chemotaxis [135]. Rashid et al. demonstrated that MDSC-derived exosomes inherit pro-tumorigenic factors and functionally resemble parental cells in immunosuppression, tumor growth, angiogenesis, invasion, and metastasis [136]. Furthermore, the study suggests that MDSC-derived exosomes are capable of increased reactive oxygen species (ROS) generation and inciting the Fas/FasL pathway, which precipitates activation-induced cell death (AICD) of CD8+ T-cells. Exosomes from MDSC has also been shown to carry TGF-β1 which participates in the inhibition of NK-cells [137]. Further research also showed that exosomes from MDSC, in vitro, promoted Treg differentiation and proliferation from CD4+ T cells in the presence of TGF-β and inhibited CD4+ cell proliferation [138]. MDSC exosomes has also been implicated in the suppression of the innate arm of immune system by their ability to polarize macrophages to the type 2 tumor promoting phenotype [139]. While implicated in tumorogenesis primarily, further research may shed light on whether MDSC and their exosome cause immune modulation in non-tumorous setup.
5.5. Mast Cells-Derived Exosomes
Mast cells are bone marrow-derived immune cells that reside in the mucosa and epithelial cells of the body and has a wide range of function- they contain an array of granules that are responsible for vasodilation, angiogenesis, killing certain bacteria and eliminating parasites from our body in addition to their well-documented role in type 1 hypersensitivity [140]. Mast cells also secrete a myriad of cytokines which can also influence both B and T cell function [141,142]. Studies suggest does not necessarily activate B and T lymphocytes through direct contact but rather through secretion of exosomes [143]. Mast cell-derived exosomes carry OX40L and it has been suggested that the interaction of OX40L and OX40 on CD4+ cells cause differentiation of Th2 cells [144]. Exosomes derived from mast cells can also influence activation of DC by inducing release of IL-6, IL-12 and IFN-γ release from CD4+ T cells [145]. There are conflicting studies on the role of exosomes on perpetuating IgE-dependent immune response (and subsequently allergic reactions), with some suggesting it alleviates it [146] while others suggesting it potentiates it [147]. Hence more study must be done to explore this topic.
5.6. Neutrophil-Derived Exosomes
Neutrophils are polymorphonuclear cells that act as “first responders” in an inflammatory reaction. Their main function is to destroy microorganisms, mainly through phagocytosis [148]. Neutrophils are also capable of secreting exosomes like so many of their counterpart cells in the immune system [149]. Exosomes derived from neutrophils carry a range of antimicrobial agents such as lactoferrin, OLFM4, LCN2, S100A8 and MMP9 [150]. There are many stimuli to the production and release of exosomes from neutrophils. Some of these are bacterial antigens (e.g., fMLP and LPS), proinflammatory cytokines (e.g., TNFα, IFN γ, GM-CSF, etc.), presence of specific pathogens (e.g., M. tuberculosis, P. aeruginosa, etc.) and even some drugs like ionomycin. These exosomes can have a multitude of abilities depending on the stimulus inducing their release and the effector cells. For example, exosomes released due to LPS/fMLP stimulus can cause an anti-inflammatory effect on certain cells of the immune system like monocytes and macrophages by causing TGF-β1 release which in turn suppresses their function [151]. The very same stimulus (LPS/fMLP) is proinflammatory, containing LTB4 which acts as a chemoattractant, causing chemotaxis of neighboring neutrophils towards the inciting inflammatory area [152]. Interestingly, it has been demonstrated that M. tuberculosis can induce secretion of exosomes from neutrophils which interfere with macrophages’ activity against the bacteria which suggests they play a vital role in the pathogenesis of M. tuberculosis [153]. Research has also shown that apoptotic neutrophils can release exosomes which can suppress resting CD4+ Helper T cells by causing downregulation of IL-2 and IL-2R. This downregulates unnecessary activation of resting helper T cells and thus seems to play a supporting role in maintaining immune tolerance [154]. Thus, even though neutrophils normally play an important role in innate immunity, it intercellular communication via exosomes, they also regulate the adaptive component of the immune system.
5.7. NK Cells-Derived Exosomes
NK cell is a specialized lymphocyte that mainly takes part in innate immunity, their main function is to eradicate tumor and viral infected cells [155]. NK cells also secrete exosomes. The exosomes derived from NK cell has NK cell markers (e.g., CD 56) as well as proteins to destroy cells (FasL and perforins) and take part in immune homeostasis [156]. NK cell exosomes also are found to carry miRNA-186, which acts as a tumor suppressor miRNA and also prevents inhibition of NK cells by TGF-β [157]. Exosomes from NK cells can modulate other immune cells via expression of various molecules on their surface. One study has established presence of TNF receptors and ligands along with interferon-induced transmembrane proteins on exosome surface and it was hypothesized that via use of these molecules, these exosomes can influence T and B cells [158].
6. Modulation and Evasion of the Complement System by Exosomes
Often regarded as the first line of defense against pathogens, the complement system is an important component of innate immunity [159]. Not only does it play an important role in opsonization and elimination of pathogens, activation of complement cascade results in the release of chemokines and cytokines, interact with coagulation cascade and regulate inflammation process and repair [160]. The complement system consists of more than 30 proteins and activated in one of three pathways, classic pathway, alternate pathway and lectin pathway [161]. Each of the three pathway results in the generation of C3 convertase which subsequently triggers formation of membrane attack complex (MAC) and cell lysis [162]. During local and systemic inflammation, there are more circulating exosomes, as well as elevated complement activation products, this suggests a link between exosome and complement system [163]. Exosomes has been shown to play a role in complement activation via different molecules on their surface (Figure 3). Exosomes released from APCs are likely to be loaded with antigenic protein and hence have the potential to bind to immunoglobulins and thereafter cause activation of classic pathway [164].
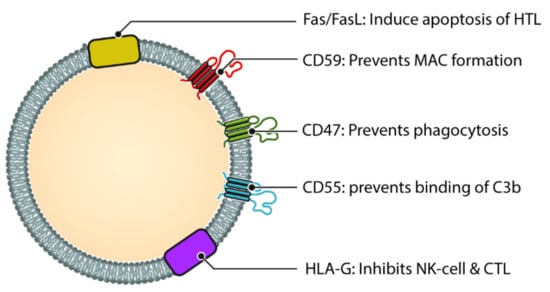
Figure 3.
Different molecules that help exosomes escaping complement and immune system. Helper T cell (HTL), Natural Killer cell (NK Cell) and Cytotoxic T-lymphocyte (CTL). CD47, CD55 and CD59 help prevent the exosome from being destroyed by the complement cascade, either directly (via MAC) or indirectly. Molecules such as HLA-G and Fas/FasL protect the exosome from immune mediated destruction via cell mediated immunity (Cytotoxic T cell and NK cell).
Also, it appears that APC derived exosomes are able to escape complement-mediated lysis by expressing CD55 which regulates C3 and C5 convertases and CD59 which inhibits the MAC formation on the exosome surface [164,165]. Moreover, exosomes from PMN cells also express complement regulators like Complement Receptor 1 (CR1) which helps them bind to opsonized bacteria efficiently. Furthermore, these exosomes can scavenge C1q and subsequently C3 which allows them to bind to erythrocytes in circulation [166]. This resembles a circulating immune complex and are transported to the liver and spleen and cleared along with the opsonized bacteria. Studies have also suggested that erythrocytes at the end of their life span secrete exosomes rich in CR1 and CD59 and thus makes the aged erythrocyte vulnerable to opsonization and complement attack, helping in clearance of these erythrocytes by phagocytosis [167].
Studies also suggest that exosomes interacting with complements may further amplify complement activation. The synovial fluid of patients with rheumatoid arthritis showed significant increased levels of exosomes derived from leucocytes with complements bound to their surface, including C1q, C3 and C4 [168]. It is also intriguing to note that in vitro studies have shown exosomes from leucocytes can bind C1q and cause activation of classic pathway (CP), causing deposition of C3 and C4 [169,170]. Research by Gasser, O. and J.A. Schifferli further reaffirmed that once C1q was deposited on exosome, there was C3 and C4 deposition on the exosome surface [166].
Exosomes also cause complement system activation through CRP binding on its surface. Liver synthesizes C-reactive protein (CRP) in pentameric form (pCRP) [171] which localizes to site of inflammation, undergoes a change in conformation and becomes pCRP*. The pCRP* has the ability to dissociate to monomeric CRP (mCRP). Both pCRP* and mCRP has been implicated in the activation of complement pathway [172,173]. Studies have demonstrated pCRP molecules bound to exosome surfaces producedpCRP*/mCRP, which can activate the complement system. Moreover, exosomes with CRP bound to their surface also acquired the ability to attach to C1q which, as discussed previously, activated the complement pathway [173].
Moreover, new research suggests that exosomes may play a role in complement cascade suppression. A study by Loh, J.T., et al. showed that exosomes derived from MSC can inhibit the terminal steps of the complement cascade. This leads to suppression of neutrophils activation via complement pathway. The exact mechanism by which this is achieved is unknown but it has been seen that CD 59 is implicated in this suppression of complement system [174]. These exosomes have also inhibited the release of Neutrophil Extracellular Trap (NET) and IL-17 from neutrophils.
Interestingly, exosomes are secreted with unique molecules on their surface that helps them evade the immune and the complement system and prevent their destruction. Some of these molecules are CD47, CD55, CD59 and CK2. Both CD55 and CD59 has been discovered in exosomes released from APCs in high quantity which protects exosomes from complement mediated destruction [164]. CD55 prevents binding of C3b on the surface of exosome and thus prevents its phagocytosis while CD59 prevents MAC formation [164]. Additionally, evidence suggests that the anti-phagocytic protein CD47 is found on the surface of exosomes made by T lymphocytes, among others. CD47 works by interacting with Sirp-α to prevent macrophages from ingesting exosomes and preventing them from being phagocytosed [175]. CK2 is another protein found in membrane vesicles that phosphorylates complement C9 and protects against complement-mediated lysis [176].
Intriguingly, exosomes also express several molecules on their surface which confers them with immune tolerogenicity. One of them is Human leucocyte antigen (HLA)-G [177]. Immunomodulatory properties of these HLA-G molecule include inhibition of natural killer (NK) cytolysis and CTL activation [178,179]. Similarly, exosomes released from professional antigen presenting cells like B lymphocytes and Dendritic cells also contain MHC class I and II protein [180]. Also, what is noteworthy is that activated B cells release exosomes containing FasL and MHC II. Evidence suggests that these molecules can induce self-tolerance by inducing apoptosis of activated helper T cell [181]. This demonstrates the complex interplay between the exosomes derived from immune cells and the immune and complement system.
7. Engineered Exosomes in Immunotherapy
Exosomes have opened a new direction for treating life-threatening diseases by immunotherapy which is further strengthened by various engineering mechanisms to trigger a stronger immune response. Not only biomimetic but also artificial exosomes can be used for immunotherapy. Biomimetic exosomes can deliver monoclonal antibody DEC205 to DCs [182]. Artificial exosomes are utilized for targeting DCs and activating T cells by modifying them with MHC-1 peptides and liposomes; liposome-containing exosomes presented to be stable and appropriate as well [183].
Different engineering methods have been identified to modify the function of exosomes. Some of them are described below.
7.1. Genetic Engineering
Several mechanisms of genetic engineering have been applied to modify the exosomes to elicit better outcomes in immunotherapy. They are proven to be a suitable delivery vehicle for antibodies as well as chemotherapeutic drugs based on their promising modifiability, compatibility and cyclical half-life [184].
A new method called exosome reprogramming is getting a lot of attention which increases the efficacy of natural exosomes. To genetically modify these exosomes a novel platform called synthetic multivalent antibodies retargeted exosome (SMART-Exo) is established [185]. Furthermore, to improve aimed drug delivery in the presence of an external magnetic field, another system called exosome-based superparamagnetic nanoparticle cluster (SMNC-EXO) has been developed [186]. Different exosome engineered genetic methods are giving a promising outlook in the immunotherapy field for the near future (Table 2).
Table 2.
Genetic engineering methods of exosomes in immunotherapy.
7.2. miRNA Modification
Exosomes treated with DHA (docosahexaenoic acid) have been found to express increased levels of let-7a, miR-23b, miR-27a/b, miR-21, let-7, and miR-320b and these are tumor suppressor miRNA. When these treated exosomes are incubated with unexposed epithelial cells, the epithelial cells also express higher levels of tumor-suppressing miRNA. Consequently, exosome-mediated signaling between tumor cells should be targeted for immunotherapy [198]. A study has revealed that upregulation of Let-7i and miR-142 in TEXs result in stronger stimulatory effects on either DC maturation or cytotoxic T cell induction and cytokine release [199]. Another study result indicated that miRNA-depleted TEXs proteins could be considered a potential agonist to trigger DCs activation [200].
As biomarkers, immune cell-derived exosomes containing miRNA play an important role. A study revealed that 150 unique exosomal miRNAs were identified in cancer patients. Among these miRNAs, hsa-miR-320d, hsa-miR-320c, and hsa-miR-320b might be promising biomarkers for predicting the efficacy of PD-1/PD-L1 immunotherapy in lung cancer [201]. Likewise, in epithelial ovarian cancer, plasma-derived exosomal miR-4732-5p is found at a higher level and could be used as a prognostic factor [202].
