Abstract
Herbivore-induced plant organic volatiles (HIPVs) have recently been studied to improve biological pest control. In contrast, the effects of volatile organic compounds (VOCs) that are not induced by herbivory (non-HIPVs) have received less attention. The latter are essential in the first stages of crop colonization by entomophagous insects (predators and parasitoids) used in biological pest control programs. Furthermore, the effects on entomophagous insects of different cultivars of a cultivated botanical species have not been studied. The aim of this work was to study the different non-HIPVs found in 10 tomato cultivars used in tomato greenhouses on two entomophages: the egg parasitoid Trichogramma achaeae (Hymenoptera, Trichogrammatidae) and the zoo-phytophagous predator Nesidiocoris tenuis (Hemiptera, Miridae). The results indicate that although there is considerable quantitative and qualitative variation in the emission of VOCs in the 10 tomato cultivars analysed, this variability made it difficult to determine the influence of the volatiles on the attraction of the predatory species N. tenuis, with only one cultivar (Rebelion) exhibiting a significantly higher attractiveness than the rest of the cultivars. For the parasitoid T. achaeae, these same volatiles had a significant effect (in part) on parasitoid behaviour. However, this attraction was not reflected in the discriminant analysis, at least for the volatiles analysed. The analysis showed four groups of well-differentiated cultivars, according to the non-HIPV composition, and this bore no relation to the levels of attractiveness registered in the different cultivars, with the exception again of the Rebelion cultivar, which seems not to be very attractive for the parasitoid and its parasitism activity. The implications of non-herbivore-induced (non-HPV) VOCs in the biological control of greenhouse pest species are described and discussed.
1. Introduction
Entomophagous responses can be directly influenced, either positively or negatively, by various characteristics of the host plants of their phytophagous pests [1]. Especially important are the abundant studies documenting the tritrophic effects of host plant chemistry on the host location and acceptance behaviour of the natural enemies (in particular, parasitoids) of insect herbivores, including induced volatiles released after herbivore damage [2]. The effects of plant host chemical compounds are usually positive from the point of view that they tend to result in an increase in the parasitoid’s capabilities for greater success and efficiency in finding the host insect and parasitizing it [3]. Moreover, the chemical components of the host plant can also affect the behaviour and success of the parasitoid species by different pathways (e.g., increased mortalities, number of offspring, body size and adult fertilities and fecundities). Side effects on the behaviour of insect parasitoids can be direct (e.g., when the immature stages of the parasitoid meet toxins or metabolites in the haemocoel of their phytophagous host) or indirect (e.g., when the behaviour of female parasitoids is affected by the size and/or quality of the phytophagous host). In contrast to the relationships of plant chemical constituents and host insect foraging, locating and acceptance behaviour, a small (though growing) amount of the literature has been devoted to elucidating the processes by which plant chemistry negatively affects parasitoid biology [2].
The volatile compounds released from plants can be divided into constitutive compounds, which are continuously present in the plant, and inducible compounds, which are released when structures are damaged after pathogen or herbivore attacks [4,5]. In addition, plants have evolved mechanisms that recruit herbivores’ natural enemies, and this attraction is often mediated by induced volatile compounds that function as prey-associated signals for experienced foraging carnivores. This phenomenon is referred to as indirect defence [1,6,7].
Although most plants release constitutive volatile compounds, the composition and quantity of the emitted blend vary greatly in response to biotic stresses, especially after herbivore and pathogen attack [4,6]; they also vary between varieties within plant species [8,9,10,11]. These differences between varieties may result in differences in their attractiveness to herbivores’ natural enemies [10,11]. However, little to nothing is known about the consequences of these differences between plant genotypes in terms of the effectiveness of natural enemies under field or greenhouse conditions [12].
Tomato cultivars, Lycopersicum esculentum, and their wild ancestors of the genus produce methyl ketones and acyl sugars, which are present in trichomes, which play an important role in resistance to some insect pests and/or mites [13,14,15].
To comprehend how entomophagous species face potentially toxic host plant chemistry (antibiosis and/or antixenosis effects), it is critical to assess whether the effects of host plant chemistry on entomophages are indirect or direct [2]. Thus, the manipulation of plant signals offers very promising prospects for increasing the effectiveness of biological control agents (predators and parasitoids) under field conditions [16]. Although chemical signals have never been part of plant breeding criteria, it is known that cultivars differ in their production of natural volatiles that attract predatory and parasitoid species [12]. Therefore, instead of this selection being accidental, varieties should be improved for their ability to attract entomophagous species [16]. An important objective in research on tritrophic relationships, when behaviour is applied to the control of arthropod pest species (insects and mites), is to demonstrate whether it is possible to obtain and subsequently use cultivars, on the one hand, with resistance to their arthropod pest species and, on the other hand, at the same time increase the effectiveness of the biological control agents (predators and parasitoids species) that control those pest species [17]. However, it should not be forgotten that the genetic variability in any plant trait that may affect biological control agents is likely to be fortuitous (especially in Classical Breeding) [17,18]. Applied systems, therefore, are primarily valuable in showing how pre-existing host plant variation may affect phytophagous–entomophagous interactions [17].
In Spanish tomato greenhouses, the egg parasitoid Trichogramma achaeae (Hym.: Trichogrammatidae) and the predator Nesidiocoris tenuis (Hem.: Miridae) are the two species most used in biological control, the first one for Bemisia tabaci (Hem.: Aleyrodidae) and the second one for Tuta absoluta (Lep.: Gelechiidae) [19].
However, earlier studies found that the tomato cultivar (e.g., [20]) can affect both entomophagous insect species. Thus, this work had multiple objectives: firstly, to study the adult behaviour of the two entomophages, N. tenuis and T. achaeae, in relation to the non-HIPV VOCs released from ten tomato cultivars; secondly, to study the chemical composition of these VOCs and their main components; and finally, to study the effects of the cultivars on the population dynamics of both entomophagous species under semi-field conditions, based on the results obtained.
2. Materials and Methods
2.1. Plants
Ten L. esculentum varieties were used in this study, encompassing the range of tomato types most utilized in the greenhouse horticultural industry in south-west Spain (Table 1). The seeds were germinated in a seedbed and, reaching 13 BBCH, approximately 15–20 cm in height and 3 leaves, were individually transferred to 250 mL pots containing coco fibre. They were maintained under these conditions until carrying out the trials.

Table 1.
Origins of tomato cultivars used in the trials.
2.2. Insects
The N. tenuis predator specimens used in the trials were bought from a local supplier (Agrobio S.L., La Mojonera, Almeria, Spain). They were reared under laboratory conditions for two generations before being used in the laboratory trials. For this purpose, plastic containers (20 L) containing dispersion material were used, and the alternative prey, E. kuehniella, was used as the food source.
The parasitoid, T. achaeae, came from populations collected in the Canary Islands. They were reared under laboratory conditions using E. kuehniella eggs as the alternative host and a honey drop as a food source, following the method developed by Cabello [21].
In all cases, the environmental conditions for growing the plants, rearing the insects and the bioassays were as follows: 25 ± 1 °C, 60–80% H.R. and 16:8 h of a light–dark photoperiod under homogeneous light conditions.
2.3. Olfactory Response of T. achaeae and N. tenuis to Different Tomato Cultivars
To choose the most effective olfactometer for each insect species, three different olfactometer types were evaluated beforehand using E. kuehniella eggs and their scales as an attractive odour source for the predator and the parasitoid. Prior to this, moving-air bioassays were conducted with a linear tube, Y-tube and a four-arm olfactometer [22,23] as well as a 9-choice still-air olfactometer. The most suitable olfactometer for the T. achaeae evaluation (in terms of measuring the greatest number of insects trapped in each device) was the 9-choice still-air olfactometer. For the N. tenuis bioassays, the four-arm moving-air olfactometer proved the most appropriate.
