Abstract
Obesity and related metabolic disorders, including chronic inflammation and enhanced oxidative stress, are closely associated with the development and progression of colorectal cancer. Previous epidemiological studies have demonstrated that increased serum uric acid is associated with the risk for various types of cancer, including colon cancer. This study examined the effects of a xanthine oxidase inhibitor allopurinol, widely used as a uric acid lowering medicine, on colorectal tumorigenesis in obese mice. Male C57BL/KsJ-db/db mice were injected with azoxymethane (15 mg/kg body weight) and then received drinking water containing allopurinol (30 mg/kg body weight) for fourteen weeks. At the time of sacrifice, allopurinol treatment significantly inhibited the development of colonic premalignant lesions. In the allopurinol-treated group, cellular proliferation in colonic mucosa was significantly suppressed, which was evaluated by the expression of proliferating cell nuclear antigen. Allopurinol also inhibited macrophage infiltration in the adipose tissue and decreased the serum level of TNF-α. The values of oxidative stress markers were markedly decreased in the allopurinol-treated group compared to those in the control group. These findings suggest that allopurinol attenuated chronic inflammation and decreased oxidative stress, preventing the development of colonic pre-neoplastic lesions in obesity-associated colon tumorigenesis model.
1. Introduction
Colorectal cancer (CRC) is a serious health concern, with high mortality and an increasing incidence worldwide [1]. Early detection and treatment as well as the prevention of CRC, including the use of chemopreventive agents and improvements in lifestyle, especially in patients at high risk for CRC, have received attention. Obesity and the related metabolic syndrome are known to be major risk factors for the development and progression of various malignancies, including CRC [2,3,4]. Metabolic syndrome comprises pathological conditions, such as high blood pressure, hyperglycemia, elevated serum triglyceride levels, and dyslipidemia [5].
Uric acid (UA) is the final product of purine nucleotide catabolism. While UA has potent antioxidant ability [6,7], elevated UA levels cause hyperuricemia, leading to gout. As serum UA levels increase with alcohol consumption and other dietary factors, hyperuricemia is considered to be a lifestyle-related disease. Increased serum UA levels are clearly associated with the prevalence of metabolic syndrome components, although the association has been debated over many years [8]. In addition, epidemiological studies have demonstrated that serum UA is associated with an increased risk of cancers, such as colorectal, breast, and prostate cancer [9,10,11]. Moreover, reducing UA levels contributes to attenuating oxidative stress and chronic inflammation, which are crucial factors for the development of CRC [8].
Allopurinol is known as a UA-lowering medicine (Figure 1a) and is widely used for hyperuricemia and gout. In addition to the effect on UA, the relationship between allopurinol and cancer has been investigated. A previous clinical trial indicated that using allopurinol improved the prognosis of patients with advanced CRC [12]. A recent epidemiological study revealed that allopurinol might decrease the risk of prostate cancer in patients with gout [13], while it was reported that allopurinol had no alteration on the risk of prostate cancer in a population-based cohort study [14]. In an in vitro study, allopurinol exhibited a cytotoxic effect on human prostate cancer cells [15].
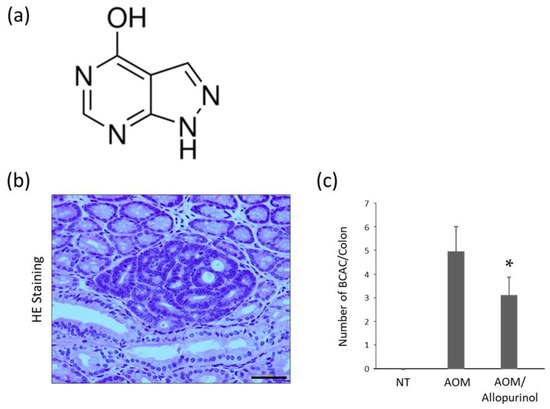
Figure 1.
The chemical structure of allopurinol and AOM-induced colorectal pre-neoplastic lesions in the experimental mice. (a) The chemical structure of allopurinol. (b) A representative image of the β-catenin accumulated crypt (BCAC) stained with hematoxylin and eosin in the colon of the experimental mice. The epithelium has basophilic cytoplasm and hyper chromatic nuclei. Scale bar, 100 µm. (c) The number of BCACs per colon. Each column represents the mean ± SD. Asterisk (*) indicates statistically significant difference compared to the azoxymethane (AOM)-treated group; p < 0.05. NT, no treatment.