7.3. Conjugation
The process of conjugation is used to bind different activator molecules with exosomes to stimulate and activate the immune cells. TLR ligands such as CpG-DNA, Poly:IC and IL15Rα conjugated to TEXs can enhance immune response. NKG2D ligand binding can also activate natural killer cells [203]. Chondroitin sulfate (CS) is increasingly expressed in cancer cells. VAR2CSA is a malarial protein that binds to VAR2CSA-ligand or CS which is expressed in placental tissue. Therefore, CS in malignant cells could be potentially targeted by recombinant VAR2CSA (rVAR2). Targeting the common CS chain which is present on various cancer cells could offer a novel treatment pathway [204]. A new strategy named exosomes for protein loading via optically reversible protein-protein interactions (EXPLORs) where blue light controls the integration of protein-protein interaction during endogenous exosome biogenesis. This method is used for intracellular delivery of target proteins [205]. Moreover, a study has found that IRF-1 which has tumor apoptosis properties can be conjugated with TEXs and resulting cells reveal higher levels of IL-15R and MHC-I triggering stronger activation and infiltration of both CD4+ and CD8+ T cells [206].
7.4. HSP70/90 Overexpression (Heat Treatment)
HSP expressed on exosomes are recognized as a potential target for immunotherapy. Different HSP have been identified on the surface of TEXs such as 71-kDa HSP, the 70-kDa HSP4, and HSP90 alpha and beta [134]. HSP70 expressing exosomes derived from pancreatic carcinoma cells enhance lytic activity of natural killer cells against HSP70 surface positive tumors [21]. If exogenous lipid properties are modified during interaction between cells and engineered exosomes, it will improve targeted drug delivery or antigen presentation to immune cells in lymph node and delivered CRISPR/Cas9 system in Mesenchymal Stem cells (MSCs) to prevent in vivo gene editing [207]. Glycosylation also protects exosomes from proteolytic degradation and directs them to specific targets. Therefore, this method improves targeted peptide expression and strengthens the effect of therapeutic drugs [208]. Tumor cell-derived chaperon-rich cell lysates, containing Hsp70, Hsp90, calreticulin, and glucose-regulated protein 94 are strong stimulators of DCs and improve immunotherapeutic activity of T cells against intracranial glioma [209].
8. Advantages of Using Exosomes for Immunotherapy
The application of exosomes in immunotherapy offers noteworthy prospects for novel & effective treatment in an extensive range of pathological conditions. There are a plethora of positive findings regarding this in numerous clinical trials.
Structurally, exosome possesses an intermediate level of organization that distinguishes them from other molecular or organ levels of therapeutics. They contain heterogeneous functional molecules as well as avoid the complexity of organ-level [90]. Also, the gel-like cytoplasm and deformable cytoskeleton provide structural integrity and rupture resistance that makes exosome an exclusive diagnostic, prognostic, or therapeutic tool [210]. Natural cargos like lipids, proteins, nucleic acids are used for these purposes. These molecules can also be modified according to specific use. The modification strategies include adapting surface molecules to enable specific targeting, passive loading of hydrophobic drugs on membrane & introducing hydrophilic cargos into the core [210].
Being comprised with lipid-bilayer, exosomes effectively transfer their cargo without any interference by extra-cellular enzymes & maintain their bioavailability. Most of the drugs have poor oral bioavailability as they have to encounter the first-pass metabolism by the liver. So, these drugs are introduced intravenously to retain their bioavailability [211]. But concerning the convenience of oral administration over IV administration, cow milk exosomes have demonstrated the potential for boosting oral drug bioavailability in several clinical trials [212,213,214].
Exosomes play a key role in the detection of various diseases as they contain specific biomarkers (proteins, lipids, RNAs) in their lumen. They possess ’clathrin-coat’ on their membrane which protects their content from degradation by enzymes such as RNases. This coat makes them more stable, resistant & enables them to transport various components efficiently (drug delivery) [36,215,216,217]. So, they have been significantly used as a diagnostic tool to detect lung cancer, Alzheimer’s disease, colon cancer & many other diseases [218,219,220]. Also, they can be used as non-invasive biomarkers as they are found in several body fluids like plasma, saliva, urine, gingival crevicular fluid, etc. [221].
As an endogenous nanoparticle, the exosome may act as a potential carrier in gene therapy. The purpose of gene therapy is gene silencing with specific siRNA but the delivery of this was always a concern due to the lack of efficient carrier. The viral and non-viral carriers used in this aspect often result in systemic toxicity [222]. In this respect, milk exosome loaded with siRNA was introduced and a significant anti-proliferative effect against lung cancer cells was observed. This result indicated exosome as a safe & viable natural carrier in gene therapy [223].
Additionally, exosomes are utilized to induce phagocytosis as well as to block phagocytosis for therapeutic convenience. By expressing phosphatidylserine on their surface, they provide a vigorous “Eat-Me” signal to macrophages & thus they can trigger the phagocytosis process. Cancer cells also express phosphatidylserine on their surface but it isn’t sufficient to initiate phagocytosis [224]. On the other hand, immunocyte-derived exosomes express CD47 receptor on their surface that interacts with Signal Regulating Protein α (SIRPα) which triggers the “Don’t-Eat-Me” signal & blocks their consumption by phagocytes [225,226].
Vaccine immunotherapy using exosomes has emerged as a promising approach to the treatment of many cancer patients. Patients of Non-Small Cell Lung Cancer (NSCLC) experienced leukapheresis to produce dendritic cell (DC)-derived exosomes (DEX) which are loaded with MAGE-A3, -A4, -A10, and MAGE-3DP04 peptides. This phase-I study suggested that DEX therapy was admissible in patients with advanced NSCLC [227]. In another study, autologous DEX was applied as a vaccine to treat metastatic melanoma patients [228].
Similarly, the cell-surface modification strategy can be adopted to achieve targeted cellular action. For example, engineered exosomes expressing transmembrane domain of platelet-derived growth factor fused to the GE11 peptide can successfully deliver miRNA to epidermal growth factor receptor-expressing breast cancer cells [229]. Thus, the expected cellular action of nucleic acid drugs can be secured through exosome-based drug delivery.
Being a nano-vesicle, exosomes are capable of penetrating deeper tissue or overcoming tissue barriers. For instance, the efficiency of exosomes to cross the blood-brain barrier has been demonstrated in several studies. A recent study tested 10 exosome populations derived from mouse, human, cancerous, or non-cancerous cells to cross BBB. All exosome types efficiently crossed BBB and volume of distribution inside the brain was satisfactory. The uptake pattern was homogenous in most cases, but some exosomes showed precedence of uptake by the olfactory bulb [230]. Another study demonstrated brain endothelial cell (ECs)-derived exosome as a potential carrier of anticancer drugs to treat brain cancer as they efficiently delivered siRNA into brain tissue crossing BBB in Zebrafish having brain tumor [231]. Although the exact mechanisms how exosomes cross BBB yet to be fully elucidated, it is postulated that exosomes from circulation are transferred via brain vascular ECs in three main mechanisms as follows; physical contact (fusion or ligand-receptor interaction, etc.), paracytosis, and transcytosis [232,233]. In the fusion method, exosomes attach and fuse with barrier type ECs to release cargo onto the cytosol [234]. Exosome membrane proteins and specific receptors on the ECs surface initiate the existence of gap junction-like communications and several research articles have reported involvement of specific interaction molecules [235], such as lymphocyte function-associated antigen-1 (LFA-1) on EVs interacting with intercellular adhesion molecule (ICAM)-1/C-type lectin receptor on endothelial cells [236], transferrin-transferrin receptor interactions [237], CD46 as major EV uptake receptor [238] or 6-mannose-receptor involvement for one specific EV type [230]. In the transcytosis route, following cellular entry, exosomes are directed for degradation or to endosomes for transmitting to the abluminal surface of ECs [232]. Brain endothelial cell-derived exosomes can be potentially used as the carrier of anticancer drugs to treat brain cancer [231]. Additionally, the nanosize of the exosome ensures easy passing through tumor vasculature. The physiological upper-limit of pore-size within Blood Tumor Barrier is 7–100 nm in case of malignant brain tumor, whereas within the BTB of peripheral tumor, upper limit of pore-size ranges between 200–1200 nm [239]. Another study reported that tumor of subcutaneous micro-environment has a tumor-dependent functional pore cut-off size ranges between 200 nm to 1.2 µm [240]. Having size range of 30–100 nm, exosomes can cross all these biological barriers without effort [241].
Synthetic nanoparticles, liposomes, are small, unilamellar-vesicles, composed of phospholipids which are exogenously synthesized & are applied as drug delivery nano-carrier. Being a foreign particle, liposome has encountered multiple defensive mechanisms inside the body [242]. Furthermore, in contrast to the exosomes, they can produce toxicity in overdose. All these drawbacks diminish liposomal function & their potency [As summarized in Table 3 [5,243,244,245,246]].
Table 3.
Comparison between liposomes and exosomes as drug delivery vehicles.
Though there is impressive progress in developing the isolation techniques of exosomes, it is also a matter of concern that rapid and efficient isolation is still challenging and underway [28]. Consequently, a gold standard consensus of isolation & purification is unsubstantiated [247]. Furthermore, the isolation techniques face more challenges as some other components of biological fluid possess the same size as exosomes [247,248]. Also, to maintain maximum functionality, proper storage is crucial. But, a recent study indicated significant morphological changes due to the storage of mouse bronchoalveolar lavage fluid exosome and also its content. Exosomes stored at +4 °C presented a 10% increase in diameter and reduction of zeta potential, while a 25% increase in diameter and a significantly reduced zeta potential were seen at −80 °C compared to fresh exosomes [249]. But there are also some contradictions in this aspect as some other studies showed the efficacy of exosomes even after storage. For instance, the exosomes were stable and biologically active even after 90 days of storage by using OptiPrep(TM) density gradient technique [250]. Furthermore, a recent study revealed that in comparison to freshly collected exosome, iExosome stored at −80 °C for a period of 45 days was unaffected in respect to their number, size, distribution & biological activity. But a reduction of stability and functionality was observed at room temperature or +4 °C for 2 or more days [251]. Other challenges include large scale production, proper purification, maintaining good manufacturing practice, limited possibility of drug loading etc. [252].
9. Clinical Trials of Exosomes as Immunotherapy
Numerous clinical trials have been conducted with exosomes based on broad research into the contribution of exosomes as immune-modulation, diagnostic biomarker, therapeutic target & drug delivery [253,254]. A summary of ongoing clinical trials of exosomes for immunotherapy is described in Table 4. MSC and DC are two well-received sources in this regard. DeX can trigger inflammation and induce specific immune responses while MEX can be applied in the treatment of inflammation [255]. In this pandemic, the treatment of COVID-19 may be achieved by using MEX secreted particles, incorporating specific miRNA or mRNA into exosomes, or using MEX as drug carriers [256]. There are several trials in this aspect. As mentioned, the anti-inflammatory effect of MEX may alleviate the hyperinflammatory condition in the body arising from cytokine storm. Intravenously administered or inhaled MEX may be a way to suppress the immune response and also activate the regeneration of inflamed type II alveolar epithelium in patients with pneumonia, acute respiratory distress syndrome & mild or severe cases of COVID-19 (Table 4). Engineered exosomes overexpressing CD 24 can control the homeostatic proliferation of T cells and alleviate the symptoms of SARS-CoV-2. In this trial, patients received exosomes derived from embryonic kidney T cells that can directly suppress cytokine storm. Also having a highly compatible vehicle delivery, exosome ensured a reduction of the required dose. In an ongoing trial, MEX nebulizer is being used as a treatment of lung infection caused by gram-negative bacilli that are already antibiotic-resistant. Visible improvement of the pathological changes caused by infection, reduction of pulmonary edema & mitigation of alveolar inflammation were seen. In the case of craniofacial neuralgia, exosomes were introduced via epineural injection guided with ultrasound. Having the ability to cross the blood-brain barrier, exosomes may give an anti-inflammatory effect by initiating some intercellular pathway [230]. MEX is luminally loaded with small interference RNA (siRNA) that can act against KRAS G12D & thus may cure metastatic pancreatic ductal adenocarcinoma. This study revealed that participants developed no toxicity after receiving MEX for 14 days. Alongside, plant-derived exosomes, like mammalian exosomes, play a pivotal role by acting as an anti-inflammatory agent [257]. Exploring the role of grape extract exosome to improve oral mucositis during chemoradiation therapy for the treatment of head & neck cancer is underway. Likewise, oral administration of exosomes liberated from ginger extract may elevate anti-inflammatory cytokines (IL-10 & IL-22) & bring down the proinflammatory cytokines (TNF-α, IL-6 & IL-1β). Hence, they may be highly suitable for the treatment of Inflammatory Bowel Disease & Colitis associated cancer [258]. MEX serves as an anti-inflammatory agent by expression of very low MHC antigens. Thus, they can be reliably used for the treatment of Type 1 Diabetes Mellitus. Besides the immunotherapeutic application, the role of exosomes as diagnostic or prognostic biomarkers has also been explored in many trials. For example, in sepsis-induced immunosuppression, observing circulating exosomes may define the immune status of patients. In type-1 Diabetes mellitus, β-cells of the pancreas liberate exosomes that contain β-cell-specific autoantigens. This may provoke the immune response at the initiation of Type-1 Diabetes Mellitus & thus they aid in the early detection of disease. Similarly, exosomes may help to identify Obstructive Sleep Apnea Syndrome by upregulating the PD1/PDL1 pathway. Exosomes liberated from B-cell Non-Hodgkin Lymphoma can favor lymphoma cells to escape from humoral immunotherapy. They express CD20 that could act as “Decoy Target” upon Rituximab exposure. Thus, exosomes significantly contribute to therapeutic resistance. Again, in Diffuse Large B-Cell Lymphoma patients, combined treatment of Rituximab & chemotherapy can elevate serum exosome miR-451a which acts as an index for evaluating the potency of combined treatment with Rituximab & chemotherapy in DLBCL patients [259]. The use of exosomes for vaccination purposes has also come into interest in some trials. Immunotherapy with mCTX followed by vaccination with tumor antigen-loaded DeX was performed in NSCLC patients. Phase 1 trial showed the safety and efficacy of DeX vaccine but T cell induction was undetected. Consequently, phase 2 trial was conducted and the study premise was DeX based treatment to improve Progression free survival rate at 4 months in advanced NSCLC patients.