The 9-choice still-air olfactometer was the same device used for N. pseudoferus (Hem.: Nabidae) adults, previously changed from Prokopy et al. [24] and Van Tol et al. [25,26]. The design was the same with different dimensions, as follows. The olfactometer consisted of a square aluminium frame (length: 240 mm; width: 240 mm; height: 40 mm) and closed below and above with two plastic plates (as decision chamber). The upper plate had nine holes (diameter 10 mm), evenly distributed. These holes had a cylinder (length: 35 mm and of the same diameter) attached to them, which led into a plastic container (volume 100 mL) (as trapping device), which in turn was separated from another container (as odour source) but separated with a metal mesh.
The four-arm moving air olfactometer was the same as that described by Pettersson [27] and adapted by Vet et al. [23]. It consisted of a four-arm olfactometer chamber (length: 300 mm; width: 300 mm; height: 20 mm) constructed from a transparent methacrylate lid. Air was drawn into the system through the chamber’s central outlet using a vacuum pump. It then passed through the following elements: firstly, through a 30 mL chamber filled with charcoal, then through a 250 mL humidifying reservoir and finally a 1500 mL scent bottle (the treatment chamber). Each airflow channel was accurately adjusted using precision flowmeters situated just before the entrance to the decision chamber. The flowrate was regulated at 50–60 mL/min per channel.
2.4. Bioassay Procedure—T. achaeae
The experimental design was completely randomized, having only a ‘treatment’ (tomato cultivar) factor, with 3 replications performed per trial. All the trials were carried out over 24 h in climatic cabinets under homogeneous light conditions (Memmert®, Memmert GmbH + Co. KG, ICP 600, Schwabach, Germany) (25 °C ± 1, RH: 75–85%, 16:8 h L:D photoperiod).
The T. achaeae tomato cultivar preference was evaluated in 7 different varieties at the same time. A leaflet from a completely formed leaf was used an odour source. Each petiole was submerged and sealed with Parafilm® inside an Eppendorf® tube (1.5 mL), which was filled with tap water and placed on each hole. An empty hole of each olfactometer acted as a control. Approximately 150 T. achaeae adults were released in the centre of each olfactometer and given access to E. kuehniella eggs. After 24 h, the number of T. achaeae adults captured in each trapping device was counted. This method had already been evaluated in an earlier trial [28].
2.5. Bioassays Procedure—N. tenuis
The experimental design was completely randomized, having only a ‘treatment’ (tomato cultivar) factor, with 3 replications performed per trial. A total of 3 trials were carried out. Twenty N. tenuis adult females (less than 72 h old) were released at the central point of the 4-arm olfactometer. These females had previously been isolated and starved, with only a moisturized sponge as a water source, 24 trials.
As an odour source, a complete seedling (height 15–20 cm) was placed inside the odour chamber. Four different varieties were tested at the same time using the Vernal cultivar as the control; this cultivar was chosen because it was shown to have a good capacity for N. tenuis attraction and nymphal development in previous works [29]. The olfactometer exposure chamber was divided into a central area (the release point) and 4 others, each for one of the four odour sources. After 20 min of continuous airflow, the number of insects found in each of the four areas was recorded, and this measurement was taken as the final behavioural choice of the N. tenuis adult females. After three repetitions, the exposure chamber was cleaned with ethanol and rinsed with distilled water.
2.6. Volatile Collection
To identify the most common volatiles, namely those appearing at the highest concentrations in the tomato leaf [30,31,32], the chromatographic peaks were compared with 6 standards with a GC purity level. The following compounds were sought and analysed: carene, β-caryophyllene, carvacrol, eugenol α-phellandrene, α-humulene and α-Pinene (all provided by Sigma–Aldrich, Chemie GmbH, Mühldorf, Germany).
The samples were taken from small plants (cultivars that were transplanted with 30 days before from the seedbed and three leaves completely developed, approximately 15–25 cm in height) at the same time as the olfactometer trials were carried out. To conduct the volatile analysis, a complete seedling leaf was weighed and sequentially placed in a chromatography test tube. Three replicates were sampled with the different tomato varieties.
The compounds emitted from the leaves were analysed using the GC-MS mass spectrometry technique. The equipment used was a gas chromatograph (Bruker 450-GC) coupled to a mass detector (Bruker 320-MS triple quadrupole mass spectrometer) (Bruker Inc., Fremont, CA, USA) equipped with an automatic self-sampler (CTC analytics, Zwingen, Switzerland) to perform the headspace injection technique. A total of 500 μL to each injection aliquot was injected onto a Rxi-5 Sil MS column (30 m × 0.25mm × 0.25 µm) (Restek, Bellefonte, PA, USA). The oven temperature was set at 65 °C for 10 min, increasing to 150 °C at a rate of 10 °C/min holding for 3 min, finally increasing to 280 °C at a rate of 20 °C/min holding for 3 min. The injector temperature was 60 °C for 1 min, increasing to 150 °C and 250 °C for 5 min and 3 min, respectively (a rate of 20 °C/min 200 °C/min), while the detector temperature was 250 °C. The ion source temperature was 250 °C. Selected ion monitoring modes were used for the qualitative measurements. Helium was used as the carrier gas at a 1 mL/min flowrate. The identification of the compounds was based on comparisons of their mass spectra with those recorded in the spectra library database.
2.7. Tomato Cultivar Effects on T. achaeae and N. tenuis Population Dynamics
The trial design was completely randomized with one factor: treatment (tomato cultivar) and 3 repetitions over time. The plants were grown for five weeks after transplanting.
The randomized trials were conducted in cages inside a climatic chamber (CCM-FP, SCLAB: 3.20 × 2.40 × 2.20 m3) (conditions set at 25 °C, 50% RH, L:D = 16:8) between April and July 2015, at the Tecnova Technological Center (Almeria, Spain). Ten cages (40 × 50 × 70 cm3) were constructed. These comprised a wood frame with a wood bottom, a Plexiglas® sheet on top and on two sides, and on two sides covered with a fine mesh (10 × 20 threads/cm). One side of the fine mesh was used as a door; the cage was closed with Velcro® strips.
Thirty days after sowing, six potted tomato plants from each cultivar (with a height of about 15 cm and three real leaves) were transferred to the cages. Watering and fertilization management followed local commercial practices.
All cages were inoculated on Day 1; this involved the releases of the N. tenuis adults at a ratio of 2 individuals per plant. T. achaeae were released in the cages five times a week at a ratio of 92 adults/plant in accordance with the commercial protocols [33].
Parasitoid activity was recorded weekly using the ‘sentinel eggs egg’ method. A total of 100 UV-irradiated E. kuehniella eggs were glued onto cardboard (4.3 × 2.8 cm) and placed in the centre of each plant using a pin to avoid predation by N. tenuis. After a week, the sentinel eggs were removed and incubated at 25 ± 1 °C and 60–80% RH, until hatching, at which time the T. achaeae parasitization was determined. We also determined possible “collapsed” eggs with and without contents that could be due to possible predation by N. tenuis.
To establish the N. tenuis population, a biweekly source of E. kuehniella eggs was supplied to each cage. About 200 frozen eggs glued with water onto cardboard (4.3 × 2.8 cm) were placed on the upper part of the central plant. After five weeks, the total N. tenuis population was assessed in each cage.
2.8. Statistical Analysis
To investigate whether parasitoid and predator olfactory preference differed between the various combinations of tomato cultivar treatments provided, the data were analysed using a generalized linear model (GZLM) with a Poisson distribution and a logit link function, with the tomato cultivar as the fixed factor. The response variable was the number of adult insects responding to one of the volatile sources.
The volatile profiles overall were analysed by using principal component analysis (PCA) following the log10 transformation of the data and subtracting the average value of all the treatments to normalize the data [34]. For the volatile emission patterns, the measured peak areas divided by the aboveground fresh mass of the plant were analysed through multivariate data analysis using projection to latent structures discriminant analysis (PLS-DA).
This projection method can determine whether samples belonging to each treatment group can be separated based on quantitative and qualitative differences in their volatile blends [35].
Individual volatiles were analysed for significant changes between cultivars using a GZLM with a binomial distribution and a logit link function, with the tomato cultivar as the fixed factor. A quasi-binomial distribution was fitted in the model due to overdispersion.