In the present study, we focused on serum UA levels in the rodent CRC model and agents against hyperuricemia. A useful preclinical model has been developed in C57BL/KsJ-db/db (db/db) mice, which have a leptin receptor mutation and display hyperphagic obesity following the injection of colonic carcinogen azoxymethane (AOM) [16,17,18]. As db/db mice exhibit hyperuricemia [19], this mouse CRC model appears to be suitable to investigate obesity-related colorectal carcinogenesis accompanied by hyperuricemia. In this study, we investigated the effects of allopurinol on the development of colorectal premalignant lesions β-catenin accumulated crypts (BCACs) [20,21] in the obesity-related colorectal carcinogenesis model described above.
2. Results
2.1. General Observations
As shown in Table 1, the body weight of the mice treated with AOM/allopurinol was significantly lower than that of the mice treated with AOM alone. The relative kidney weight of the mice treated with AOM/allopurinol was significantly higher than that of the mice treated with AOM alone at the end of the experiment. There was no significant difference among all the groups in terms of relative liver weight and relative white adipose tissue weight. During the experiment, there were no clinical symptoms in any of the groups. Histopathological examination did not show toxicity of allopurinol in the liver, spleen, or kidney.

Table 1.
General observations of the experimental mice.
2.2. Serum Parameters
The serum concentrations of alanine aminotransferase (ALT), free fatty acid (FFA), triglyceride, and UA in each group are listed in Table 2. The serum levels of ALT, FFA, and UA in the mice treated with AOM/allopurinol were significantly lower than those in the mice treated with AOM alone. In this study, allopurinol administration had no adverse effect on the serum parameters in the experimental mice. Lowered UA levels in the allopurinol-treated group were expected as the agent works as a xanthine oxidase inhibitor. As increased serum UA was reported to be associated with the exacerbation of steatohepatitis [22], decreased levels of ALT were thought to be related with reduced UA levels in the groups administered allopurinol.
Table 2.
Serum parameters of the experimental mice.
2.3. AOM-Induced Colorectal Pre-Neoplastic Lesions in the Experimental Mice
Colorectal pre-neoplastic lesion BCAC [20,21] developed in the colons of AOM-injected mice. Representative optical microscopic images of AOM-induced BCACs are shown in Figure 1b. The total number of BCACs was significantly lower in the mice treated with AOM plus allopurinol than in mice treated with AOM alone (Figure 1c), indicating that allopurinol might inhibit the development of colorectal premalignant lesions in AOM-injected db/db mice.
2.4. The Expression Levels of mRNAs in Colonic Mucosa of Experimental Mice
The effects of allopurinol on the mRNA expression levels of specific molecules, such as insulin-like growth factor-1 (Igf1), IGF-1 receptor (Igf1r), IGF-binding protein-3 (Igfbp3), and proliferating cell nuclear antigen (Pcna), were examined by quantitative real-time reverse transcription-polymerase chain reaction (qRT-PCR) analysis. While there were no significant differences among the experimental groups with respect to the levels of Igf1, Igf1r, and Igfbp3, the levels of Pcna were markedly downregulated following allopurinol treatment in AOM-injected mice (Figure 2a). A previous report demonstrated that IGF/IGF-1R signaling is essential for obesity-related CRC development [23,24]. The results from the present study demonstrate that allopurinol had no significant effect on this pathway.
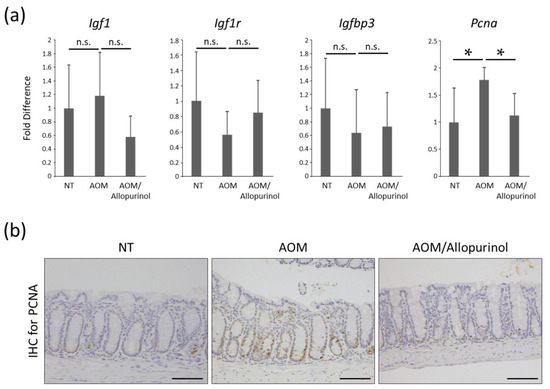
Figure 2.