Table 4.
Ongoing clinical trials of exosomes for immunotherapy.
10. Conclusions
Because of its specificity and fewer side effects compared to the other modality of therapy, immunotherapy emerged as one of the most explored areas in the new era of numerous diseases, specifically in cancer treatment. To develop targeted immunotherapy, it is indispensable to understand the mechanism of immune responses during the disease process and reciprocal interaction between the pathological tissue and cellular components of the immune system. In this regard, accumulating evidence suggests that exosomes play a crucial role in intercellular communication and depending on the parent cell type and the cargo, exosomes can have immunosuppressive or immunostimulatory effects. Due to their intrinsic properties and advantages, exosomes can be utilized as immunotherapeutics and as a carrier for other immunomodulatory agents. Although there are some limitations in isolation and clinical application, exosome-based immunotherapies have immense potential which can revolutionize the treatment of cancer and other pathologies. Hence, this is a compelling area of research that needs extensive exploration to comprehend the role of exosomes in disease pathology and immune modulation and to use these concepts for the development of new exosome-based immunotherapies.
Author Contributions
Conceptualization and supervision, M.H.R.; writing—original draft preparation, M.W.A.H., S.J., B.G., F.Y. and A.A.; writing—review and editing, M.H.R., M.W.A.H., S.J., F.Y., A.A. and S.N.S.; graphical figures, F.A. and M.W.A.H. All authors have read and agreed to the published version of the manuscript.
Funding
This review received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Toro, J.; Herschlik, L.; Waldner, C.; Mongini, C. Emerging roles of exosomes in normal and pathological conditions: New insights for diagnosis and therapeutic applications. Front. Immunol. 2015, 6, 203. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Rajasingh, S.; Drosos, N.; Zhou, Z.; Dawn, B.; Rajasingh, J. Exosomes: New molecular targets of diseases. Acta Pharmacol. Sin. 2018, 39, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Naseri, M.; Bozorgmehr, M.; Zöller, M.; Pirmardan, E.R.; Madjd, Z. Tumor-derived exosomes: The next generation of promising cell-free vaccines in cancer immunotherapy. OncoImmunology 2020, 9, 1779991. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Batista, I.A.; Melo, S.A. Exosomes and the Future of Immunotherapy in Pancreatic Cancer. Int. J. Mol. Sci. 2019, 20, 567. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Tkach, M.; Théry, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef]
- Kalluri, R. The biology and function of exosomes in cancer. J. Clin. Investig. 2016, 126, 1208–1215. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. Discovery of Double-Stranded Genomic DNA in Circulating Exosomes. Cold Spring Harb. Symp. Quant. Biol. 2016, 81, 275–280. [Google Scholar] [CrossRef]
- Robbins, P.D.; Morelli, A.E. Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol. 2014, 14, 195–208. [Google Scholar] [CrossRef]
- Nilsson, P.; Sekiguchi, M.; Akagi, T.; Izumi, S.; Komori, T.; Hui, K.; Sörgjerd, K.; Tanaka, M.; Saito, T.; Saido, T.C.; et al. Autophagy-related protein 7 deficiency in amyloid beta (Abeta) precursor protein transgenic mice decreases Abeta in the multivesicular bodies and induces Abeta accumulation in the Golgi. Am. J. Pathol. 2015, 185, 305–313. [Google Scholar] [CrossRef]
- De Gassart, A.; Geminard, C.; Hoekstra, D.; Vidal, M. Exosome secretion: The art of reutilizing nonrecycled proteins? Traffic 2004, 5, 896–903. [Google Scholar] [CrossRef]
- He, C.; Zheng, S.; Luo, Y.; Wang, B. Exosome Theranostics: Biology and Translational Medicine. Theranostics 2018, 8, 237–255. [Google Scholar] [CrossRef]
- Cordonnier, M.; Chanteloup, G.; Isambert, N.; Seigneuric, R.; Fumoleau, P.; Garrido, C.; Gobbo, J. Exosomes in cancer theranostic: Diamonds in the rough. Cell Adhes. Migr. 2017, 11, 151–163. [Google Scholar] [CrossRef]
- Williams, R.L.; Urbe, S. The emerging shape of the ESCRT machinery. Nat. Rev. Mol. Cell Biol. 2007, 8, 355–368. [Google Scholar] [CrossRef]
- Hurley, J.H. ESCRT complexes and the biogenesis of multivesicular bodies. Curr. Opin. Cell Biol. 2008, 20, 4–11. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Bu, H.; He, D.; He, X.; Wang, K. Exosomes: Isolation, Analysis, and Applications in Cancer Detection and Therapy. ChemBioChem 2019, 20, 451–461. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Schorey, J.S. Exosomes Released from Infected Macrophages Contain Mycobacterium avium Glycopeptidolipids and Are Proinflammatory. J. Biol. Chem. 2007, 282, 25779–25789. [Google Scholar] [CrossRef]
- Vega, V.L.; Rodríguez-Silva, M.; Frey, T.; Gehrmann, M.; Diaz, J.C.; Steinem, C.; Multhoff, G.; Arispe, N.; De Maio, A. Hsp70 Translocates into the Plasma Membrane after Stress and Is Released into the Extracellular Environment in a Membrane-Associated Form that Activates Macrophages. J. Immunol. 2008, 180, 4299–4307. [Google Scholar] [CrossRef]
- Gastpar, R.; Gehrmann, M.; Bausero, M.A.; Asea, A.; Gross, C.; Schroeder, J.A.; Multhoff, G. Heat Shock Protein 70 Surface-Positive Tumor Exosomes Stimulate Migratory and Cytolytic Activity of Natural Killer Cells. Cancer Res. 2005, 65, 5238–5247. [Google Scholar] [CrossRef] [PubMed]
- Skokos, D.; Botros, H.G.; Demeure, C.; Morin, J.; Peronet, R.; Birkenmeier, G.; Boudaly, S.; Mécheri, S. Mast Cell-Derived Exosomes Induce Phenotypic and Functional Maturation of Dendritic Cells and Elicit Specific Immune Responses In Vivo. J. Immunol. 2003, 170, 3037–3045. [Google Scholar] [CrossRef] [PubMed]
- Segura, E.; Amigorena, S.; Théry, C. Mature dendritic cells secrete exosomes with strong ability to induce antigen-specific effector immune responses. Blood Cells Mol. Dis. 2005, 35, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Budnik, V.; Ruiz-Cañada, C.; Wendler, F. Extracellular vesicles round off communication in the nervous system. Nat. Rev. Neurosci. 2016, 17, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, J.; Purkayastha, S.; Tang, Y.; Zhang, H.; Yin, Y.; Li, B.; Liu, G.; Cai, D. Hypothalamic programming of systemic ageing involving IKK-beta, NF-kappaB and GnRH. Nature 2013, 497, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Lässer, C. Identification and Analysis of Circulating Exosomal microRNA in Human Body Fluids. Methods in Pharmacology and Toxicology 2013, 1024, 109–128. [Google Scholar] [CrossRef]
- Rashid, M.H.; Borin, T.F.; Ara, R.; Angara, K.; Cai, J.; Achyut, B.R.; Liu, Y.; Arbab, A.S. Differential in vivo biodistribution of 131I-labeled exosomes from diverse cellular origins and its implication for theranostic application. Nanomed. Nanotechnol. Biol. Med. 2019, 21, 102072. [Google Scholar] [CrossRef]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in Exosome Isolation Techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef]
- Saleh, R.; Elkord, E. Exosomes: Biological Carriers and Promising Tools for Cancer Immunotherapy. Vaccines 2020, 8, 390. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, W.; Zhang, H.; Zhang, F.; Chen, L.; Ma, L.; Larcher, L.M.; Chen, S.; Liu, N.; Zhao, Q.; et al. Progress, opportunity, and perspective on exosome isolation—Efforts for efficient exosome-based theranostics. Theranostics 2020, 10, 3684–3707. [Google Scholar] [CrossRef]
- Lamparski, H.G.; Metha-Damani, A.; Yao, J.-Y.; Patel, S.; Hsu, D.-H.; Ruegg, C.; Le Pecq, J.-B. Production and characterization of clinical grade exosomes derived from dendritic cells. J. Immunol. Methods 2002, 270, 211–226. [Google Scholar] [CrossRef]
- Busatto, S.; Vilanilam, G.; Ticer, T.; Lin, W.-L.; Dickson, D.W.; Shapiro, S.; Bergese, P.; Wolfram, J. Tangential Flow Filtration for Highly Efficient Concentration of Extracellular Vesicles from Large Volumes of Fluid. Cells 2018, 7, 273. [Google Scholar] [CrossRef]
- Haraszti, R.A.; Miller, R.; Stoppato, M.; Sere, Y.Y.; Coles, A.; Didiot, M.-C.; Wollacott, R.; Sapp, E.; Dubuke, M.L.; Li, X.; et al. Exosomes Produced from 3D Cultures of MSCs by Tangential Flow Filtration Show Higher Yield and Improved Activity. Mol. Ther. 2018, 26, 2838–2847. [Google Scholar] [CrossRef]
- Reiner, A.T.; Witwer, K.W.; van Balkom, B.W.; de Beer, J.; Brodie, C.; Corteling, R.L.; Gabrielsson, S.; Gimona, M.; Ibrahim, A.G.; de Kleijn, D.; et al. Concise Review: Developing Best-Practice Models for the Therapeutic Use of Extracellular Vesicles. Stem Cells Transl. Med. 2017, 6, 1730–1739. [Google Scholar] [CrossRef]
- Lee, J.H.; Ha, D.H.; Go, H.-K.; Youn, J.; Kim, H.-K.; Jin, R.C.; Miller, R.B.; Kim, D.-H.; Cho, B.S.; Yi, Y.W. Reproducible Large-Scale Isolation of Exosomes from Adipose Tissue-Derived Mesenchymal Stem/Stromal Cells and Their Application in Acute Kidney Injury. Int. J. Mol. Sci. 2020, 21, 4774. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.-H.; Jeyaraj, M.; Qasim, M.; Kim, J.-H. Review of the Isolation, Characterization, Biological Function, and Multifarious Therapeutic Approaches of Exosomes. Cells 2019, 8, 307. [Google Scholar] [CrossRef]
- Rashid, M.H.; Borin, T.F.; Ara, R.; Alptekin, A.; Liu, Y.; Arbab, A.S. Generation of Novel Diagnostic and Therapeutic Exosomes to Detect and Deplete Protumorigenic M2 Macrophages. Adv. Ther. 2020, 3, 1900209. [Google Scholar] [CrossRef]
- Abusamra, A.J.; Zhong, Z.; Zheng, X.; Li, M.; Ichim, T.E.; Chin, J.L.; Min, W.-P. Tumor exosomes expressing Fas ligand mediate CD8+ T-cell apoptosis. Blood Cells Mol. Dis. 2005, 35, 169–173. [Google Scholar] [CrossRef]
- Andreola, G.; Rivoltini, L.; Castelli, C.; Huber, V.; Perego, P.; Deho, P.; Squarcina, P.; Accornero, P.; Lozupone, F.; Lugini, L.; et al. Induction of lymphocyte apoptosis by tumor cell secretion of FasL-bearing microvesicles. J. Exp. Med. 2002, 195, 1303–1316. [Google Scholar] [CrossRef]
- Kim, J.W.; Wieckowski, E.; Taylor, D.D.; Reichert, T.E.; Watkins, S.; Whiteside, T.L. Fas ligand-positive membranous vesicles isolated from sera of patients with oral cancer induce apoptosis of activated T lymphocytes. Clin. Cancer Res. 2005, 11, 1010–1020. [Google Scholar]