The percentages of parasitism by T. achaeae when in contact with the different tomato cultivar plants were analysed using a GZLM with a binomial distribution and a logit link function, with the tomato cultivar as fixed factor. In turn, the numerical values for the N. tenuis nymphs and adults per plant were analysed using GZLM Poisson distribution and a logit link function, with the tomato cultivar as the fixed factor.
All the analyses were performed using IBM SPSS Statistics 23.0 (IBM, Armonk, NY, USA) statistical software.
3. Results
3.1. Olfactory Response of T. achaeae and N. tenuis to Different Tomato Varieties
Ten cultivars of L. esculentum were compared for their attractiveness to the egg parasitoid T. achaeae (Table 2) and the zoo-phytophagous predator N. tenuis (Table 3) in two and three different trials, respectively.
Table 2.
The mean number (± SE) of T. achaeae adults attracted to different tomato cultivars in a 9-choice still-air olfactometer (* in each trial; values followed by the same letter do not show significant differences at p = 0.05).
Table 3.
The percentage (± SE) of N. tenuis adult females attracted to different tomato cultivars in a 4-arm olfactometer (* in each trial; values followed by the same letter do not show significant differences at p = 0.05).
Regarding the parasitoid, the results for individuals attracted to different odour sources are shown in Table 2. In the first trial, T. achaeae showed a significant greater preference for Montenegro (in the first group), a second group with Creativo and Vernal, a third including Bronco, Byelsa and Caniles and a minor group with Rebelion and the control. Statistically significant difference existed between each group (Omnibus test: Chi-square likelihood ratio = 83.626; df = 7; p < 0.01).
In the second egg parasitoid trial, three clear response ranges were found again (Table 2). The first group contained Genio, followed by a second group with Bronco, Byelsa and Vernal, while the last group (with a low level of attractiveness) had similar results to the control; this group included the Pasadena, Ramyle and Rebelion cultivars (Omnibus test: Chi-square likelihood ratio = 114.498; df = 7; p < 0.01).
In the first trial (Table 3) for the zoo-phytophagous predator N. tenuis, no significant differences were observed when comparing the cultivars Bronco, Byelsa, Ramyle and Vernal in a four-arm olfactometer (Omnibus test: Chi-square likelihood ratio = 0.941; df = 3; p = 0.815). The attraction of adult females was similar in each cultivar, with only Byelsa showing slightly less attraction, but this was not significant.
In the second trial (Table 3), N. tenuis exhibited a significantly higher attraction to Rebelion than to the other cultivars evaluated (Omnibus test: Chi-square likelihood ratio = 10.102; df = 3; p = 0.018). The attraction to Rebelion was more than twice that found in Montenegro (Chi-square = 4.800; df = 1; p = 0.028) with N. tenuis showing equal distribution between them.