Effects of allopurinol on the IGF/IGF1R axis and cellular proliferation in the colonic mucosa of the experimental mice. (a) The mRNA expression levels of Igf1, Igf1r, Igfbp3, and Pcna in the colonic mucosa were analyzed by quantitative real-time RT-PCR with specific primers. Each column represents the mean ± SD. Asterisk (*) indicates the statistically significant difference between the groups; p < 0.05. (b) Evaluation of cellular proliferation in colonic mucosa of the experimental mice using an antibody for proliferating cell nuclear antigen (PCNA). Representative images are shown. Scale bars, 100 µm. AOM, azoxymethane; n.s., not significant; IHC, immunohistochemistry. NT, no treatment.
2.5. Cellular Proliferation in Colonic Mucosa of Experimental Mice
Treatment with allopurinol markedly decreased PCNA in colon mucosa evaluated by immunohistochemistry (Figure 2b). This finding, together with the result of the Pcna mRNA expression level described above, might indicate that allopurinol significantly inhibited cellular proliferation in the colonic mucosa of AOM-injected db/db mice, leading to the suppression of the development of premalignant lesions.
2.6. Immunohistochemistry for F4/80 in Adipose Tissue of the Experimental Mice
Immunohistochemical staining for F4/80 was performed to evaluate the chronic inflammation associated with the infiltration of macrophages in white adipose tissue. Compared with mice treated with AOM alone, the number of F4/80 positive cells was 0.31-fold lower in mice treated with AOM/allopurinol (Figure 3), suggesting that allopurinol administration attenuated inflammation in adipose tissue.
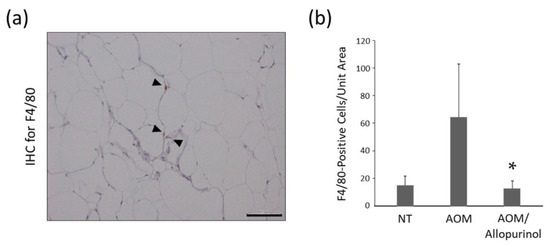
Figure 3.
Effect of allopurinol treatment on macrophage infiltration in the adipose tissue of the experimental mice. (a) A representative picture of immunohistochemical staining for F4/80 in the adipose tissue of the experimental mice. F4/80-stained cells are indicated by arrow heads. Scale bar, 100 µm. (b) The number of F4/80-positive cells per unit area was counted. The values are expressed as the mean ± SD. Asterisk (*) indicates the statistically significant difference compared to the azoxymethane (AOM)-treated group; p < 0.05. IHC, immunohistochemistry; NT, no treatment.
2.7. Serum Levels of TNF-α in the Experimental Mice
Chronic inflammation in the adipose tissue is known to contribute to the increased production of tumor necrosis factor-alpha (TNF-α). Serum levels of TNF-α was analyzed by enzyme-linked immunosorbent assay (ELISA) (Figure 4a). Previously, serum TNF-α levels were reported to be significantly elevated in the AOM-administered db/db mice compared to the no treatment db/db mice [25]. The TNF-α levels were significantly lower in the AOM/allopurinol group than in the AOM group. Because TNF-α is considered an essential tumor promoter in inflammation-related tumorigenesis [26], the administration of allopurinol might contribute to the suppression of colonic neoplasms by decreasing the TNF-α levels.
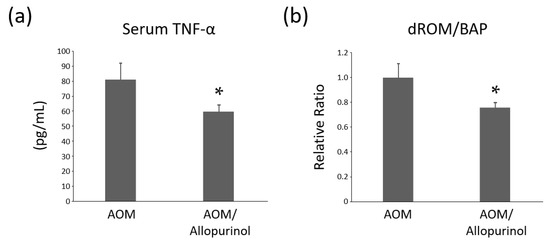
Figure 4.
Effects of allopurinol on serum TNF-α levels and oxidative stress in the experimental mice. (a) Serum concentrations of TNF-α were determined by ELISA. (b) Serum levels of derivatives of reactive oxygen metabolite (d-ROM) and biological antioxidant potential (BAP) were evaluated and the ratio was compared as a relative value. Each column represents the mean ± SD. The asterisk (*) indicates the statistically significant difference compared to azoxymethane (AOM)-treated group; p < 0.05.
2.8. Systemic Oxidative Stress Levels of the Experimental Mice
In the previous study, systemic oxidative stress was evaluated in db/db mice, and the stress was markedly enhanced in the AOM-administered mice compared to no treatment mice [25]. In our study, to investigate systemic oxidative stress, tests for the reactive oxygen metabolite (d-ROM) and biological antioxidant potential (BAP) were performed, and the ratio of d-ROM/BAP was calculated and compared (Figure 4b). The ratio, indicating the degree of oxidative stress [27], was markedly lower in the AOM/allopurinol group than in the AOM alone group, suggesting that allopurinol administration attenuated systemic oxidative stress in AOM-treated db/db mice.