- Huber, V.; Fais, S.; Iero, M.; Lugini, L.; Canese, P.; Squarcina, P.; Zaccheddu, A.; Colone, M.; Arancia, G.; Gentile, M.; et al. Human Colorectal Cancer Cells Induce T-Cell Death Through Release of Proapoptotic Microvesicles: Role in Immune Escape. Gastroenterology 2005, 128, 1796–1804. [Google Scholar] [CrossRef]
- Taylor, D.D.; Gerçel-Taylor, C.; Lyons, K.S.; Stanson, J.; Whiteside, T.L. T-cell apoptosis and suppression of T-cell receptor/CD3-zeta by Fas ligand-containing membrane vesicles shed from ovarian tumors. Clin. Cancer Res. 2003, 9, 5113–5119. [Google Scholar]
- Klinker, M.W.; Lizzio, V.; Reed, T.J.; Fox, D.A.; Lundy, S.K. Human B Cell-Derived Lymphoblastoid Cell Lines Constitutively Produce Fas Ligand and Secrete MHCII(+)FasL(+) Killer Exosomes. Front. Immunol. 2014, 5, 144. [Google Scholar] [CrossRef]
- Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al. Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature 2018, 560, 382–386. [Google Scholar] [CrossRef]
- Chen, J.; Song, Y.; Miao, F.; Chen, G.; Zhu, Y.; Wu, N.; Pang, L.; Chen, Z.; Chen, X. PDL1-positive exosomes suppress antitumor immunity by inducing tumor-specific CD8 + T cell exhaustion during metastasis. Cancer Sci. 2021, 112, 3437–3454. [Google Scholar] [CrossRef]
- Theodoraki, M.-N.; Yerneni, S.S.; Hoffmann, T.K.; Gooding, W.E.; Whiteside, T.L. Clinical Significance of PD-L1+ Exosomes in Plasma of Head and Neck Cancer Patients. Clin. Cancer Res. 2018, 24, 896–905. [Google Scholar] [CrossRef]
- Yang, Y.; Li, C.-W.; Chan, L.-C.; Wei, Y.; Hsu, J.-M.; Xia, W.; Cha, J.-H.; Hou, J.; Hsu, J.L.; Sun, L.; et al. Exosomal PD-L1 harbors active defense function to suppress T cell killing of breast cancer cells and promote tumor growth. Cell Res. 2018, 28, 862–864. [Google Scholar] [CrossRef]
- Wada, J.; Onishi, H.; Suzuki, H.; Yamasaki, A.; Nagai, S.; Morisaki, T.; Katano, M. Surface-bound TGF-beta1 on effusion-derived exosomes participates in maintenance of number and suppressive function of regulatory T-cells in malignant effusions. Anticancer Res. 2010, 30, 3747–3757. [Google Scholar]
- Wang, X.; Shen, H.; Zhangyuan, G.; Huang, R.; Zhang, W.; He, Q.; Jin, K.; Zhuo, H.; Zhang, Z.; Wang, J.; et al. 14-3-3ζ delivered by hepatocellular carcinoma-derived exosomes impaired anti-tumor function of tumor-infiltrating T lymphocytes. Cell Death Dis. 2018, 9, 159. [Google Scholar] [CrossRef]
- Han, Q.; Zhao, H.; Jiang, Y.; Yin, C.; Zhang, J. HCC-Derived Exosomes: Critical Player and Target for Cancer Immune Escape. Cells 2019, 8, 558. [Google Scholar] [CrossRef]
- Yin, C.; Han, Q.; Xu, D.; Zheng, B.; Zhao, X.; Zhang, J. SALL4-mediated upregulation of exosomal miR-146a-5p drives T-cell exhaustion by M2 tumor-associated macrophages in HCC. OncoImmunology 2019, 8, e1601479. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.-C.; Liao, T.-T.; Lin, C.-C.; Yuan, L.-T.E.; Lan, H.-Y.; Lin, H.-H.; Teng, H.-W.; Chang, H.-S.; Lin, C.-H.; Yang, C.-Y.; et al. RAB27B-activated secretion of stem-like tumor exosomes delivers the biomarker microRNA-146a-5p, which promotes tumorigenesis and associates with an immunosuppressive tumor microenvironment in colorectal cancer. Int. J. Cancer 2019, 145, 2209–2224. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Cai, X.; Chen, X.; Liang, H.; Zhang, Y.; Li, J.; Wang, Z.; Chen, X.; Zhang, W.; Yokoyama, S.; et al. Tumor-secreted miR-214 induces regulatory T cells: A major link between immune evasion and tumor growth. Cell Res. 2014, 24, 1164–1180. [Google Scholar] [CrossRef] [PubMed]
- Ashiru, O.; Boutet, P.; Fernández-Messina, L.; Agüera-González, S.; Skepper, J.N.; Vales-Gomez, M.; Reyburn, H.T. Natural Killer Cell Cytotoxicity Is Suppressed by Exposure to the Human NKG2D Ligand MICA*008 That Is Shed by Tumor Cells in Exosomes. Cancer Res. 2010, 70, 481–489. [Google Scholar] [CrossRef]
- Vulpis, E.; Cecere, F.; Molfetta, R.; Soriani, A.; Fionda, C.; Peruzzi, G.; Palchheti, S.; Masuelli, L.; Simonelli, L.; D’Oro, U.; et al. Genotoxic stress modulates the release of exosomes from multiple myeloma cells capable of activating NK cell cytokine production: Role of HSP70/TLR2/NF-kB axis. Oncoimmunology 2017, 6, e1279372. [Google Scholar] [CrossRef]
- Elsner, L.; Muppala, V.; Gehrmann, M.; Lozano-Kuehne, J.; Malzahn, D.; Bickeböller, H.; Brunner, E.; Zientkowska, M.; Herrmann, T.; Walter, L.; et al. The Heat Shock Protein HSP70 Promotes Mouse NK Cell Activity against Tumors That Express Inducible NKG2D Ligands. J. Immunol. 2007, 179, 5523–5533. [Google Scholar] [CrossRef]
- Clayton, A.; Mitchell, J.P.; Court, J.; Linnane, S.; Mason, M.D.; Tabi, Z. Human Tumor-Derived Exosomes Down-Modulate NKG2D Expression. J. Immunol. 2008, 180, 7249–7258. [Google Scholar] [CrossRef]
- Hedlund, M.; Nagaeva, O.; Kargl, D.; Baranov, V.; Mincheva-Nilsson, L. Thermal- and Oxidative Stress Causes Enhanced Release of NKG2D Ligand-Bearing Immunosuppressive Exosomes in Leukemia/Lymphoma T and B Cells. PLoS ONE 2011, 6, e16899. [Google Scholar] [CrossRef]
- Zhao, J.; Schlößer, H.A.; Wang, Z.; Qin, J.; Li, J.; Popp, F.; Popp, M.C.; Alakus, H.; Chon, S.-H.; Hansen, H.P.; et al. Tumor-derived extracellular vesicles inhibit natural killer cell function in pancreatic cancer. Cancers 2019, 11, 874. [Google Scholar] [CrossRef]
- Ye, L.; Zhang, Q.; Cheng, Y.; Chen, X.; Wang, G.; Shi, M.; Zhang, T.; Cao, Y.; Pan, H.; Zhang, L.; et al. Tumor-derived exosomal HMGB1 fosters hepatocellular carcinoma immune evasion by promoting TIM-1+ regulatory B cell expansion. J. Immunother. Cancer 2018, 6, 145. [Google Scholar] [CrossRef]
- Yu, S.; Liu, C.; Su, K.; Wang, J.; Liu, Y.; Zhang, L.; Li, C.; Cong, Y.; Kimberly, R.; Grizzle, W.E.; et al. Tumor Exosomes Inhibit Differentiation of Bone Marrow Dendritic Cells. J. Immunol. 2007, 178, 6867–6875. [Google Scholar] [CrossRef]
- Ding, G.; Zhou, L.; Qian, Y.; Fu, M.; Chen, J.; Chen, J.; Xiang, J.; Wu, Z.; Jiang, G.; Cao, L. Pancreatic cancer-derived exosomes transfer miRNAs to dendritic cells and inhibit RFXAP expression via miR-212-3p. Oncotarget 2015, 6, 29877–29888. [Google Scholar] [CrossRef]
- Ren, W.; Zhang, X.; Li, W.; Feng, Q.; Feng, H.; Tong, Y.; Rong, H.; Wang, W.; Zhang, D.; Zhang, Z.; et al. Exosomal miRNA-107 induces myeloid-derived suppressor cell expansion in gastric cancer. Cancer Manag. Res. 2019, 11, 4023–4040. [Google Scholar] [CrossRef]
- Guo, X.; Qiu, W.; Liu, Q.; Qian, M.; Wang, S.; Zhang, Z.; Gao, X.; Chen, Z.; Xue, H.; Li, G. Immunosuppressive effects of hypoxia-induced glioma exosomes through myeloid-derived suppressor cells via the miR-10a/Rora and miR-21/Pten Pathways. Oncogene 2018, 37, 4239–4259. [Google Scholar] [CrossRef]
- Li, C.; Li, C.; Zhi, C.; Liang, W.; Wang, X.; Chen, X.; Lv, T.; Shen, Q.; Song, Y.; Lin, D.; et al. Clinical significance of PD-L1 expression in serum-derived exosomes in NSCLC patients. J. Transl. Med. 2019, 17, 355. [Google Scholar] [CrossRef]
- Neal, C.L.; Yu, D. 14-3-3ζ as a prognostic marker and therapeutic target for cancer. Expert Opin. Ther. Targets 2010, 14, 1343–1354. [Google Scholar] [CrossRef]
- Zhou, J.; Li, X.; Wu, X.; Zhang, T.; Zhu, Q.; Wang, X.; Wang, H.; Wang, K.; Lin, Y.; Wang, X. Exosomes Released from Tumor-Associated Macrophages Transfer miRNAs That Induce a Treg/Th17 Cell Imbalance in Epithelial Ovarian Cancer. Cancer Immunol. Res. 2018, 6, 1578–1592. [Google Scholar] [CrossRef]
- Zhou, J.; Yang, Y.; Wang, W.; Zhang, Y.; Chen, Z.; Hao, C.; Zhang, J. Melanoma-released exosomes directly activate the mitochondrial apoptotic pathway of CD4+ T cells through their microRNA cargo. Exp. Cell Res. 2018, 371, 364–371. [Google Scholar] [CrossRef]
- Ni, C.; Fang, Q.-Q.; Chen, W.-Z.; Jiang, J.-X.; Jiang, Z.; Ye, J.; Zhang, T.; Yang, L.; Meng, F.-B.; Xia, W.-J.; et al. Breast cancer-derived exosomes transmit lncRNA SNHG16 to induce CD73+γδ1 Treg cells. Signal Transduct. Target. Ther. 2020, 5, 41. [Google Scholar] [CrossRef]
- Di Silvestre, D.; Garavelli, S.; Procaccini, C.; Prattichizzo, F.; Passignani, G.; De Rosa, V.; Mauri, P.; Matarese, G.; de Candia, P. CD4+ T-Cell Activation Prompts Suppressive Function by Extracellular Vesicle-Associated MicroRNAs. Front. Cell Dev. Biol. 2021, 9, 753884. [Google Scholar] [CrossRef]
- Mittelbrunn, M.; Gutiérrez-Vázquez, C.; Villarroya-Beltri, C.; González, S.; Sánchez-Cabo, F.; González, M.Á.; Bernad, A.; Sánchez-Madrid, F. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat. Commun. 2011, 2, 282. [Google Scholar] [CrossRef]
- Batista, I.; Quintas, S.; Melo, S. The Interplay of Exosomes and NK Cells in Cancer Biology. Cancers 2021, 13, 473. [Google Scholar] [CrossRef]
- Liu, C.; Yu, S.; Zinn, K.; Wang, J.; Zhang, L.; Jia, Y.; Kappes, J.C.; Barnes, S.; Kimberly, R.P.; Grizzle, W.E.; et al. Murine mammary carcinoma exosomes promote tumor growth by suppression of NK cell function. J. Immunol. 2006, 176, 1375–1385. [Google Scholar] [CrossRef]
- Bald, T.; Krummel, M.F.; Smyth, M.J.; Barry, K.C. The NK cell–cancer cycle: Advances and new challenges in NK cell–based immunotherapies. Nat. Immunol. 2020, 21, 835–847. [Google Scholar] [CrossRef]
- Pritchard, A.; Tousif, S.; Wang, Y.; Hough, K.; Khan, S.; Strenkowski, J.; Chacko, B.K.; Darley-Usmar, V.M.; Deshane, J.S. Lung Tumor Cell-Derived Exosomes Promote M2 Macrophage Polarization. Cells 2020, 9, 1303. [Google Scholar] [CrossRef]
- Khare, D.; Or, R.; Resnick, I.; Barkatz, C.; Almogi-Hazan, O.; Avni, B. Mesenchymal Stromal Cell-Derived Exosomes Affect mRNA Expression and Function of B-Lymphocytes. Front. Immunol. 2018, 9, 3053. [Google Scholar] [CrossRef]
- Shang, J.; Zha, H.; Sun, Y. Phenotypes, Functions, and Clinical Relevance of Regulatory B Cells in Cancer. Front. Immunol. 2020, 11, 582657. [Google Scholar] [CrossRef]
- Hinata, M.; Kunita, A.; Abe, H.; Morishita, Y.; Sakuma, K.; Yamashita, H.; Seto, Y.; Ushiku, T.; Fukayama, M. Exosomes of Epstein-Barr Virus-Associated Gastric Carcinoma Suppress Dendritic Cell Maturation. Microorgoorganisms 2020, 8, 1776. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, H.; Zeng, Q.; Wang, P.; Zhang, G.; Ji, J.; Li, M.; Shen, S.; Wang, X. Exosomes from 5-aminolevulinic acid photodynamic therapy-treated squamous carcinoma cells promote dendritic cell maturation. Photodiagnosis Photodyn. Ther. 2020, 30, 101746. [Google Scholar] [CrossRef]