In the third trial (Table 3), N. tenuis exhibited no behavioural preference when presented a choice between the odour of the Creativo, Genio, Pasadena and Vernal tomato cultivars (Omnibus test: Chi-square likelihood ratio = 1.211; df = 3; p = 0.750).
3.2. Volatile Collection
To address the question of whether the blend of volatiles might mediate the differences in the attractiveness of the predator or parasitoid in tomato cultivar choice, the following analyses were performed.
In the first analysis of the total values of the six compounds emitted by the leaves of the 10 tomato cultivars (Table 4), the significant effect is the same (Omnibus test: Chi-square likelihood ratio = 56.584; df = 0, p < 0.01). The value found was significantly higher in the cultivar Rebelion, with no significant differences with the next one, corresponding to the cultivar Vernal.
Table 4.
The total values (kcount ±SE) of 6 chemical compounds emitted by leaves, according to each of the 10 tomato cultivars studied. (* Values followed by the same letter do not show significant differences at p = 0.05).
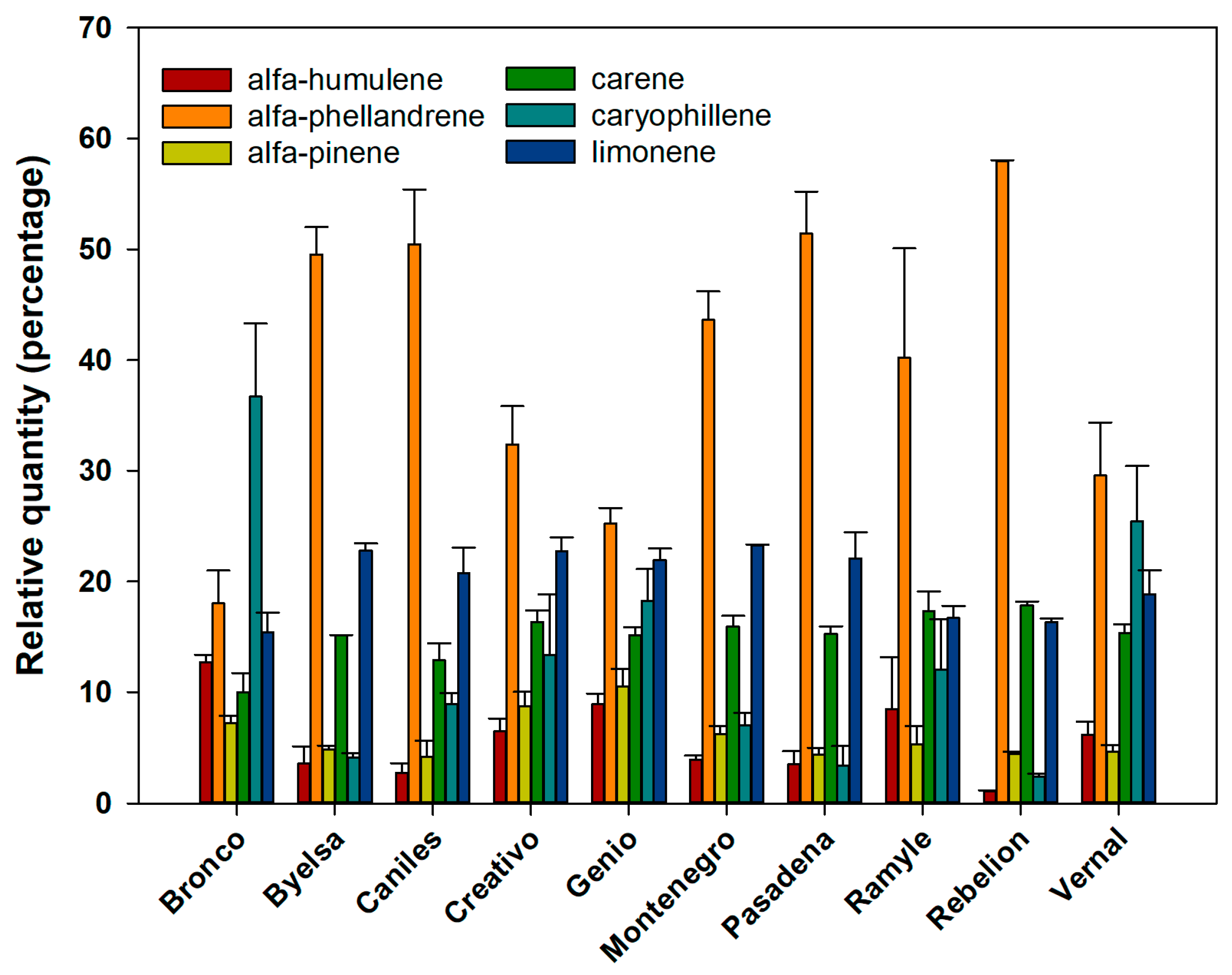
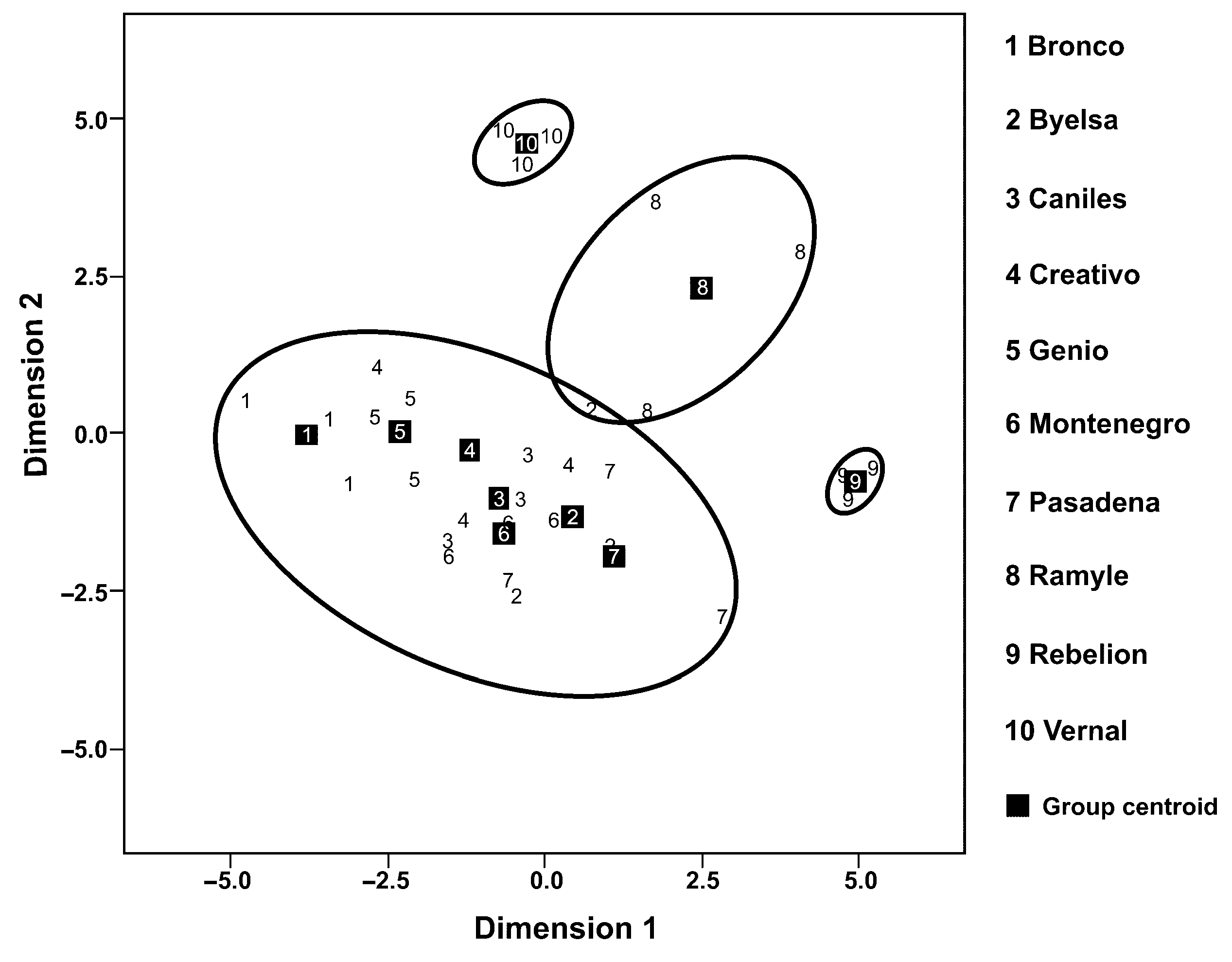
In a second analysis of the percentage composition of the six volatiles analysed, all the cultivars showed significant differences in the volatiles’ emission rates (Figure 1). However, the discriminant analysis (Figure 2), based on the composition of the mixture of volatiles analysed, made it possible to discern four well-defined cultivar groups (the percentage of variance explained by Dimension 1 = 46.2% and by Dimension 2 = 31.1%; accumulated % = 77.3%) (Canonical correlation: Dimension 1 = 0.943; Dimension 2 = 0.919; Wilks’ Lambda: Dimension 1: Wilks’ Lambda = 0.001, Chi-square = 137.242, df = 54, p < 0.001; Dimension 2: Wilks’ Lambda = 0.013, Chi-square = 90.904, df = 40, p < 0.001). The first group includes Bronco, Byelsa, Caniles, Creativo, Genio and Montenegro, well differentiated from the other three single cultivar groups (Ramile, Rebelion and Vernal). For the canonical discriminant function coefficients, we obtained for Dimensions 1 and 2 the following values:
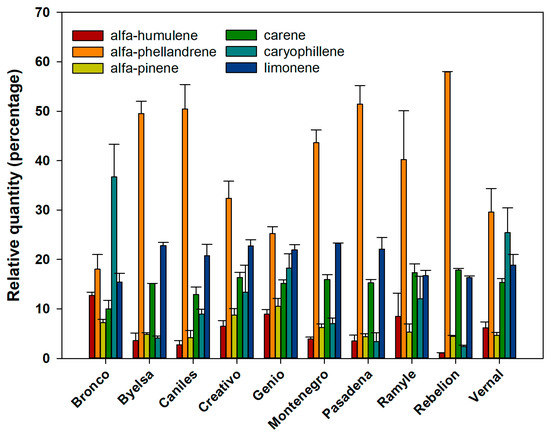
Figure 1.
The relative values (percentage ± SE) of each of the 6 chemical compounds emitted by the leaves, according to each of the 10 tomato cultivars studied.
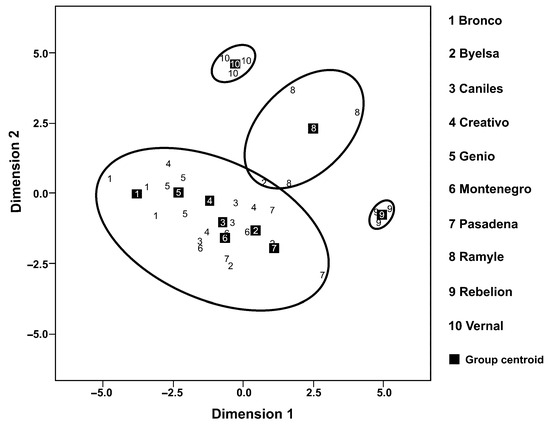
Figure 2.
A principal component analysis (PCA) of the volatiles presents in ten tomato cultivars based on the principal component values (Dimension 1 and Dimension 2) (numbers in squares: midpoint coordinates for each cultivar).
Dimension 1 = 0.005 + 1.274 × alfa-humulene + 8.249 × alfa-phellandrene + 6.937 × alfa-pinene + 8.051 × carene – 2.410 × caryophyllene – 18.932 × limonene.
Dimension 2 = 0.062 + 0.670 × alfa-humulene – 8.619 × alfa-phellandrene + 1.932 × alfa-pinene + 8.866 × carene + 2.121 × caryophyllene + 0.597 × limonene.