3. Discussion
Increased serum UA, namely, hyperuricemia, is associated with an excess cancer risk and mortality, although UA is theoretically thought to be protective against cancer owing to its antioxidant properties [6]. Furthermore, hyperuricemia increases cancer prevalence and mortality among patients with cancer. In addition, obesity and diabetes are associated with chronic inflammation, cancer, and hyperuricemia, suggesting that UA may represent an important link between these disorders and cancer development [28]. In the present study, we demonstrated that the xanthine oxidase inhibitor, allopurinol, which is widely used for lowering serum UA levels, markedly suppressed the development of colorectal pre-neoplastic lesions induced by AOM in obese db/db mice. The anti-tumorigenesis action appeared to be caused by the attenuation of chronic inflammation and a reduction in oxidative stress.
Chronic inflammation is one of the most important mechanisms in the development of obesity-associated CRC. Remarkably, the enhancement of inflammation in adipose tissue is considered essential for linking obesity and CRC development and progression [29]. During the early phase of systemic inflammation, macrophage infiltration into white adipose tissues occurs as a representative condition. In the meantime, TNF-α is produced from the adipose tissue and affects various organs as a pro-inflammatory cytokine [30]. In our study, the infiltration of macrophages in adipose tissue, evaluated using immunohistochemistry for F4/80, was significantly inhibited following the administration of allopurinol. In addition, serum levels of TNF-α, analyzed using ELISA, were markedly reduced in allopurinol-administered mice, which might be owing to the attenuation of inflammation in adipose tissues. These results might indicate that attenuating inflammation in the adipose tissue by reducing macrophage infiltration is crucial to suppress the development of colonic malignant lesions.
Obesity and the related chronic inflammation were often accompanied by increased oxidative stress. Oxidative stress is enhanced by the excess generation of reactive oxygen species [31], which are derivatives of molecular oxygen, including superoxide and hydrogen peroxide, and can induce mutagenic changes and damage DNA repair functions, leading to carcinogenesis [32,33]. In the present study, allopurinol administration significantly decreased the levels of oxidative stress markers and the ratio of serum d-ROM/BAP in AOM-treated mice. This finding appeared to indicate that the attenuation of oxidative stress played a vital role in suppressing the development of premalignant lesions in the colorectum of experimental mice. While UA is known to possess antioxidant effects [6], the mechanisms by which allopurinol, a UA-lowering agent, attenuated systemic oxidative stress were not revealed in the present study. The altered condition following allopurinol treatment is consistent with a previous report showing reduced oxidative stress by this agent [34].
In this study, we also focused on the effects of allopurinol on the lipid metabolism in the experimental mice. The weights of adipose tissue tended to be heavier in groups 2–4 than that of the no treatment group, but there were no statistically significant differences. The results demonstrated that the serum FFA level in the AOM/allopurinol-treated group was significantly lower compared to that of the AOM-treated group, while there was no marked difference in the level of FFA between the allopurinol-treated group and the no treatment control group. Although a previous study demonstrated that the serum UA concentration was correlated with serum triglyceride [22], in our study there was no significant difference in the serum triglyceride levels among the groups in spite of a marked decrease in UA levels by allopurinol treatment. According to a previous retrospective study, allopurinol modestly decreased serum triglyceride levels, but did not affect cholesterol [35], while a systematic review with meta-analysis demonstrated that allopurinol had no improvement effect on serum lipid levels [36]. These findings indicate that allopurinol may have no significant effect on lipid metabolism, or at least in humans.
The IGF/IGF-1 receptor signaling is known to contribute to colorectal carcinogenesis in obese and diabetic mice [23,24]. As allopurinol treatment exhibited no significant alteration in the gene expression associated with the IGF/IGF-1 receptor axis, the preventive effects of this agent were presumably through the attenuation of both oxidative stress and chronic inflammation. The present study, however, did not investigate the protein levels of IGFs, IGF receptors, and their related molecules. In addition, it was not examined in detail how allopurinol affects stress-related pathways, which might be one of the main mechanisms to inhibit colon tumorigenesis. These are considered the limitations of this study.