- Zhou, M.; Chen, J.; Zhou, L.; Chen, W.; Ding, G.; Cao, L. Pancreatic cancer derived exosomes regulate the expression of TLR4 in dendritic cells via miR-203. Cell. Immunol. 2014, 292, 65–69. [Google Scholar] [CrossRef]
- Tung, S.L.; Boardman, D.A.; Sen, M.; Letizia, M.; Peng, Q.; Cianci, N.; Dioni, L.; Carlin, L.; Lechler, R.; Bollati, V.; et al. Regulatory T cell-derived extracellular vesicles modify dendritic cell function. Sci. Rep. 2018, 8, 6065. [Google Scholar] [CrossRef] [PubMed]
- Torralba, D.; Baixauli, F.; Villarroya-Beltri, C.; Fernández-Delgado, I.; Latorre-Pellicer, A.; Acín-Pérez, R.; Martín-Cófreces, N.B.; Jaso-Tamame, Á.L.; Iborra, S.; Jorge, I.; et al. Priming of dendritic cells by DNA-containing extracellular vesicles from activated T cells through antigen-driven contacts. Nat. Commun. 2018, 9, 2658. [Google Scholar] [CrossRef] [PubMed]
- Kitai, Y.; Kawasaki, T.; Sueyoshi, T.; Kobiyama, K.; Ishii, K.J.; Zou, J.; Akira, S.; Matsuda, T.; Kawai, T. DNA-Containing Exosomes Derived from Cancer Cells Treated with Topotecan Activate a STING-Dependent Pathway and Reinforce Antitumor Immunity. J. Immunol. 2017, 198, 1649–1659. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J.M.; Vanpouille-Box, C.; Spada, S.; Rudqvist, N.-P.; Chapman, J.R.; Ueberheide, B.; Pilones, K.A.; Sarfraz, Y.; Formenti, S.C.; DeMaria, S. Exosomes Shuttle TREX1-Sensitive IFN-Stimulatory dsDNA from Irradiated Cancer Cells to DCs. Cancer Immunol. Res. 2018, 6, 910–920. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef]
- Andre, F.; Escudier, B.; Angevin, E.; Tursz, T.; Zitvogel, L. Exosomes for cancer immunotherapy. Ann. Oncol. 2004, 15, iv141–iv144. [Google Scholar] [CrossRef]
- Sahin, U.; Türeci, Ö. Personalized vaccines for cancer immunotherapy. Science 2018, 359, 1355–1360. [Google Scholar] [CrossRef]
- Taghikhani, A.; Farzaneh, F.; Sharifzad, F.; Mardpour, S.; Ebrahimi, M.; Hassan, Z.M. Engineered Tumor-Derived Extracellular Vesicles: Potentials in Cancer Immunotherapy. Front. Immunol. 2020, 11, 221. [Google Scholar] [CrossRef]
- Dutta, A. Exosomes-based cell-free cancer therapy: A novel strategy for targeted therapy. Immunol. Med. 2020, 44, 116–123. [Google Scholar] [CrossRef]
- Nam, G.; Choi, Y.; Kim, G.B.; Kim, S.; Kim, S.A.; Kim, I. Emerging Prospects of Exosomes for Cancer Treatment: From Conventional Therapy to Immunotherapy. Adv. Mater. 2020, 32, e2002440. [Google Scholar] [CrossRef]
- Vincent-Schneider, H.; Stumptner-Cuvelette, P.; Lankar, D.; Pain, S.; Raposo, G.; Benaroch, P.; Bonnerot, C. Exosomes bearing HLA-DR1 molecules need dendritic cells to efficiently stimulate specific T cells. Int. Immunol. 2002, 14, 713–722. [Google Scholar] [CrossRef]
- Graner, M.W.; Cumming, R.I.; Bigner, D.D. The Heat Shock Response and Chaperones/Heat Shock Proteins in Brain Tumors: Surface Expression, Release, and Possible Immune Consequences. J. Neurosci. 2007, 27, 11214–11227. [Google Scholar] [CrossRef]
- Kunigelis, K.E.; Graner, M.W. The Dichotomy of Tumor Exosomes (TEX) in Cancer Immunity: Is It All in the ConTEXt? Vaccines 2015, 3, 1019–1051. [Google Scholar] [CrossRef]
- Rao, Q.; Zuo, B.; Lu, Z.; Gao, X.; You, A.; Wu, C.; Du, Z.; Yin, H. Tumor-derived exosomes elicit tumor suppression in murine hepatocellular carcinoma models and humans in vitro. Hepatology 2016, 64, 456–472. [Google Scholar] [CrossRef]
- Whiteside, T.L. Stimulatory role of exosomes in the context of therapeutic anti-cancer vaccines. Biotarget 2017, 1, 5. [Google Scholar] [CrossRef]
- Wolfers, J.; Lozier, A.; Raposo, G.; Regnault, A.; Théry, C.; Masurier, C.; Flament, C.; Pouzieux, S.; Faure, F.; Tursz, T.; et al. Tumor-derived exosomes are a source of shared tumor rejection antigens for CTL cross-priming. Nat. Med. 2001, 7, 297–303. [Google Scholar] [CrossRef]
- Gao, J.; Qiu, X.; Li, X.; Fan, H.; Zhang, F.; Lv, T.; Song, Y. Expression profiles and clinical value of plasma exosomal Tim-3 and Galectin-9 in non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2018, 498, 409–415. [Google Scholar] [CrossRef]
- Farhood, B.; Najafi, M.; Mortezaee, K. CD8+cytotoxic T lymphocytes in cancer immunotherapy: A review. J. Cell. Physiol. 2019, 234, 8509–8521. [Google Scholar] [CrossRef]
- Poggio, M.; Hu, T.; Pai, C.-C.; Chu, B.; Belair, C.D.; Chang, A.; Montabana, E.; Lang, U.E.; Fu, Q.; Fong, L.; et al. Suppression of exosomal PD-L1 induces systemic anti-tumor immunity and memory. Cell 2019, 177, 414–427.e13. [Google Scholar] [CrossRef]
- Hou, W.; Yuan, Q.; Yuan, X.; Wang, Y.; Mo, W.; Wang, H.; Yu, M. A novel tetravalent bispecific antibody targeting programmed death 1 and tyrosine-protein kinase Met for treatment of gastric cancer. Investig. New Drugs 2018, 37, 876–889. [Google Scholar] [CrossRef]
- Tavasolian, F.; Hosseini, A.Z.; Rashidi, M.; Soudi, S.; Abdollahi, E.; Momtazi-Borojeni, A.A.; Sathyapalan, T.; Sahebkar, A. The Impact of Immune Cell-derived Exosomes on Immune Response Initiation and Immune System Function. Curr. Pharm. Des. 2021, 27, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Vatter, F.A.P.; Cioffi, M.; Hanna, S.J.; Castarede, I.; Caielli, S.; Pascual, V.; Matei, I.; Lyden, D. Extracellular vesicle–and particle-mediated communication shapes innate and adaptive immune responses. J. Exp. Med. 2021, 218, e20202579. [Google Scholar] [CrossRef] [PubMed]
- Buschow, S.I.; Nolte-‘t Hoen, E.N.M.N.; Van Niel, G.; Pols, M.S.; ten Broeke, T.T.; Lauwen, M.; Ossendorp, F.; Melief, C.J.M.; Raposo, G.; Wubbolts, R.; et al. MHC II in dendritic cells is targeted to lysosomes or t cell-induced exosomes via distinct multivesicular body pathways. Traffic 2009, 10, 1528–1542. [Google Scholar] [CrossRef] [PubMed]
- Amigorena, S. Cancer immunotherapy using dendritic cell-derived exosomes. Medicina 2000, 60, 51–54. [Google Scholar]
- Muntasell, A.; Berger, A.C.; Roche, P.A. T cell-induced secretion of MHC class II–peptide complexes on B cell exosomes. EMBO J. 2007, 26, 4263–4272. [Google Scholar] [CrossRef]
- Mellman, I. Dendritic Cells: Master Regulators of the Immune Response. Cancer Immunol. Res. 2013, 1, 145–149. [Google Scholar] [CrossRef]
- Tai, Y.; Wang, Q.; Korner, H.; Zhang, L.; Wei, W. Molecular Mechanisms of T Cells Activation by Dendritic Cells in Autoimmune Diseases. Front. Pharmacol. 2018, 9, 642. [Google Scholar] [CrossRef]
- Nolte-’t Hoen, E.N.M.; Buschow, S.I.; Anderton, S.M.; Stoorvogel, W.; Wauben, M.H.M. Activated T cells recruit exosomes secreted by dendritic cells via LFA-1. Blood 2009, 113, 1977–1981. [Google Scholar] [CrossRef]
- Gutierrez-Vazquez, C.; Villarroya-Beltri, C.; Mittelbrunn, M.; Sánchez-Madrid, F. Transfer of extracellular vesicles during immune cell-cell interactions. Immunol. Rev. 2012, 251, 125–142. [Google Scholar] [CrossRef]
- Segura, E.; Nicco, C.; Lombard, B.; Véron, P.; Raposo, G.; Batteux, F.; Amigorena, S.; Théry, C. ICAM-1 on exosomes from mature dendritic cells is critical for efficient naive T-cell priming. Blood 2005, 106, 216–223. [Google Scholar] [CrossRef]
- Wei, G.; Jie, Y.; Haibo, L.; Chaoneng, W.; Dong, H.; Jianbing, Z.; Junjie, G.; Leilei, M.; Hongtao, S.; Yunzeng, Z.; et al. Dendritic cells derived exosomes migration to spleen and induction of inflammation are regulated by CCR7. Sci. Rep. 2017, 7, srep42996. [Google Scholar] [CrossRef]
- Munich, S.; Sobo-Vujanovic, A.; Buchser, W.J.; Beer-Stolz, D.; Vujanovic, N.L. Dendritic cell exosomes directly kill tumor cells and activate natural killer cells via TNF superfamily ligands. OncoImmunology 2012, 1, 1074–1083. [Google Scholar] [CrossRef]
- Viaud, S.; Terme, M.; Flament, C.; Taieb, J.; Andre, F.; Novault, S.; Escudier, B.; Robert, C.; Caillat-Zucman, S.; Tursz, T.; et al. Dendritic Cell-Derived Exosomes Promote Natural Killer Cell Activation and Proliferation: A Role for NKG2D Ligands and IL-15Rα. PLoS ONE 2009, 4, e4942. [Google Scholar] [CrossRef]
- Elashiry, M.; Elashiry, M.M.; Elsayed, R.; Rajendran, M.; Auersvald, C.; Zeitoun, R.; Rshid, M.H.; Ara, R.; Meghil, M.M.; Liu, Y.; et al. Dendritic cell derived exosomes loaded with immunoregulatory cargo reprogram local immune responses and inhibit degenerative bone disease in vivo. J. Extracell. Vesicles 2020, 9, 1795362. [Google Scholar] [CrossRef]
- Lindenbergh, M.F.S.; Stoorvogel, W. Antigen presentation by extracellular vesicles from professional antigen-presenting Cells. Annu. Rev. Immunol. 2018, 36, 435–459. [Google Scholar] [CrossRef]
- Mauri, C.; Bosma, A. Immune Regulatory Function of B Cells. Annu. Rev. Immunol. 2012, 30, 221–241. [Google Scholar] [CrossRef]
- Arita, S.; Baba, E.; Shibata, Y.; Niiro, H.; Shimoda, S.; Isobe, T.; Kusaba, H.; Nakano, S.; Harada, M. B cell activation regulates exosomal HLA production. Eur. J. Immunol. 2008, 38, 1423–1434. [Google Scholar] [CrossRef]
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.J.; Geuze, H.J. B lymphocytes secrete antigen-presenting vesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef]
- Zou, J.; Peng, H.; Liu, Y. The Roles of Exosomes in Immunoregulation and Autoimmune Thyroid Diseases. Front. Immunol. 2021, 12, 4283. [Google Scholar] [CrossRef]
- Saunderson, S.C.; McLellan, A.D. Role of lymphocyte subsets in the immune response to primary B cell–derived exosomes. J. Immunol. 2017, 199, 2225–2235. [Google Scholar] [CrossRef]
- Kato, T.; Fahrmann, J.F.; Hanash, S.M.; Vykoukal, J. Extracellular Vesicles Mediate B Cell Immune Response and Are a Potential Target for Cancer Therapy. Cells 2020, 9, 1518. [Google Scholar] [CrossRef]
- Clayton, A.; Turkes, A.; DeWitt, S.; Steadman, R.; Mason, M.D.; Hallett, M.B. Adhesion and signaling by B cell-derived exosomes: The role of integrins. FASEB J. 2004, 18, 977–979. [Google Scholar] [CrossRef]
- Papp, K.; Vegh, P.; Prechl, J.; Kerekes, K.; Kovács, J.; Csikós, G.; Bajtay, Z.; Erdei, A. B lymphocytes and macrophages release cell membrane deposited C3-fragments on exosomes with T cell response-enhancing capacity. Mol. Immunol. 2008, 45, 2343–2351. [Google Scholar] [CrossRef]
- Rabb, H. The T cell as a bridge between innate and adaptive immune systems: Implications for the kidney. Kidney Int. 2002, 61, 1935–1946. [Google Scholar] [CrossRef]
- Anel, A.; Gallego-Lleyda, A.; De Miguel, D.; Naval, J.; Martínez-Lostao, L. Role of Exosomes in the Regulation of T-cell Mediated Immune Responses and in Autoimmune Disease. Cells 2019, 8, 154. [Google Scholar] [CrossRef]