3.3. Tomato Cultivar Effects on T. achaeae and N. tenuis Population Dynamic
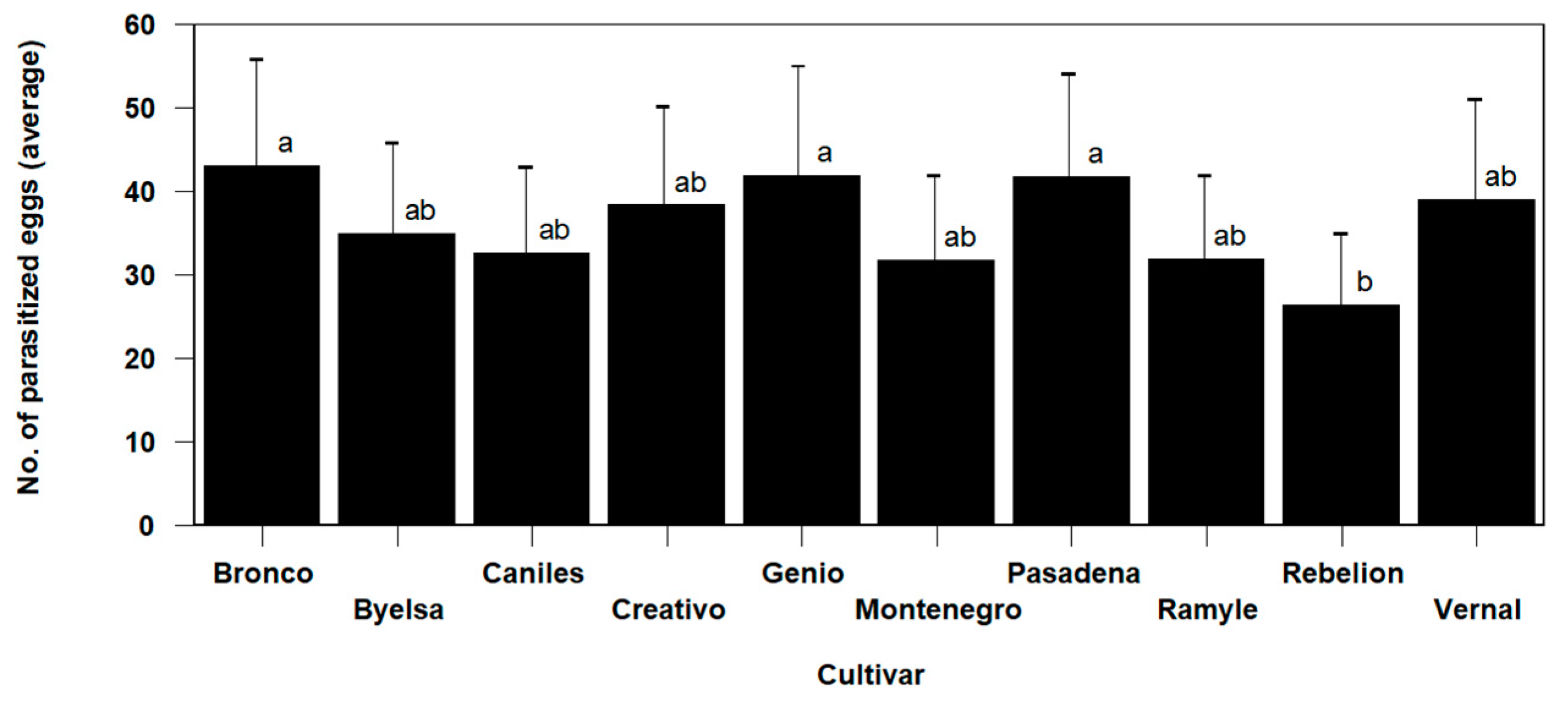
The statistical analysis of the data shows a significant cultivar effect on the parasitization activity of T. achaeae (Omnibus test: Chi-square likelihood ratio = 106.834, df = 12, p < 0.01). The mean number of eggs parasitized by T. achaeae was between the maximum value reached (42.90 ± 4.54% parasitized eggs) in the Bronco cultivar and the 26.22 ± 4.54% value in the cultivar with the lowest levels (Rebelion in this case), highlighting significant differences between this cultivar and the Genio, Bronco and Pasadena cultivars (Figure 3). No differences were found among the other cultivars, with all of them showing a higher average parasitism level.
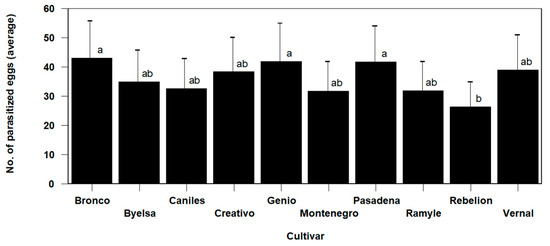
Figure 3.
Mean values (±SE) per plant for the number of Ephestia kuehniella eggs parasitized by Trichogramma achaeae, according to cultivar, in the trial conducted in cages under laboratory conditions (values followed by the same letter do not present significant differences at p = 0.05).
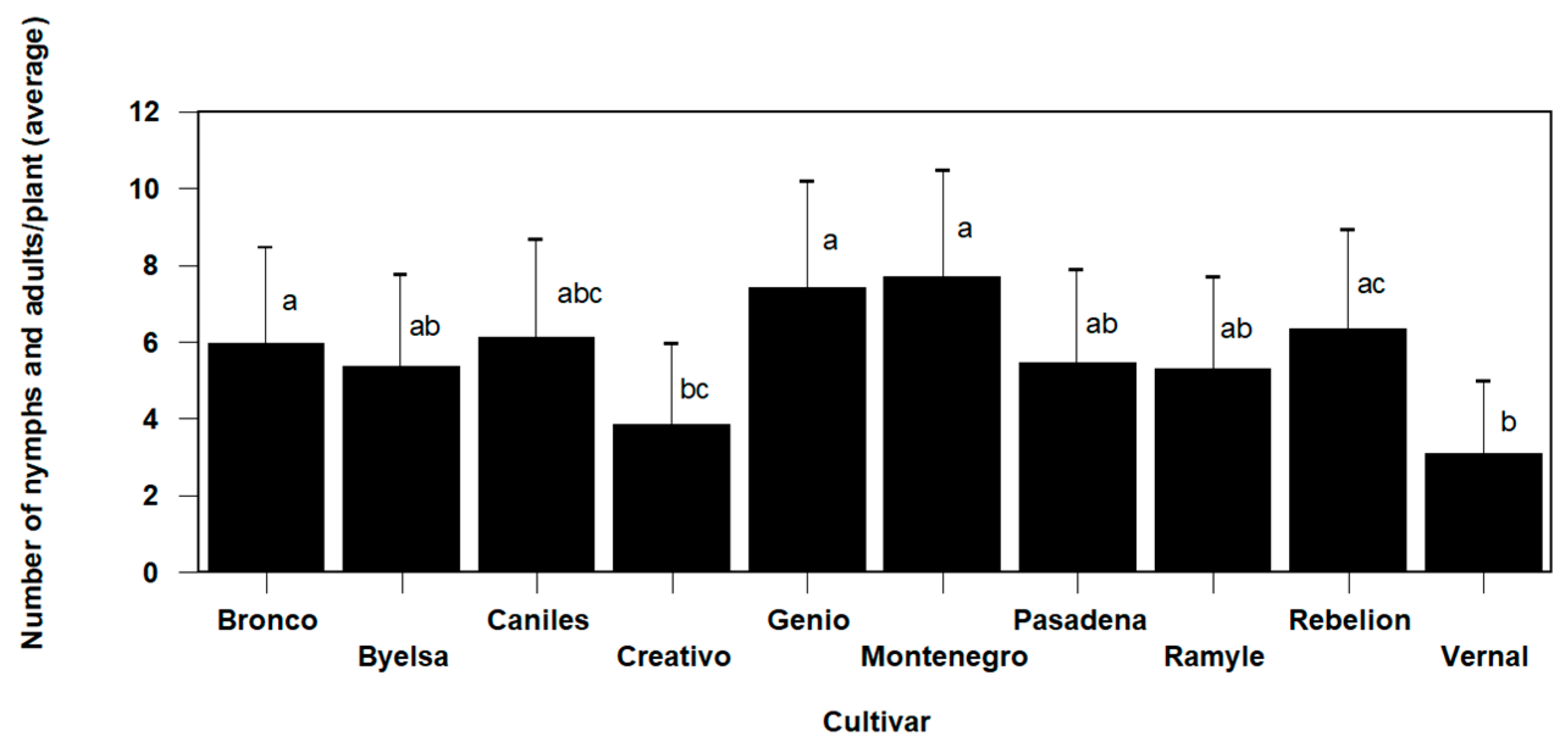
N. tenuis populations were significantly different after five weeks according to the cultivar (Omnibus test: Chi-square likelihood ratio = 20.395, df = 9, p = 0.016). The Montenegro (7.7 ± 1.21 predators/plant) and Genio (7.39 ± 1.19 predators/plant) cultivars had a higher total population of nymphs and adults, showing statistically significant differences compared to the variety with the lowest recorded population levels (3.06 ± 0.76 predators/plant), in this case the Vernal cultivar (Figure 4). The other varieties presented intermediate values between these three varieties.