In summary, the present study demonstrated the chemopreventive effects of allopurinol on early-phase obesity-related colon tumorigenesis. As the risk of CRC is increased by obesity and related metabolic disorders, targeting obesity-associated abnormalities, including chronic inflammation and oxidative stress, might be a potential preventive and therapeutic strategy for obese patients with CRC. Allopurinol is considered a possible and practical candidate for this direction because it has been widely used in clinical practice without severe adverse effects for patients with metabolic syndrome. Further studies should be performed to investigate the effects of allopurinol, directly or indirectly, on oxidative stress and inflammation, as well as to examine the chemopreventive effects of CRC development in clinical trials.
4. Materials and Methods
4.1. Animals and Care Conditions
Thirty-six male db/db mice were obtained from Japan SLC Inc. (Shizuoka, Japan) and were used for experiments after acclimatized rearing for a week. The animals were cared for and humanely maintained at the Gifu University Life Science Research Center in accordance with the Institutional Animal Care Guidelines. The mice were housed in cages with free access to a basic diet CE-2 obtained from Oriental Yeast (Tokyo, Japan) and sterilized tap water. The bedding was changed once a week. The environmental conditions inside the room were maintained at a humidity of 50% ± 20% and a temperature of 23 ± 1 °C; a 12 h alternating light/dark cycle was followed.
4.2. Chemicals and Administration Methods
AOM was obtained from Wako Pure Chemical Co. (Osaka, Japan). AOM was intraperitoneally injected once weekly for 1–4 weeks after starting the experiment. The dose was formulated by diluting AOM with ultrapure water to obtain a final dose of 15 mg/kg body weight. The model, AOM-induced colorectal carcinogenesis in db/db mice, used in this study, was established and reported in 2004 for the examination of obese- and diabetes-related colorectal carcinogenesis [37], and has been used in a number of researches [16,17,18,25,38,39,40]. Allopurinol was obtained from Wako Pure Chemical Co. (Osaka, Japan) and was dissolved in ultrapure water to obtain a final dose of 30 mg/kg body weight. These solutions were poured into their respective bottles and exchanged twice a week.
4.3. Experimental Procedure
At 5 weeks of age, the mice were randomly divided into four experimental groups and treated as follows: no treatment (group 1, n = 6), AOM alone (group 2, n = 12), AOM plus allopurinol (group 3, n = 12), and allopurinol alone (group 4, n = 6). The mice in groups 2 and 3 were intraperitoneally injected with AOM (15 mg/kg body weight) once weekly for 1–4 weeks after starting the experiment. At 9 weeks of age, the mice in groups 3 and 4 received drinking water containing allopurinol until the end of the experiment. Allopurinol was administered via drinking water at a dose of 30 mg/kg body weight. At 22 weeks (after the 14 week allopurinol treatment) of age, all mice were euthanized for analysis. This experimental protocol was approved by the Committee of Institutional Animal Experiments of Gifu University (the authorization code 30-7 on 5 April 2018).
4.4. Histopathological Examination
At the end of the experiment (22 weeks of age), all mice were euthanized for histopathological analysis. The liver, kidneys, spleen, white adipose tissue, and colorectum were excised. The colorectum was dissected longitudinally and fixed on filter paper in 10% buffered formalin for more than 24 h. It was further divided into the rectum portion (approximately 1 cm in length, oral side from the dentate line) and the colon portion. Each portion and the other tissues were paraffin embedded, stained with hematoxylin and eosin, and histologically analyzed. Immunohistochemical staining was performed for PCNA and was performed in colon mucosa using the labeled streptavidin-biotin method (LSAB kit; Dako, Glostrup, Denmark) as described previously [16,27]. Primary antibodies for PCNA were obtained from Santa Cruz Biotechnology (1:100 dilution, sc-7907; Dallas, TX, USA). Immunohistochemical staining for F4/80 was also performed to examine macrophage infiltration in adipose tissues, according to a previous study with a primary antibody (1:100 dilution, ab111101; Abcam, Cambridge, United Kingdom) [17].
4.5. Clinical Chemistry
Blood samples were collected from the inferior vena cava of the mice at sacrifice after a 12 h fasting period. The serum was centrifuged from whole blood and used for chemical analyses. Serum alanine aminotransferase (ALT), free fatty acid (FFA), triglyceride, and UA levels were determined by a commercial laboratory (SRL, Inc., Tokyo, Japan). To investigate the systemic oxidative stress, d-ROMs and BAP were evaluated using FREE Carpe Diem (Diacron International s.r.l., Grosseto, Italy) in accordance with the manufacturer’s protocol. Serum TNF-α levels were measured using the Mouse TNF-α ELISA Kit (AKMTM-011, Shibayagi Co. Ltd., Gunma, Japan) in accordance with the manufacturer’s protocol.