- Martínez-Lorenzo, M.J.; Anel, A.; Gamen, S.; Monein, N.; Lasierra, P.; Larrad, L.; Piñeiro, A.; Alava, A.M.; Naval, J. Activated human T cells release bioactive Fas ligand and APO2 ligand in microvesicles. J. Immunol. 1999, 163, 1274–1281. [Google Scholar]
- Hong, X.; Schouest, B.; Xu, H. Effects of exosome on the activation of CD4+ T cells in rhesus macaques: A potential application for HIV latency reactivation. Sci. Rep. 2017, 7, 15611. [Google Scholar] [CrossRef]
- Ito, H.; Seishima, M. Regulation of the Induction and Function of Cytotoxic T Lymphocytes by Natural Killer T Cell. J. Biomed. Biotechnol. 2010, 2010, 641757. [Google Scholar] [CrossRef]
- Li, L.; Jay, S.M.; Wang, Y.; Wu, S.-W.; Xiao, Z. IL-12 stimulates CTLs to secrete exosomes capable of activating bystander CD8+ T cells. Sci. Rep. 2017, 7, 13365. [Google Scholar] [CrossRef]
- Li, P.; Liu, C.; Yu, Z.; Wu, M. New Insights into Regulatory T Cells: Exosome- and Non-Coding RNA-Mediated Regulation of Homeostasis and Resident Treg Cells. Front. Immunol. 2016, 7, 574. [Google Scholar] [CrossRef]
- Okoye, I.S.; Coomes, S.M.; Pelly, V.S.; Czieso, S.; Papayannopoulos, V.; Tolmachova, T.; Seambra, M.C.; Wilson, M.C. MicroRNA-Containing T-Regulatory-Cell-Derived Exosomes Suppress Pathogenic T Helper 1 Cells. Immunity 2014, 41, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Veglia, F.; Sanseviero, E.; Gabrilovich, D.I. Myeloid-derived suppressor cells in the era of increasing myeloid cell diversity. Nat. Rev. Immunol. 2021, 21, 485–498. [Google Scholar] [CrossRef]
- Atretkhany, K.-S.N.; Drutskaya, M.S. Myeloid-derived suppressor cells and proinflammatory cytokines as targets for cancer therapy. Biochemistry 2016, 81, 1274–1283. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.; Choksawangkarn, W.; Edwards, N.; Ostrand-Rosenberg, S.; Fenselau, C. Exosomes from Myeloid-Derived Suppressor Cells Carry Biologically Active Proteins. J. Proteome Res. 2013, 13, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.; Sinha, P.; Horn, L.A.; Clements, V.K.; Yang, H.; Li, J.; Tracey, K.J.; Ostrand-Rosenberg, S. HMGB1 Enhances Immune Suppression by Facilitating the Differentiation and Suppressive Activity of Myeloid-Derived Suppressor Cells. Cancer Res. 2014, 74, 5723–5733. [Google Scholar] [CrossRef]
- Rashid, M.H.; Borin, T.F.; Ara, R.; Piranlioglu, R.; Achyut, B.R.; Korkaya, H.; Liu, Y.; Arbab, A.S. Critical immunosuppressive effect of MDSC-derived exosomes in the tumor microenvironment. Oncol. Rep. 2021, 45, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Geis-Asteggiante, L.; Belew, A.T.; Clements, V.K.; Edwards, N.J.; Ostrand-Rosenberg, S.; El-Sayed, N.M.; Fenselau, C. Differential Content of Proteins, mRNAs, and miRNAs Suggests that MDSC and Their Exosomes May Mediate Distinct Immune Suppressive Functions. J. Proteome Res. 2017, 17, 486–498. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tian, J.; Tang, X.; Rui, K.; Tian, X.; Ma, J.; Ma, B.; Xu, H.; Lu, L.; Wang, S. Exosomes released by granulocytic myeloid-derived suppressor cells attenuate DSS-induced colitis in mice. Oncotarget 2016, 7, 15356–15368. [Google Scholar] [CrossRef]
- Sinha, P.; Clements, V.K.; Bunt, S.K.; Albelda, S.M.; Ostrand-Rosenberg, S. Cross-Talk between Myeloid-Derived Suppressor Cells and Macrophages Subverts Tumor Immunity toward a Type 2 Response. J. Immunol. 2007, 179, 977–983. [Google Scholar] [CrossRef]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef]
- Nakae, S.; Suto, H.; Kakurai, M.; Sedgwick, J.D.; Tsai, M.; Galli, S.J. Mast cells enhance T cell activation: Importance of mast cell-derived TNF. Proc. Natl. Acad. Sci. USA 2005, 102, 6467–6472. [Google Scholar] [CrossRef]
- Palma, A.M.; Hanes, M.R.; Marshall, J.S. Mast Cell Modulation of B Cell Responses: An Under-Appreciated Partnership in Host Defence. Front. Immunol. 2021, 12, 3677. [Google Scholar] [CrossRef]
- Exosomes, I.A. Mast Cell-Dependent B and T Lymphocyte. J. Immunol. 2001, 166, 868–876. [Google Scholar]
- Li, F.; Wang, Y.; Lin, L.; Wang, J.; Xiao, H.; Li, J.; Peng, X.; Dai, H.; Li, L. Mast Cell-Derived Exosomes Promote Th2 Cell Differentiation via OX40L-OX40 Ligation. J. Immunol. Res. 2016, 2016, 3623898. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, X.; Wang, M.; Chen, Z.; Yan, Y.; Gu, W.; Tan, J.; Jiang, W.; Ji, W. Exosomes from Thymic Stromal Lymphopoietin-Activated Dendritic Cells Promote Th2 Differentiation through the OX40 Ligand. Pathobiology 2018, 86, 111–117. [Google Scholar] [CrossRef]
- Xie, G.; Yang, H.; Peng, X.; Lin, L.; Wang, J.; Lin, K.; Cui, Z.; Li, J.; Xiao, H.; Liang, Y.; et al. Mast cell exosomes can suppress allergic reactions by binding to IgE. J. Allergy Clin. Immunol. 2017, 141, 788–791. [Google Scholar] [CrossRef]
- Molfetta, R.; Lecce, M.; Quatrini, L.; Caracciolo, G.; Digiacomo, L.; Masuelli, L.; Milito, N.D.; Vulpis, E.; Zingoni, A.; Galandrini, R.; et al. Immune complexes exposed on mast cell-derived nanovesicles amplify allergic inflammation. Allergy 2019, 75, 1260–1263. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef]
- Vargas, A.; Roux-Dalvai, F.; Droit, A.; Lavoie, J.-P. Neutrophil-Derived Exosomes: A New Mechanism Contributing to Airway Smooth Muscle Remodeling. Am. J. Respir. Cell Mol. Biol. 2016, 55, 450–461. [Google Scholar] [CrossRef]
- Shao, S.; Fang, H.; Zhang, J.; Jiang, M.; Xue, K.; Ma, J.; Zhang, J.; Lei, J.; Zhang, Y.; Li, B.; et al. Neutrophil exosomes enhance the skin autoinflammation in generalized pustular psoriasis via activating keratinocytes. FASEB J. 2019, 33, 6813–6828. [Google Scholar] [CrossRef]
- Eken, C.; Gasser, O.; Zenhaeusern, G.; Oehri, I.; Hess, C.; Schifferli, J.A. Polymorphonuclear Neutrophil-Derived Ectosomes Interfere with the Maturation of Monocyte-Derived Dendritic Cells. J. Immunol. 2008, 180, 817–824. [Google Scholar] [CrossRef]
- Majumdar, R.; Tameh, A.T.; Parent, C.A. Exosomes Mediate LTB4 Release during Neutrophil Chemotaxis. PLoS Biol. 2016, 14, e1002336. [Google Scholar] [CrossRef]
- Duarte, T.A.; Noronha-Dutra, A.A.; Nery, J.S.; Ribeiro, S.B.; Pitanga, T.N.; e Silva, J.R.L.; Arruda, S.; Boéchat, N. Mycobacterium tuberculosis-induced neutrophil ectosomes decrease macrophage activation. Tuberculosis 2012, 92, 218–225. [Google Scholar] [CrossRef]
- Shen, G.; Krienke, S.; Schiller, P.; Nießen, A.; Neu, S.; Eckstein, V.; Schiler, M.; Lorenz, H.-M.; Tykocinski, L.-O. Microvesicles released by apoptotic human neutrophils suppress proliferation and IL-2/IL-2 receptor expression of resting T helper cells. Eur. J. Immunol. 2017, 47, 900–910. [Google Scholar] [CrossRef]
- Abel, A.M.; Yang, C.; Thakar, M.S.; Malarkannan, S. Natural Killer Cells: Development, Maturation, and Clinical Utilization. Front. Immunol. 2018, 9, 1869. [Google Scholar] [CrossRef]
- Lugini, L.; Cecchetti, S.; Huber, V.; Luciani, F.; Macchia, G.; Spadaro, F.; Paris, L.; Abalsamo, L.; Colone, M.; Molinari, A.; et al. Immune surveillance properties of human NK cell-derived exosomes. J. Immunol. 2012, 189, 2833–2842. [Google Scholar] [CrossRef]
- Neviani, P.; Wise, P.M.; Murtadha, M.; Liu, C.W.; Wu, C.-H.; Jong, A.Y.; Seeger, R.C.; Fabbri, M. Natural killer–derived exosomal miR-186 inhibits neuroblastoma growth and immune escape mechanisms. Cancer Res. 2019, 79, 1151–1164. [Google Scholar] [CrossRef]
- Federici, C.; Shahaj, E.; Cecchetti, S.; Camerini, S.; Casella, M.; Iessi, E.; Camisaschi, C.; Paolino, G.; Calvieri, S.; Ferro, S.; et al. Natural-killer-derived extracellular vesicles: Immune sensors and interactors. Front. Immunol. 2020, 11, 262. [Google Scholar] [CrossRef] [PubMed]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Schraufstatter, I.U.; Khaldoyanidi, S.K.; Discipio, R.G. Complement activation in the context of stem cells and tissue repair. World J. Stem Cells 2015, 7, 1090–1108. [Google Scholar] [CrossRef] [PubMed]
- Sarma, J.V.; Ward, P.A. The complement system. Cell Tissue Res. 2011, 343, 227–235. [Google Scholar] [CrossRef]
- Taylor, P.; Botto, M.; Walport, M. The complement system. Curr. Biol. 1998, 8, R259–R261. [Google Scholar] [CrossRef][Green Version]
- Unnewehr, H.; Rittirsch, D.; Sarma, J.V.; Zetoune, F.; Flierl, M.A.; Perl, M.; Denk, S.; Weiss, M.; Schneider, M.E.; Monk, P.N.; et al. Changes and Regulation of the C5a Receptor on Neutrophils during Septic Shock in Humans. J. Immunol. 2013, 190, 4215–4225. [Google Scholar] [CrossRef]
- Clayton, A.; Harris, C.L.; Court, J.; Mason, M.D.; Morgan, P. Antigen-presenting cell exosomes are protected from complement-mediated lysis by expression of CD55 and CD59. Eur. J. Immunol. 2003, 33, 522–531. [Google Scholar] [CrossRef]
- Brodbeck, W.G.; Kuttner-Kondo, L.; Mold, C.; Medof, M.E. Structure/function studies of human decay-accelerating factor. Immunology 2000, 101, 104–111. [Google Scholar] [CrossRef]
- Gasser, O.; Schifferli, J.A. Microparticles released by human neutrophils adhere to erythrocytes in the presence of complement. Exp. Cell Res. 2005, 307, 381–387. [Google Scholar] [CrossRef]
- Pascual, M.; Lutz, H.U.; Steiger, G.; Stammler, P.; Schifferli, J.A. Release of vesicles enriched in complement receptor 1 from human erythrocytes. J. Immunol. 1993, 151, 397–404. [Google Scholar]
- Biró, E.; Nieuwland, R.; Tak, P.P.; Pronk, L.M.; Schaap, M.C.L.; Sturk, A.; Hack, C.E. Activated complement components and complement activator molecules on the surface of cell-derived microparticles in patients with rheumatoid arthritis and healthy individuals. Ann. Rheum. Dis. 2007, 66, 1085–1092. [Google Scholar] [CrossRef]
- Stein, J.M.; Luzio, J.P. Ectocytosis caused by sublytic autologous complement attack on human neutrophils. The sorting of endogenous plasma-membrane proteins and lipids into shed vesicles. Biochem. J. 1991, 274, 381–386. [Google Scholar] [CrossRef]
- Iida, K.; Whitlow, M.B.; Nussenzweig, V. Membrane vesiculation protects erythrocytes from destruction by complement. J. Immunol. 1991, 147, 2638–2642. [Google Scholar]
- Volanakis, E.J. Human C-reactive protein: Expression, structure, and function. Mol. Immunol. 2001, 38, 189–197. [Google Scholar] [CrossRef]
- Braig, D.; Nero, T.; Koch, H.-G.; Kaiser, B.; Wang, X.; Thiele, J.R.; Morton, C.; Zeller, J.; Kiefer, J.; Potempa, L.A.; et al. Transitional changes in the CRP structure lead to the exposure of proinflammatory binding sites. Nat. Commun. 2017, 8, 14188. [Google Scholar] [CrossRef]