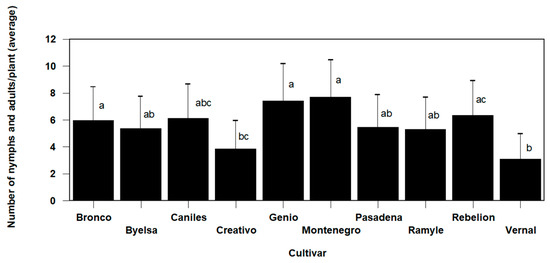
Figure 4.
The mean values (±SE) per plant of the number of Nesidiocoris tenuis nymphs and adults according to the cultivar, in the trial conducted in cages under laboratory conditions (values followed by the same letter do not present significant differences at p = 0.05).
4. Discussion
Due to the process of domestication and breeding, the tomato has great genotypic variability generated amongst its diverse types and commercial cultivars. It is characterized by having shared ancestral variations, a historical gene flow with closely related species and the introgression of resistance genes (R genes) from wild species [35]. Hence, there is great variability across the spectrum and in the level of arthropod resistance among accessions of this species. Arthropod resistance has been associated with a set of characteristics, including the physical and chemical properties of glandular trichomes and chemical defences expressed constitutively and when injuries or wounds associated with leaf lamellae occur [36]. These defences affect both pest species and natural enemies that feed on them as they share the same habitat.
During host plant choice by insects, orientation and landing are performed using visual and olfactory stimuli, the latter playing a predominant role [37]. In entomophages, information from the plant is important in searching for host or prey. A sequence of responses to various sources of information brings the predator/parasitoid closer and closer to its potential victim. Chemical information from plant volatiles plays a key role in the foraging behaviour, especially over long distances, while the importance of herbivore-induced signals increases as the distance to the victim decreases [18]. Volatiles in plants are biosynthesized in specialized tissues and stored in sub-dermal storage cavities; these secretory cavities are in the glandular trichomes of plants such as Solanaceae [38,39]. However, tomato domestication appears to have reduced the attraction of herbivore predatory attraction to HIPVs [40].
Once natural enemies are in contact with the plant, they will be affected by the plant’s own constitutive defences. Thus, in tomato, we find two types of constitutive defences, glandular and non-glandular trichomes, both acting physically and chemically by exuding toxic and sticky compounds against arthropods and defences based on the production of toxic and growth-inhibiting chemicals [39,41]; this is of great importance for the predator N. tenuis due to its zoo-phytophagous regime.
In this study, it was found that although there was considerable variation in the emission of constituent volatiles in the 10 tomato cultivars analysed (Figure 1 and Figure 2), this variability did not influence their attractiveness to the zoo-phytophagous N. tenuis. Only one cultivar (Rebelion) exhibited significantly greater attractiveness than the other varieties.
For the egg parasitoid, T. achaeae, constitutively emitted volatiles (in the 10 cultivars evaluated) did influence parasitoid behaviour. However, this attractiveness was not reflected in the discriminant analysis (Figure 1), at least for these volatiles, since the analysis showed four well-differentiated groups of varieties which were not related to the different levels of attractiveness we recorded. This may be modulated by the significantly higher total amount of volatiles emitted in the cultivars Rebelion and Vernal (Table 4).
Once in contact with the plant, each variety affected the parasitoid and predator populations differently, both in terms of the number of E. kuehniella eggs parasitized by T. achaeae and the total number of N. tenuis we found on them. This suggests that differences in the constitutive defences of the tomato varieties will affect the two natural enemies differently when they encounter the plant.
Initially, one would not expect information from the plant to be adaptive for generalist parasitoids such as Trichogramma spp., since they attack a wide range of hosts. At the plant level, they would be expected to respond more to plant-derived volatiles than to species-specific ones [42]. However, as in our case with T. achaeae, it has been reported in studies using airflow olfactometers [22,42] that they and other Trichogramma spp. respond to tomato constituent volatiles [43,44] as well as to other volatiles derived from fresh plant parts and plant extracts.
Although the response of Trichogramma spp. to plant volatile info chemicals seems to be a common phenomenon [42,45], this response is not always positive towards plant constituent volatiles; for example, Romeis et al. [46] and Bar et al. [47] reported T. chilonis repellence to pigeon pea pods and T. semifumatum repellence to cotton, respectively. Other authors have documented attraction and/or repellence depending on the part of the plant studied—Cabello and Vargas [22] found that T. sp. nr. buesi exhibited repellence to cotton leaves but not to the flowers.
In the case of the varieties we studied, the olfactory response of T. achaeae varied between 2.0 and 26.0 for the most and least attractive cultivars, respectively (Table 2). This range is greater than that reported by Tandon et al. [48] (23–40%) for T. chilonis, although their study looked at other tomato varieties and used a Y-olfactometer. It has been suggested that the variability found in volatile mixtures may be because the host plant genotype informs the foraging predators and parasitoids about the identity and quality of the host plant and the prey, but the utility of innate and immutable responses to volatile mixtures may be limited, thus favouring learning instead [49]. In contrast, Bjorksten et al. [50] suggested that the learning of the tomato stimuli in Trichogramma is not always necessary for successful “host foraging”.
Once Trichogramma has selected the host plant, the plant’s own constitutive herbivory defences act on the parasitoid. In our study, we found significant variation between varieties, with Rebelion having the lowest value (26.22 ± 4.54 parasitized eggs/plant), while Bronco had the highest (42.90 ± 5.82 parasitized eggs/plant) (Figure 3). These differences may be due to the tomato’s constitutive defences against arthropods, which were expressed to varying degrees depending on the genotypes studied. These defences include gland-like trichomes that physically impede movement or trap and (in some cases) intoxicate arthropods on the plant surface. They can also be affected by constitutively expressed defences associated with leaf lamellae that function as growth inhibitors or toxins [41]. These effects range from reduced host or prey foraging efficiency (as the predator or parasitoid movement physically interfered directly) to high mortalities caused by complete entrapment or content lethal intoxications when in contact with sticky trichomes or from toxic contents discharged into such trichomes [41]. In the absence of further studies, it is unclear whether the density and levels of various toxicants found in trichomes might affect T. achaeae parasitism. Thus, Kauffman and Kennedy [51] found reductions in the percentage of the parasitism of Manduca sexta (Lep.: Sphingidae) by T. pretiosum and T. exiguum as a function of trichome density and composition depending on the tomato genotype.
In the case of N. tenuis, the detection and preference implications of different volatile bouquets are twofold due to the insect’s zoo-phytophagous regime. Firstly, they can be used for habitat location and, secondly, as an alternative food source in the case of prey scarcity. However, Lins et al. [36] found that N. tenuis has no preference for healthy tomato plants when faced with a draught (the absence of scent), and prior experience is necessary for it to show a predilection for healthy tomato plants. In our case, this prior experience did not occur, as the only plant material to which they had been previously exposed were bean pods, which were used as an oviposition substrate during bedbug breeding. This learning requirement for N. tenuis selection may partly explain the results obtained, namely no preference for any cultivar, considering the variability in the volatile amount and composition between varieties. However, the differences in Rebelion and Vernal with respect to the other varieties show that this cultivar has a certain characteristic in its volatile composition that triggers an innate response in N. tenuis females (Figure 1 and Figure 2). On the other hand, Naselli et al. [52] did find a positive response in N. tenuis females towards healthy tomato plants, while Rim et al. [53] found the same response towards aubergine and sesame plants. This seems not only to be a learning process but also to be influenced by the constitutive composition mix of the tomato cultivar. As with the two previous references [36,52], the trials were conducted on different tomato varieties, Moneymaker and Marmande, respectively.
Being in contact with the different varieties and with a low level of food input (E. kuehniella eggs) induced phytophagy and thus detected a more marked effect on the N. tenuis populations; they exhibited a different level of development depending on the cultivar. This is also expected considering the initial nymphal stage development feeding on the host plant [53]. The tomato cultivar has previously been described as an influential factor in the development of N. tenuis populations. Cabello et al. [29] found a significant effect on the development of N. tenuis populations among several types of tomato varieties in field trials against T. absoluta.