4.6. Extraction of mRNA and Quantitative Real-Time RT-PCR Analysis
The expression levels of mRNA in the colon mucosa of the experimental mice were examined using qRT-PCR analysis. Total RNA was isolated from the scraped colonic mucosa of all experimental mice using the PureLinkTM RNA Mini Kit (Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s instructions. Complementary DNA (cDNA) was amplified from the total RNA of each sample using a High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). The primers used for the amplification of Igf1, Igf1r, Igfbp3, Pcna, and 18s were previously reported [17,41] or as follows: Igf1r forward, 5′-GAG AAT TTC CTT CAC AAT TCC ATC-3′ and reverse, 5′-CAC TTG CAT GAC GTC TCT CC-3′ and Pcna forward, 5′-CTA GCC ATG GGC GTG AAC-3′ and reverse, 5′-GAA TAC TAG TGC TAA GGT GTC TGC ATT-3′. Real-time PCR was performed in a LightCycler (Roche Diagnostics Co., Indianapolis, IN, USA) with SYBR Premix Ex Taq (TaKaRa Bio, Shiga, Japan) as previously described [42]. The expression levels of these genes were normalized to 18s gene expression levels.
4.7. Statistical Analysis
The results are presented as the mean ± standard deviation. The Tukey–Kramer multiple comparison test was performed to compare each experimental group. The Mann–Whitney U test was performed between the two groups when a specific p-value was required, or for the re-test for the presence or absence of a significant difference between the two groups. All differences were considered significant at p < 0.05.
5. Conclusions
The preventive effects of the UA lowering agent allopurinol on the development of obesity-related CRC were demonstrated in this study. The mechanisms appeared to be the attenuation of chronic inflammation and oxidative stress by allopurinol administration. The risk of CRC is higher in obese individuals and in those with related metabolic abnormalities, including chronic inflammation and oxidative stress. Therefore, the use of allopurinol for CRC chemoprevention can be an effective strategy in patients with obesity.
Author Contributions
Conceptualization, J.K. and Y.S.; methodology, J.K. and Y.S.; validation, M.K., H.S. and T.I. (Takashi Ibuka); investigation, J.K., Y.S., K.Y., T.M., T.I. (Takayasu Ideta), H.N., S.N., M.K. and H.S.; writing—original draft preparation, J.K.; writing—review and editing, Y.S., T.T. and M.S.; supervision, M.S.; funding acquisition, Y.S., M.K. and M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by JSPS KAKENHI Grant Numbers JP17K15936, JP18K07968, and JP19K08465.
Acknowledgments
The authors thank Miho Yagi, Chiyoko Sano, Hitomi Fujisawa, and Eriko Kunishima for secretarial assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef]
- González, N.; Prieto, I.; Del Puerto-Nevado, L.; Portal-Nuñez, S.; Ardura, J.A.; Corton, M.; Fernández-Fernández, B.; Aguilera, O.; Gomez-Guerrero, C.; Mas, S.; et al. 2017 update on the relationship between diabetes and colorectal cancer: Epidemiology, potential molecular mechanisms and therapeutic implications. Oncotarget 2017, 8, 18456–18485. [Google Scholar] [CrossRef] [PubMed]
- Giovannucci, E. Metabolic syndrome, hyperinsulinemia, and colon cancer: A review. Am. J. Clin. Nutr. 2007, 86, s836–s842. [Google Scholar] [CrossRef]
- Giovannucci, E.; Michaud, D. The role of obesity and related metabolic disturbances in cancers of the colon, prostate, and pancreas. Gastroenterology 2007, 132, 2208–2225. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M. Metabolic syndrome: A multiplex cardiovascular risk factor. J. Clin. Endocrinol. Metab. 2007, 92, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Glantzounis, G.K.; Tsimoyiannis, E.C.; Kappas, A.M.; Galaris, D.A. Uric acid and oxidative stress. Curr. Pharm. Des. 2005, 11, 4145–4151. [Google Scholar] [CrossRef]