- Braig, D.; Kaiser, B.; Thiele, J.R.; Bannasch, H.; Peter, K.; Stark, G.B.; Koch, H.-G.; Eisenhardt, S.U. A conformational change of C-reactive protein in burn wounds unmasks its proinflammatory properties. Int. Immunol. 2014, 26, 467–478. [Google Scholar] [CrossRef]
- Loh, J.T.; Zhang, B.; Teo, J.K.H.; Lai, R.C.; Choo, A.B.H.; Lam, K.-P.; Lim, S.K. Mechanism for the attenuation of neutrophil and complement hyperactivity by MSC exosomes. Cytotherapy 2022. [Google Scholar] [CrossRef]
- Kamerkar, S.; LeBleu, V.S.; Sugimoto, H.; Yang, S.; Ruivo, C.; Melo, S.; Lee, J.J.; Kalluri, R. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature 2017, 546, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Bohana-Kashtan, O.; Pinna, L.A.; Fishelson, Z. Extracellular phosphorylation of C9 by protein kinase CK2 regulates complement-mediated lysis. Eur. J. Immunol. 2005, 35, 1939–1948. [Google Scholar] [CrossRef]
- Riteau, B.; Faure, F.; Menier, C.; Viel, S.; Carosella, E.D.; Amigorena, S.; Rouas-Freiss, N. Exosomes bearing HLA-G are released by melanoma cells. Hum. Immunol. 2003, 64, 1064–1072. [Google Scholar] [CrossRef]
- Le Gal, F.-A.; Riteau, B.; Sedlik, C.; Khalil-Daher, I.; Menier, C.; Dausset, J.; Guillet, J.-G.; Carosella, E.D.; Rouas-Freiss, N. HLA-G-mediated inhibition of antigen-specific cytotoxic T lymphocytes. Int. Immunol. 1999, 11, 1351–1356. [Google Scholar] [CrossRef]
- Riteau, B.; Rouas-Freiss, N.; Menier, C.; Paul, P.; Dausset, J.; Carosella, E.D. HLA-G2, -G3, and -G4 Isoforms Expressed as Nonmature Cell Surface Glycoproteins Inhibit NK and Antigen-Specific CTL Cytolysis. J. Immunol. 2001, 166, 5018–5026. [Google Scholar] [CrossRef] [PubMed]
- Chaput, N.; Angevin, E.; Zitvogel, L. Exosome-based immunotherapy. Cancer Immunol. Immunother. 2004, 53, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Lundy, S.K.; Klinker, M.W.; Fox, D.A. Killer B Lymphocytes and Their Fas Ligand Positive Exosomes as Inducers of Immune Tolerance. Front. Immunol. 2015, 6, 122. [Google Scholar] [CrossRef]
- Li, K.; Chang, S.; Wang, Z.; Zhao, X.; Chen, D. A novel micro-emulsion and micelle assembling method to prepare DEC205 monoclonal antibody coupled cationic nanoliposomes for simulating exosomes to target dendritic cells. Int. J. Pharm. 2015, 491, 105–112. [Google Scholar] [CrossRef]
- De La Peña, H.; Madrigal, A.; Rusakiewicz, S.; Bencsik, M.; Cave, G.W.; Selman, A.; Rees, R.C.; Travers, P.J.; Dodi, I.A. Artificial exosomes as tools for basic and clinical immunology. J. Immunol. Methods 2009, 344, 121–132. [Google Scholar] [CrossRef]
- Zhao, L.; Gu, C.; Gana, Y.; Shaoa, L.; Chenc, H.; Zhua, H. Exosome-mediated siRNA delivery to suppress postoperative breast cancer metastasis. J. Control. Release 2020, 318, 1–15. [Google Scholar] [CrossRef]
- Cheng, Q.; Shi, X.; Han, M.; Smbatyan, G.; Lenz, H.-J.; Zhang, Y. Reprogramming Exosomes as Nanoscale Controllers of Cellular Immunity. J. Am. Chem. Soc. 2018, 140, 16413–16417. [Google Scholar] [CrossRef]
- Qi, H.; Liu, C.; Long, L.; Ren, Y.; Zhang, S.; Chang, X.; Qian, X.; Jia, H.; Zhao, J.; Sun, J.; et al. Blood Exosomes Endowed with Magnetic and Targeting Properties for Cancer Therapy. ACS Nano 2016, 10, 3323–3333. [Google Scholar] [CrossRef]
- Shi, X.; Cheng, Q.; Hou, T.; Han, M.; Smbatyan, G.; Lang, J.; Epstein, A.L.; Lenz, H.-J.; Zhang, Y. Genetically Engineered Cell-Derived Nanoparticles for Targeted Breast Cancer Immunotherapy. Mol. Ther. 2020, 28, 536–547. [Google Scholar] [CrossRef]
- Hu, W.; Huang, F.; Ning, L.; Hao, J.; Wan, J.; Hao, S. Enhanced immunogenicity of leukemia-derived exosomes via transfection with lentiviral vectors encoding costimulatory molecules. Cell. Oncol. 2020, 43, 889–900. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Koyama, Y.; Ito, T.; Hasegawa, A.; Eriguchi, M.; Inaba, T.; Ushigusa, T.; Sugiura, K. Exosomes derived from tumor cells genetically modified to express Mycobacterium tuberculosis antigen: A novel vaccine for cancer therapy. Biotechnol. Lett. 2016, 38, 1857–1866. [Google Scholar] [CrossRef]
- Morishita, M.; Takahashi, Y.; Matsumoto, A.; Nishikawa, M.; Takakura, Y. Exosome-based tumor antigens–adjuvant co-delivery utilizing genetically engineered tumor cell-derived exosomes with immunostimulatory CpG DNA. Biomaterials 2016, 111, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Regnault, A.; Lozier, A.; Wolfers, J.; Flament, C.; Tenza, D.; Ricciardi-Castagnoli, P.; Raposo, G.; Amigorena, S. Eradication of established murine tumors using a novel cell-free vaccine: Dendritic cell derived exosomes. Nat. Med. 1998, 4, 594–600. [Google Scholar] [CrossRef]
- Wang, J.; Wang, L.; Lin, Z.; Tao, L.; Chen, M. More efficient induction of antitumor T cell immunity by exosomes from CD40L gene-modified lung tumor cells. Mol. Med. Rep. 2013, 9, 125–131. [Google Scholar] [CrossRef]
- Zuo, B.; Qi, H.; Lu, Z.; Chen, L.; Sun, B.; Yang, R.; Zhang, Y.; Liu, Z.; Gao, X.; You, A.; et al. Alarmin-painted exosomes elicit persistent antitumor immunity in large established tumors in mice. Nat. Commun. 2020, 11, 1790. [Google Scholar] [CrossRef]
- Koh, E.; Lee, E.J.; Nam, G.-H.; Hong, Y.; Cho, E.; Yang, Y.; Kim, I.-S. Exosome-SIRPα, a CD47 blockade increases cancer cell phagocytosis. Biomaterials 2017, 121, 121–129. [Google Scholar] [CrossRef]
- Shi, X.; Sun, J.; Li, H.; Lin, H.; Xie, W.; Li, J.; Tan, W. Antitumor efficacy of interferon-γ-modified exosomal vaccine in prostate cancer. Prostate 2020, 80, 811–823. [Google Scholar] [CrossRef] [PubMed]
- Duong, N.; Curley, K.; Brown, A.; Campanelli, A.; Do, M.A.; Levy, D.; Tantry, A.; Marriott, G.; Lu, B. Decoy exosomes as a novel biologic reagent to antagonize inflammation. Int. J. Nanomed. 2019, 14, 3413–3425. [Google Scholar] [CrossRef] [PubMed]
- Hannafon, B.N.; Carpenter, K.J.; Berry, W.L.; Janknecht, R.; Dooley, W.C.; Ding, W.-Q. Exosome-mediated microRNA signaling from breast cancer cells is altered by the anti-angiogenesis agent docosahexaenoic acid (DHA). Mol. Cancer 2015, 14, 133. [Google Scholar] [CrossRef]
- Taghikhani, A.; Hassan, Z.M.; Ebrahimi, M.; Moazzeni, S.-M. microRNA modified tumor-derived exosomes as novel tools for maturation of dendritic cells. J. Cell. Physiol. 2019, 234, 9417–9427. [Google Scholar] [CrossRef]
- Que, R.-S.; Lin, C.; Ding, G.P.; Wu, Z.R.; Cao, L.P. Increasing the immune activity of exosomes: The effect of miRNA-depleted exosome proteins on activating dendritic cell/cytokine-induced killer cells against pancreatic cancer. J. Zhejiang Univ. Sci. B 2016, 17, 352–360. [Google Scholar] [CrossRef]
- Peng, X.-X.; Yu, R.; Wu, X.; Wu, S.Y.; Pi, C.; Chen, Z.H.; Zhang, X.C.; Gao, C.Y.; Shao, Y.W.; Liu, L.; et al. Correlation of plasma exosomal microRNAs with the efficacy of immunotherapy in EGFR/ALK wild-type advanced non-small cell lung cancer. J. Immunother. Cancer 2020, 8, e000376. [Google Scholar] [CrossRef]
- Liu, J.; Yoo, J.; Ho, J.Y.; Jung, Y.; Lee, S.; Hur, S.Y.; Choi, Y.J. Plasma-derived exosomal miR-4732-5p is a promising noninvasive diagnostic biomarker for epithelial ovarian cancer. J. Ovarian Res. 2021, 14, 59. [Google Scholar] [CrossRef]
- Gehrmann, U.; Näslund, T.I.; Hiltbrunner, S.; Larssen, P.; Gabrielsson, S. Harnessing the exosome-induced immune response for cancer immunotherapy. Semin. Cancer Biol. 2014, 28, 58–67. [Google Scholar] [CrossRef]
- Salanti, A.; Clausen, T.M.; Agerbæk, M.Ø.; Nakouzi, N.A.; Dahlbäck, M.; Oo, H.Z.; Lee, S.; Gustavsson, T.; Rich, J.R.; Hedberg, B.J.; et al. Targeting Human Cancer by a Glycosaminoglycan Binding Malaria Protein. Cancer Cells 2015, 28, 500–514. [Google Scholar] [CrossRef]
- Yim, N.; Ryu, S.-W.; Choi, K.; Lee, K.R.; Lee, S.; Choi, H.; Kim, J.; Shaker, M.R.; Sun, W.; Park, J.-H.; et al. Exosome engineering for efficient intracellular delivery of soluble proteins using optically reversible protein–protein interaction module. Nat. Commun. 2016, 7, 12277. [Google Scholar] [CrossRef]
- Yang, M.-Q.; Du, Q.; Varley, P.R.; Goswami, J.; Liang, Z.; Wang, R.; Li, H.; Stolz, D.B.; Geller, D.A. Interferon regulatory factor 1 priming of tumour-derived exosomes enhances the antitumour immune response. Br. J. Cancer 2018, 118, 62–71. [Google Scholar] [CrossRef]
- Sato, Y.T.; Umezaki, K.; Sawada, S.; Mukai, S.-A.; Sasaki, Y.; Harada, N.; Shiku, H.; Akiyoshi, K. Engineering hybrid exosomes by membrane fusion with liposomes. Sci. Rep. 2016, 6, 21933. [Google Scholar] [CrossRef]
- Hung, M.E.; Leonard, J.N. Stabilization of exosome-targeting peptides via engineered glycosylation. J. Biol. Chem. 2015, 290, 8166–8172. [Google Scholar] [CrossRef]
- Zeng, Y.; Graner, M.W.; Katsanis, E. Chaperone-rich cell lysates, immune activation and tumor vaccination. Cancer Immunol. Immunother. 2005, 55, 329–338. [Google Scholar] [CrossRef]
- Hood, J.L. Post isolation modification of exosomes for nanomedicine applications. Nanomedicine 2016, 11, 1745–1756. [Google Scholar] [CrossRef]
- Carobolante, G.; Mantaj, J.; Ferrari, E.; Vllasaliu, D. Cow Milk and Intestinal Epithelial Cell-Derived Extracellular Vesicles as Systems for Enhancing Oral Drug Delivery. Pharmaceutics 2020, 12, 226. [Google Scholar] [CrossRef]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016, 371, 48–61. [Google Scholar] [CrossRef]
- Agrawal, A.; Aqil, F.; Jeyabalan, J.; Spencer, W.A.; Beck, J.; Gachuki, B.W.; Alhakeem, S.; Oben, K.; Munagala, R.; Bondada, S.; et al. Milk-derived exosomes for oral delivery of paclitaxel. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 1627–1636. [Google Scholar] [CrossRef] [PubMed]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Gupta, R.C. Abstract 5407: Milk derived exosomes: Scalable source of biologically active drug delivery nanoparticles. Cancer Chem. 2014, 74, 5407. [Google Scholar]
- Cai, Q.; Zhu, A.; Gong, L. Exosomes of glioma cells deliver miR-148a to promote proliferation and metastasis of glioblastoma via targeting CADM1. Bull. Cancer 2018, 105, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.-S.; Sharma, P.; Yerneni, S.S.; Simms, P.; Jackson, E.K.; Whiteside, T.L.; Boyiadzis, M. Circulating exosomes carrying an immunosuppressive cargo interfere with cellular immunotherapy in acute myeloid leukemia. Sci. Rep. 2017, 7, 14684. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Wang, Y.; Xia, X.; Zheng, J.C. Exosome engineering: Current progress in cargo loading and targeted delivery. NanoImpact 2020, 20, 100261. [Google Scholar] [CrossRef]