The differences in tomato varieties should be considered an important factor in biological control programs when they involve the use of N. tenuis and T. achaeae. To date, this aspect has hardly been studied (Sanchez et al. [20] being an exception). Nonetheless, our study demonstrated that it is an important aspect to consider in future breeding and cultivar improvement programs given the increase in the number of commercial varieties associated with the wide range of fruit cultivar types on the market. Indeed, tomato plant chemistry has been shown to have a marked effect on natural enemy–host plant interactions. Most studies on tritrophic relationships have examined the influence of plant chemistry on host plant foraging and the acceptance behaviour of natural enemies in contact with their herbivores’ prey or host. In contrast, relatively few studies have specifically identified the negative tritrophic effects of plant chemistry on the fitness of natural enemies, and far fewer have investigated the mechanisms by which plant chemistry affects natural enemies [2]. Moreover, in certain cases, bioriented breeding programs have eliminated or weakened this type of relationship between the plant and natural enemies. In cotton, for example, the number of volatiles emitted by damaged leaves was seven times lower in commercial varieties than in a naturalized cultivar [54].
Therefore, the physiological mechanisms by which natural enemies tolerate or detoxify need to be understood more, in terms of which constituent volatiles are present and which mixtures manage their identification and trigger a response by natural enemies. Over the long term, field studies are also required to better understand the influence of tomato constituent attractants on the population dynamics of pests and natural enemies.
Author Contributions
Conceptualization, T.C. and J.R.G.; methodology, C.S. and J.R.G.; software, I.L. and M.G.; validation, T.C. and J.G.; formal analysis, I.L. and M.G.; investigation, T.C., J.R.G. and C.S.; resources, J.R.G.; data curation, J.R.G. and M.G.; writing—original draft preparation, J.R.G. and T.C.; writing—review and editing, M.G. and J.G.; visualization, T.C. and J.R.G.; supervision, T.C. and J.G.; project administration, T.C. and J.R.G.; funding acquisition, T.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ministry of Economy, Innovation and Science of the Andalusian Regional Government (Spain), grant number P09 AGR-5000.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Brilli, F.; Loreto, F.; Baccellin, I. Exploiting plant volatile organic compounds (VOCs) in Agriculture to improve sustainable defence strategies and productivity of crops. Front. Plant Sci. 2019, 10, 264. [Google Scholar] [CrossRef] [PubMed]
- Ode, P.J. Plant chemistry and natural enemy fitness: Effects on herbivore and natural enemy interaction. Annu. Rev. Entomol. 2006, 51, 163–185. [Google Scholar] [CrossRef] [PubMed]
- Fellowes, M.D.E.; van Alphen, J.M.; Shameer, K.S. Foraging behaviour. In Jervis’s Insects as Natural Enemies: Practical Perspectives, 3rd ed.; Hardy, I.C.W., Wajnberg, E., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 1–101. [Google Scholar]
- Razo-Belman, R.; Ozuna, C. Volatile organic compounds: A review of their current applications as pest biocontrol and disease management. Horticulturae 2023, 9, 441. [Google Scholar] [CrossRef]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.R.; Bleeker, P.M.; Van Wijk, M.; Schuurink, R.C.; Haring, M.A. Plant volatiles in defence. Adv. Bot. Res. 2009, 51, 613–666. [Google Scholar] [CrossRef]
- Shrivastave, G.; Rogers, M.; Wszelaki, A.; Panthee, D.R.; Chen, F. Plant volatiles-based insect pest management in organic farming. Crit. Rev. Plant Sci. 2010, 29, 123–133. [Google Scholar] [CrossRef]
- Takabayashi, J.; Dicke, M. Plant—Carnivore mutualism through herbivore-induced carnivore attractants. Trends Plant Sci. 1996, 1, 109–113. [Google Scholar] [CrossRef]
- Ferreira, V.; Lopez, R.; Cacho, J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Hoballah, M.E.F.; Tamo, C.; Turlings, T.C. Differential attractiveness of induced odors emitted by eight maize varieties for the parasitoid Cotesia marginiventris: Is quality or quantity important? J. Chem. Ecol. 2002, 28, 951–968. [Google Scholar] [CrossRef]
- Kappers, I.F.; Hoogerbrugge, H.; Bouwmeester, H.J.; Dicke, M. Variation in herbivory-induced volatiles among cucumber (Cucumis sativus L.) varieties has consequences for the attraction of carnivorous natural enemies. J. Chem. Ecol. 2011, 37, 150–160. [Google Scholar] [CrossRef]
- Kappers, I.F.; Verstappen, F.W.; Luckerhoff, L.L.; Bouwmeester, H.J.; Dicke, M. Genetic variation in jasmonic acid-and spider mite-induced plant volatile emission of cucumber accessions and attraction of the predator Phytoseiulus persimilis. J. Chem. Ecol. 2010, 36, 500–512. [Google Scholar] [CrossRef] [PubMed]
- Duffey, S.S.; Stout, M.J. Antinutritive and toxic components of plant defense against insects. Arch. Insect Biochem. Physiol. 1996, 32, 3–37. [Google Scholar] [CrossRef]
- Glas, J.J.; Schimmel, B.C.; Alba, J.M.; Escobar-Bravo, R.; Schuurink, R.C.; Kant, M.R. Plant glandular trichomes as targets for breeding or engineering of resistance to herbivores. Int. J. Mol. Sci. 2012, 13, 17077–17103. [Google Scholar] [CrossRef]
- Scott, J.; Gardner, R. Breeding for resistance to fungal pathogens. In Genetic Improvement of Solanaceous Crops; Publ. Science: Enfield, NH, USA, 2007; pp. 421–456. [Google Scholar]
- Cortesero, A.; Stapel, J.; Lewis, W. Understanding and manipulating plant attributes to enhance biological control. Biol. Control 2000, 17, 35–49. [Google Scholar] [CrossRef]
- Hare, J.D. Plant genetic variation in tritrophic interactions. In Multitrophic Level Interactions; Cambridge University Press: Cambridge, UK, 2002; pp. 8–43. [Google Scholar]
- Vet, L.E.; Dicke, M. Ecology of infochemical use by natural enemies in a tritrophic context. Annu. Rev. Entomol. 1992, 37, 141–172. [Google Scholar] [CrossRef]
- Calvo, F.J.; Lorente, M.J.; Stansly, P.A.; Belda, J.E. Preplant release of Nesidiocoris tenuis and supplementary tactics for control of Tuta absoluta and Bemisa tabaci in greenhouse tomato. Entomol. Exp. Appl. 2012, 143, 111–119. [Google Scholar] [CrossRef]
- Sanchez, C.; Gamez, M.; Burguilo, F.J.; Garay, J.; Cabello, T. Comparison of predator-parasitoid-prey interaction models for different host plant qualities. Community Ecol. 2018, 19, 125–132. [Google Scholar] [CrossRef]
- Cabello, T. Biologia de dos Especies de Trichogramma (Hym.: Trichogrammatidae) Parasitas de Helicoverpa spp. (Lep.: Noctuidae) en Algodonero: Posibilidades de su Empleo como Agentes de Control Biológico. Ph.D. Thesis, Universidad de Córdoba, Córdoba, Spain, 1985. [Google Scholar]
- Cabello, T.; Vargas, P. Estudio con olfactometro de la influencia de la planta y del insecto huesped en la actividad de busqueda de Trichogramma cordubensis y T. sp. p. buesi (Hym.: Trichogrammatidae). Bol. Sanid. Veg. Plagas 1985, 11, 237–241. [Google Scholar]
- Vet, L.E.; Lenteren, J.V.; Heymans, M.; Meelis, E. An airflow olfactometer for measuring olfactory responses of hymenopterous parasitoids and other small insects. Physiol. Entomol. 1983, 8, 97–106. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Cooley, S.S.; Phelan, P.L. Bioassay approaches to assessing behavioral responses of plum curculio adults (Coleoptera: Curculionidae) to host fruit odor. J. Chem. Ecol. 1995, 21, 1073–1084. [Google Scholar] [CrossRef]
- Van Tol, R.; Visser, J.; Sabelis, M. Olfactory responses of the vine weevil, Otiorhynchus sulcatus, to tree odours. Physiol. Entomol. 2002, 27, 213–222. [Google Scholar] [CrossRef]
- Van Tol, R.; Visser, J.; Sabelis, M. Behavioural responses of the vine weevil, Otiorhynchus sulcatus, to semiochemicals from conspecifics, Otiorhynchus salicicola, and host plants. Entomol. Exp. Appl. 2004, 110, 145–150. [Google Scholar] [CrossRef]
- Pettersson, J. An aphid sex attractant. Insect Syst. Evol. 1970, 1, 63–73. [Google Scholar] [CrossRef]
- Sanchez, C.; Gallego, J.R.; Gamez, M.; Cabello, T. Intensive Biological Control in Spanish Greenhouses. Problems of the Success. Int. J. Agric. Biosyst. Eng. 2014, 8, 1123–1127. Available online: https://Waset.org/Publication/9999557 (accessed on 19 February 2024).