- Mikami, T.; Sorimachi, M. Uric acid contributes greatly to hepatic antioxidant capacity besides protein. Physiol. Res. 2017, 66, 1001–1007. [Google Scholar] [CrossRef]
- Ali, N.; Miah, R.; Hasan, M.; Barman, Z.; Mou, A.D.; Hafsa, J.M.; Trisha, A.D.; Hasan, A.; Islam, F. Association between serum uric acid and metabolic syndrome: A cross-sectional study in Bangladeshi adults. Sci. Rep. 2020, 10, 7841. [Google Scholar] [CrossRef]
- Hammarsten, J.; Damber, J.-E.; Peeker, R.; Mellström, D.; Högstedt, B. A higher prediagnostic insulin level is a prospective risk factor for incident prostate cancer. Cancer Epidemiol. 2010, 34, 574–579. [Google Scholar] [CrossRef]
- Siddiqui, A.A. Metabolic syndrome and its association with colorectal cancer: A review. Am. J. Med. Sci. 2011, 341, 227–231. [Google Scholar] [CrossRef]
- Bjørge, T.; Lukanova, A.; Jonsson, H.; Tretli, S.; Ulmer, H.; Manjer, J.; Stocks, T.; Selmer, R.; Nagel, G.; Almquist, M.; et al. Metabolic syndrome and breast cancer in the me-can (metabolic syndrome and cancer) project. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2010, 19, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Salim, A.S. Oxygen-derived free-radical scavengers prolong survival in colonic cancer. Chemotherapy 1992, 38, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Shih, H.-J.; Kao, M.-C.; Tsai, P.-S.; Fan, Y.-C.; Huang, C.-J. Long-term allopurinol use decreases the risk of prostate cancer in patients with gout: A population-based study. Prostate Cancer Prostatic Dis. 2017, 20, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Kukko, V.; Kaipia, A.; Talala, K.; Taari, K.; Tammela, T.L.J.; Auvinen, A.; Murtola, T.J. Allopurinol and the risk of prostate cancer in a Finnish population-based cohort. Prostate Cancer Prostatic Dis. 2019, 22, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Yoshida, T.; Goda, A.E.; Horinaka, M.; Yano, K.; Shiraishi, T.; Wakada, M.; Mizutani, Y.; Miki, T.; Sakai, T. Anti-gout agent allopurinol exerts cytotoxicity to human hormone-refractory prostate cancer cells in combination with tumor necrosis factor-related apoptosis-inducing ligand. Mol. Cancer Res. 2008, 6, 1852–1860. [Google Scholar] [CrossRef] [PubMed]
- Fukuta, K.; Shirakami, Y.; Maruta, A.; Obara, K.; Iritani, S.; Nakamura, N.; Kochi, T.; Kubota, M.; Sakai, H.; Tanaka, T.; et al. Preventive Effects of Pentoxifylline on the Development of Colonic Premalignant Lesions in Obese and Diabetic Mice. Int. J. Mol. Sci. 2017, 18, 413. [Google Scholar] [CrossRef]
- Kato, J.; Shirakami, Y.; Ohnishi, M.; Mizutani, T.; Kubota, M.; Sakai, H.; Ibuka, T.; Tanaka, T.; Shimizu, M. Suppressive effects of the sodium-glucose cotransporter 2 inhibitor tofogliflozin on colorectal tumorigenesis in diabetic and obese mice. Oncol. Rep. 2019, 42, 2797–2805. [Google Scholar] [CrossRef]
- Kato, J.; Shirakami, Y.; Mizutani, T.; Kubota, M.; Sakai, H.; Ibuka, T.; Shimizu, M. Alpha-Glucosidase Inhibitor Voglibose Suppresses Azoxymethane-Induced Colonic Preneoplastic Lesions in Diabetic and Obese Mice. Int. J. Mol. Sci. 2020, 21, 2226. [Google Scholar] [CrossRef]
- Kosugi, T.; Nakayama, T.; Heinig, M.; Zhang, L.; Yuzawa, Y.; Sanchez-Lozada, L.G.; Roncal, C.; Johnson, R.J.; Nakagawa, T. Effect of lowering uric acid on renal disease in the type 2 diabetic db/db mice. Am. J. Physiol. Renal Physiol. 2009, 297, F481–F488. [Google Scholar] [CrossRef]
- Bird, R.P.; Good, C.K. The significance of aberrant crypt foci in understanding the pathogenesis of colon cancer. Toxicol. Lett. 2000, 112–113, 395–402. [Google Scholar] [CrossRef]
- Yamada, Y.; Mori, H. Pre-cancerous lesions for colorectal cancers in rodents: A new concept. Carcinogenesis 2003, 24, 1015–1019. [Google Scholar] [CrossRef]