- Yamashita, T.; Kamada, H.; Kanasaki, S.; Maeda, Y.; Nagano, K.; Abe, Y.; Yoshioka, Y.; Tsutsumi, Y.; Katayama, S.; Inoue, M.; et al. Epidermal growth factor receptor localized to exosome membranes as a possible biomarker for lung cancer diagnosis. Pharm. Int. J. Pharm. Sci. 2013, 68, 969–973. [Google Scholar]
- Van Giau, V.; An, S.S.A. Emergence of exosomal miRNAs as a diagnostic biomarker for Alzheimer’s disease. J. Neurol. Sci. 2016, 360, 141–152. [Google Scholar] [CrossRef]
- Hosseini, M.; Khatamianfar, S.; Hassanian, S.M.; Nedaeinia, R.; Shafiee, M.; Maftouh, M.; Ghayour-Mobarhan, M.; Shahid Sales, S.; Avan, A. Exosome-Encapsulated microRNAs as Potential Circulating Biomarkers in Colon Cancer. Curr. Pharm. Des. 2017, 23, 1705–1709. [Google Scholar] [CrossRef]
- Xing, X.; Han, S.; Li, Z.; Li, Z. Emerging role of exosomes in craniofacial and dental applications. Theranostics 2020, 10, 8648–8664. [Google Scholar] [CrossRef]
- Li, X.; Corbett, A.L.; Taatizadeh, E.; Tasnim, N.; Little, J.P.; Garnis, C.; Daugaard, M.; Guns, E.; Hoorfar, M.; Li, I.T.S. Challenges and opportunities in exosome research—Perspectives from biology, engineering, and cancer therapy. APL Bioeng. 2019, 3, 11503. [Google Scholar] [CrossRef]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Agrawal, A.; Kyakulaga, A.-H.; Wilcher, S.A.; Gupta, R.C. Milk exosomes—Natural nanoparticles for siRNA delivery. Cancer Lett. 2019, 449, 186–195. [Google Scholar] [CrossRef]
- Segawa, K.; Suzuki, J.; Nagata, S. Constitutive exposure of phosphatidylserine on viable cells. Proc. Natl. Acad. Sci. USA 2011, 108, 19246–19251. [Google Scholar] [CrossRef]
- Long, K.B.; Beatty, G.L. Harnessing the antitumor potential of macrophages for cancer immunotherapy. OncoImmunology 2013, 2, e26860. [Google Scholar] [CrossRef]
- Park, S.-Y.; Kim, I.-S. Harnessing immune checkpoints in myeloid lineage cells for cancer immunotherapy. Cancer Lett. 2019, 452, 51–58. [Google Scholar] [CrossRef]
- Morse, M.A.; Garst, J.; Osada, T.; Khan, S.; Hobeika, A.; Clay, T.M.; Valente, N.; Shreeniwas, R.; Sutton, M.A.; Delcayre, A.; et al. A phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancer. J. Transl. Med. 2005, 3, 9. [Google Scholar] [CrossRef]
- Escudier, B.; Dorval, T.; Chaput, N.; André, F.; Caby, M.-P.; Novault, S.; Flament, C.; Leboulaire, C.; Borg, C.; Amigorena, S.; et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: Results of thefirst phase I clinical trial. J. Transl. Med. 2005, 3, 10. [Google Scholar] [CrossRef]
- Ohno, S.-I.; Takanashi, M.; Sudo, K.; Ueda, S.; Ishikawa, A.; Matsuyama, N.; Fujita, K.; Mizutani, T.; Ohgi, T.; Ochiya, T.; et al. Systemically Injected Exosomes Targeted to EGFR Deliver Antitumor MicroRNA to Breast Cancer Cells. Mol. Ther. 2013, 21, 185–191. [Google Scholar] [CrossRef]
- Banks, W.A.; Sharma, P.; Bullock, K.M.; Hansen, K.M.; Ludwig, N.; Whiteside, T.L. Transport of Extracellular Vesicles across the Blood-Brain Barrier: Brain Pharmacokinetics and Effects of Inflammation. Int. J. Mol. Sci. 2020, 21, 4407. [Google Scholar] [CrossRef]
- Yang, T.; Martin, P.; Fogarty, B.; Brown, A.; Schurman, K.; Phipps, R.; Yin, V.P.; Lockman, P.; Bai, S. Exosome Delivered Anticancer Drugs Across the Blood-Brain Barrier for Brain Cancer Therapy in Danio Rerio. Pharm. Res. 2015, 32, 2003–2014. [Google Scholar] [CrossRef]
- Tian, T.; Zhu, Y.-L.; Hu, F.-H.; Wang, Y.-Y.; Huang, N.-P.; Xiao, Z.-D. Dynamics of exosome internalization and trafficking. J. Cell. Physiol. 2013, 228, 1487–1495. [Google Scholar] [CrossRef]
- Heidarzadeh, M.; Gürsoy-Özdemir, Y.; Kaya, M.; Abriz, A.E.; Zarebkohan, A.; Rahbarghazi, R.; Sokullu, E. Exosomal delivery of therapeutic modulators through the blood–brain barrier; promise and pitfalls. Cell Biosci. 2021, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Toth, A.E.; Holst, M.R.; Nielsen, M.S. Vesicular Transport Machinery in Brain Endothelial Cells: What We Know and What We Do not. Curr. Pharm. Des. 2020, 26, 1405–1416. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, M.J.; Vandendriessche, C.; Vandenbroucke, R.E. Special delEVery: Extracellular Vesicles as Promising Delivery Platform to the Brain. Biomedicine 2021, 9, 1734. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Liu, L.; Ma, F.; Wong, C.W.; Guo, X.E.; Chacko, J.V.; Farhoodi, H.; Zhang, S.; Zimak, J.; Ségaliny, A.; et al. Elucidation of Exosome Migration Across the Blood–Brain Barrier Model In Vitro. Cell. Mol. Bioeng. 2016, 9, 509–529. [Google Scholar] [CrossRef]
- Qu, M.; Lin, Q.; Huang, L.; Fu, Y.; Wang, L.; He, S.; Fu, Y.; Yang, S.; Zhang, S.; Zhang, L.; et al. Dopamine-loaded blood exosomes targeted to brain for better treatment of Parkinson’s disease. J. Controll. Release 2018, 287, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, H.; Tachikawa, M.; Yagi, Y.; Umetsu, M.; Nurdin, A.; Miyauchi, E.; Watanabe, M.; Uchida, Y.; Terasaki, T. Cluster of Differentiation 46 Is the Major Receptor in Human Blood–Brain Barrier Endothelial Cells for Uptake of Exosomes Derived from Brain-Metastatic Melanoma Cells (SK-Mel-28). Mol. Pharm. 2018, 16, 292–304. [Google Scholar] [CrossRef]
- Sarin, H.; Kanevsky, A.S.; Wu, H.; Sousa, A.A.; Wilson, C.M.; Aronova, M.A.; Griffiths, G.L.; Leapman, R.D.; Vo, H.Q. Physiologic Upper Limit of Pore Size in the Blood-Tumor Barrier of Malignant Solid Tumors. J. Transl. Med. 2009, 7, 51. [Google Scholar] [CrossRef]
- Hobbs, S.K.; Monsky, W.L.; Yuan, F.; Roberts, W.G.; Griffith, L.; Torchilin, V.P.; Jain, R.K. Regulation of transport pathways in tumor vessels: Role of tumor type and microenvironment. Proc. Natl. Acad. Sci. USA 1998, 95, 4607–4612. [Google Scholar] [CrossRef]
- Akuma, P.; Okagu, O.D.; Udenigwe, C.C. Naturally Occurring Exosome Vesicles as Potential Delivery Vehicle for Bioactive Compounds. Front. Sustain. Food Syst. 2019, 3, 3. [Google Scholar] [CrossRef]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and Challenges of Liposome Assisted Drug Delivery. Front. Pharmacol. 2015, 6, 286. [Google Scholar] [CrossRef]
- Antimisiaris, S.G.; Mourtas, S.; Marazioti, A. Exosomes and Exosome-Inspired Vesicles for Targeted Drug Delivery. Pharmaceutics 2018, 10, 218. [Google Scholar] [CrossRef]
- Hood, J.L.; Pan, H.; Lanza, G.M.; Wickline, S.A.; Consortium for Translational Research in Advanced Imaging and Nanomedicine (C-TRAIN). Paracrine induction of endothelium by tumor exosomes. Lab. Investig. 2009, 89, 1317–1328. [Google Scholar] [CrossRef]
- Hood, J.L.; Scott, M.J.; Wickline, S.A. Maximizing exosome colloidal stability following electroporation. Anal. Biochem. 2014, 448, 41–49. [Google Scholar] [CrossRef]
- Qi, X.; Zhao, W.; Zhuang, S. Comparative study of the in vitro and in vivo characteristics of cationic and neutral liposomes. Int. J. Nanomed. 2011, 6, 3087–3098. [Google Scholar] [CrossRef]
- Gardiner, C.; Di Vizio, D.; Sahoo, S.; Théry, C.; Witwer, K.W.; Wauben, M.; Hill, A.F. Techniques used for the isolation and characterization of extracellular vesicles: Results of a worldwide survey. J. Extracell. Vesicles 2016, 5, 32945. [Google Scholar] [CrossRef]
- Wu, M.; Ouyang, Y.; Wang, Z.; Zhang, R.; Huang, P.-H.; Chen, C.; Li, H.; Li, P.; Quinn, D.; Dao, M.; et al. Isolation of exosomes from whole blood by integrating acoustics and microfluidics. Proc. Natl. Acad. Sci. USA 2017, 114, 10584–10589. [Google Scholar] [CrossRef]
- Maroto, R.; Zhao, Y.; Jamaluddin, M.; Popov, V.L.; Wang, H.; Kalubowilage, M.; Zhang, Y.; Luisi, J.; Sun, H.; Culbertson, C.T.; et al. Effects of storage temperature on airway exosome integrity for diagnostic and functional analyses. J. Extracell. Vesicles 2017, 6, 1359478. [Google Scholar] [CrossRef]
- Kalra, H.; Adda, C.G.; Liem, M.; Ang, C.-S.; Mechler, A.; Simpson, R.J.; Hulett, M.D.; Mathivanan, S. Comparative proteomics evaluation of plasma exosome isolation techniques and assessment of the stability of exosomes in normal human blood plasma. Proteomics 2013, 13, 3354–3364. [Google Scholar] [CrossRef]
- Mendt, M.; Kamerkar, S.; Sugimoto, H.; McAndrews, K.M.; Wu, C.-C.; Gagea, M.; Yang, S.; Blanko, E.V.R.; Peng, Q.; Ma, X.; et al. Generation and testing of clinical-grade exosomes for pancreatic cancer. JCI Insight 2018, 3, e99263. [Google Scholar] [CrossRef]
- Butreddy, A.; Kommineni, N.; Dudhipala, N. Exosomes as Naturally Occurring Vehicles for Delivery of Biopharmaceuticals: Insights from Drug Delivery to Clinical Perspectives. Nanomaterials 2021, 11, 1481. [Google Scholar] [CrossRef]
- Teng, F.; Fussenegger, M. Shedding Light on Extracellular Vesicle Biogenesis and Bioengineering. Adv. Sci. 2021, 8, 2003505. [Google Scholar] [CrossRef]
- Wiklander, O.P.B.; Brennan, M.Á.; Lötvall, J.; Breakefield, X.O.; EL Andaloussi, S. Advances in therapeutic applications of extracellular vesicles. Sci. Transl. Med. 2019, 11, 492. [Google Scholar] [CrossRef]
- Harn, H.-J.; Chen, Y.-S.; Lin, E.-Y.; Chiou, T.-W. Exosomes in clinical trial and their production in compliance with good manufacturing practice. Ci Ji Yi Xue Za Zhi 2020, 32, 113–120. [Google Scholar] [CrossRef]
- Rezakhani, L.; Kelishadrokhi, A.F.; Soleimanizadeh, A.; Rahmati, S. Mesenchymal stem cell (MSC)-derived exosomes as a cell-free therapy for patients Infected with COVID-19: Real opportunities and range of promises. Chem. Phys. Lipids 2021, 234, 105009. [Google Scholar] [CrossRef]
- Rome, S. Biological properties of plant-derived extracellular vesicles. Food Funct. 2019, 10, 529–538. [Google Scholar] [CrossRef]
- Zhang, M.; Viennois, E.; Prasad, M.; Zhang, Y.; Wang, L.; Zhang, Z.; Han, M.K.; Xiao, B.; Xu, C.; Srinivasan, S.; et al. Edible ginger-derived nanoparticles: A novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials 2016, 101, 321–340. [Google Scholar] [CrossRef]
- Xiao, X.-B.; Gu, Y.; Sun, D.-L.; Ding, L.-Y.; Yuan, X.-G.; Jiang, H.-W.; Wu, Z.-X. Effect of rituximab combined with chemotherapy on the expression of serum exosome miR-451a in patients with diffuse large b-cell lymphoma. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1620–1625. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).