- Cabello, T.; Gallego, J.R.; Fernandez, F.J.; Gamez, M.; Vila, E.; Del Pino, M.; Hernandez–Suarez, E. Biological control strategies for the South American tomato moth (Lepidoptera: Gelechiidae) in greenhouse tomatoes. J. Econ. Entomol. 2012, 105, 2085–2096. [Google Scholar] [CrossRef] [PubMed]
- Buttery, R.G.; Ling, L.C.; Light, D.M. Tomato leaf volatile aroma components. J. Agric. Food Chem. 1987, 35, 1039–1042. [Google Scholar] [CrossRef]
- Buttery, R.G.; Ling, L.C. Volatile components of tomato fruit and plant parts: Relationship and biogenesis. In Bioactive Volatile Compounds from Plant; Teranishi, R., Buttery, R.G., Sugisawa, H., Eds.; American Chemical Society: New York, NY, USA, 1993; Volume 525, pp. 23–34. [Google Scholar]
- Proffit, M.; Birgersson, G.; Bengtsson, M.; Reis, R.; Witzgall, P.; Lima, E. Attraction and oviposition of Tuta absoluta females in response to tomato leaf volatiles. J. Chem. Ecol. 2011, 37, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Vila, E.; Cabello, T. Lucha biologica: Utilizacion de entomofagos en cultivos en invernaderos. Agricultura 2012, 81, 10–15. [Google Scholar]
- Hendriks, M.M.; Cruz-Juarez, L.; De Bont, D.; Hall, R.D. Preprocessing and exploratory analysis of chromatographic profiles of plant extracts. Anal. Chim. Acta 2005, 545, 53–64. [Google Scholar] [CrossRef]
- Cusumano, A.; Weldegergis, B.T.; Colazza, S.; Dicke, M.; Fatouros, N.E. Attraction of egg-killing parasitoids toward induced plant volatiles in a multi-herbivore context. Oecologia 2015, 179, 163–174. [Google Scholar] [CrossRef]
- Lins, J.C.; van Loon, J.J.; Bueno, V.H.; Lucas-Barbosa, D.; Dicke, M.; van Lenteren, J.C. Response of the zoophytophagous predators Macrolophus pygmaeus and Nesidiocoris tenuis to volatiles of uninfested plants and to plants infested by prey or conspecifics. BioControl 2014, 59, 707–718. [Google Scholar] [CrossRef]
- Visser, J. Host-plant finding by insects: Orientation, sensory input and search patterns. J. Insect Physiol. 1988, 34, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Iijima, Y.; Okubo, N.; Tanaka, F. Analysis of internal pools of plant volatiles. In Biology of Plant Volatiles, 2nd ed.; Pichersky, E., Dudareva, N., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 27–38. [Google Scholar]
- Gasparini, K.; Gasparini, J.; Therezan, R.; Vicente, M.H.; Sakamoto, T.; Figueira, A.; Zsögön, A.; Peres, L.E. Natural genetic variationin the hairs absent (H) gene increases type-VI glandular trichomes in both wild and domesticated tomatoes. J. Plant Physiol. 2023, 280, 153859. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Garvey, M.; Kaplan, I. Domestication of tomato has reduced the attraction of herbivores natural enemies to pest-damage plants. Agric. For. Entomol. 2018, 20, 390–401. [Google Scholar] [CrossRef]
- Kennedy, G.G. Tomato, pests, parasitoids, and predators: Tritrophic interactions involving the genus Lycopersicon. Annu. Rev. Entomol. 2023, 48, 51–72. [Google Scholar] [CrossRef]
- Romeis, J.; Babendreier, D.; Wackers, F.L.; Shanower, T.G. Habitat and plant specificity of Trichogramma egg parasitoids—Underlying mechanisms and implications. Basic Appl. Ecol. 2005, 6, 215–236. [Google Scholar] [CrossRef]
- Nordlund, D.A.; Chalfant, R.B.; Lewis, W. Response of Trichogramma pretiosum females to volatile synomones from tomato plants. J. Entomol. Sci. 1985, 20, 372–376. [Google Scholar] [CrossRef]
- Raghava, T.; Ravikumar, P.; Hegde, R.; Kush, A. Spatial and temporal volatile organic compound response of select tomato cultivars to herbivory and mechanical injury. Plant Sci. 2010, 179, 520–526. [Google Scholar] [CrossRef]
- Romeis, J.; Shanower, T.G.; Zebit, C.P.W. A simple and effective bioassay to study the effect of plant surface chemicals on the behavior of Trichogramma spp. Z. Pflanzenkr. Pflanzenschutz 1996, 103, 213–216. [Google Scholar]
- Romeis, J.; Shanower, T.; Zebitz, C. Physical and chemical plant characters inhibiting the searching behaviour of Trichogramma chilonis. Entomol. Exp. Appl. 1998, 87, 275–284. [Google Scholar] [CrossRef]
- Bar, D.; Gerling, D.; Rossler, Y. Bionomics of the principal natural enemies attacking Heliothis armigera in cotton fields in Israel. Environ. Entomol. 1979, 8, 468–474. [Google Scholar] [CrossRef]
- Tandon, P.; Bakthavatsalam, N. Plant volatile diversity in different tomato genotypes and its influence on parasitization efficiency of Trichogramma chilonis Ishii on Helicoverpa armigera (Hübner). J. Biol. Control 2007, 21, 271–281. [Google Scholar]
- Glinwood, R.; Ahmed, E.; Qvarfordt, E.; Ninkovic, V. Olfactory learning of plant genotypes by a polyphagous insect predator. Oecologia 2011, 166, 637–647. [Google Scholar] [CrossRef]
- Bjorksten, T.A.; Hoffmann, A.A. Separating the effects of experience, size, egg load, and genotype on host response in Trichogramma (Hymenoptera: Trichogrammatidae). J. Insect Behav. 1998, 11, 129–148. [Google Scholar] [CrossRef]
- Kauffman, W.C.; Kennedy, G.G. Relationship between trichome density in tomato and parasitism of Heliothis spp. (Lepidoptera: Noctuidae) eggs by Trichogramma spp. (Hymenoptera: Trichogrammatidae). Environ. Entom. 1989, 18, 698–704. [Google Scholar] [CrossRef]
- Naselli, M.; Zappala, L.; Gugliuzzo, A.; Garzia, G.T.; Biondi, A.; Rapisarda, C.; Cincotta, F.; Condurso, C.; Verzera, A.; Siscaro, G. Olfactory response of the zoophytophagous mirid Nesidiocoris tenuis to tomato and alternative host plants. Arthropod Plant Interact. 2017, 11, 121–131. [Google Scholar] [CrossRef]
- Urbaneja, A.; Tapia, G.; Stansly, P. Influence of host plant and prey availability on developmental time and surviorship of Nesidiocoris tenius (Het.: Miridae). Biocontrol Sci. Technol. 2005, 15, 513–518. [Google Scholar] [CrossRef]
- Loughrin, J.H.; Manukian, A.; Heath, R.R.; Tumlinson, J.H. Volatiles emitted by different cotton varieties damaged by feeding beet armyworm larvae. J. Chem. Ecol. 1995, 21, 1217–1227. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).