- Wan, X.; Xu, C.; Lin, Y.; Lu, C.; Li, D.; Sang, J.; He, H.; Liu, X.; Li, Y.; Yu, C. Uric acid regulates hepatic steatosis and insulin resistance through the NLRP3 inflammasome-dependent mechanism. J. Hepatol. 2016, 64, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Shirakami, Y.; Sakai, H.; Adachi, S.; Hata, K.; Hirose, Y.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. (-)-Epigallocatechin gallate suppresses azoxymethane-induced colonic premalignant lesions in male C57BL/KsJ-db/db Mice. Cancer Prev. Res. 2008, 1. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Deguchi, A.; Hara, Y.; Moriwaki, H.; Weinstein, I.B. EGCG inhibits activation of the insulin-like growth factor-1 receptor in human colon cancer cells. Biochem. Biophys. Res. Commun. 2005, 334, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Kochi, T.; Shimizu, M.; Sumi, T.; Kubota, M.; Shirakami, Y.; Tanaka, T.; Moriwaki, H. Inhibitory effects of astaxanthin on azoxymethaneinduced colonic preneoplastic lesions in C57/BL/KsJ-db/db mice. BMC Gastroenterol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Szlosarek, P.; Charles, K.A.; Balkwill, F.R. Tumour necrosis factor-α as a tumour promoter. Eur. J. Cancer 2006. [Google Scholar] [CrossRef]
- Kochi, T.; Shimizu, M.; Ohno, T.; Baba, A.; Sumi, T.; Kubota, M.; Shirakami, Y.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. Preventive effects of the angiotensin-converting enzyme inhibitor, captopril, on the development of azoxymethane-induced colonic preneoplastic lesions in diabetic and hypertensive rats. Oncol. Lett. 2014, 8, 223–229. [Google Scholar] [CrossRef]
- Fini, M.A.; Elias, A.; Johnson, R.J.; Wright, R.M. Contribution of uric acid to cancer risk, recurrence, and mortality. Clin. Transl. Med. 2012, 1, 16. [Google Scholar] [CrossRef] [PubMed]
- Gunter, M.J.; Leitzmann, M.F. Obesity and colorectal cancer: Epidemiology, mechanisms and candidate genes. J. Nutr. Biochem. 2006, 17, 145–156. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W.J. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Newsholme, P.; Cruzat, V.F.; Keane, K.N.; Carlessi, R.; de Bittencourt, P.I.H.J. Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochem. J. 2016, 473, 4527–4550. [Google Scholar] [CrossRef] [PubMed]
- Schetter, A.J.; Heegaard, N.H.H.; Harris, C.C. Inflammation and cancer: Interweaving microRNA, free radical, cytokine and p53 pathways. Carcinogenesis 2010, 31, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Sethi, G.; Shanmugam, M.K.; Ramachandran, L.; Kumar, A.P.; Tergaonkar, V. Multifaceted link between cancer and inflammation. Biosci. Rep. 2012, 32, 1–15. [Google Scholar] [CrossRef]
- Wang, W.; Ding, X.-Q.; Gu, T.-T.; Song, L.; Li, J.-M.; Xue, Q.-C.; Kong, L.-D. Pterostilbene and allopurinol reduce fructose-induced podocyte oxidative stress and inflammation via microRNA-377. Free Radic. Biol. Med. 2015, 83, 214–226. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Y.-P.; Qu, Y.; Jie, L.-G.; Deng, J.-X.; Yu, Q.-H. Efficacy of uric acid-lowering therapy on hypercholesterolemia and hypertriglyceridemia in gouty patients. Int. J. Rheum. Dis. 2019, 22, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- de Castro, V.M.F.; de Melo, A.C.; Belo, V.S.; Chaves, V.E. Effect of allopurinol and uric acid normalization on serum lipids hyperuricemic subjects: A systematic review with meta-analysis. Clin. Biochem. 2017, 50, 1289–1297. [Google Scholar] [CrossRef]
- Hirose, Y.; Hata, K.; Kuno, T.; Yoshida, K.; Sakata, K.; Yamada, Y.; Tanaka, T.; Reddy, B.S.; Mori, H. Enhancement of development of azoxymethane-induced colonic premalignant lesions in C57BL/KsJ-db/db mice. Carcinogenesis 2004, 25, 821–825. [Google Scholar] [CrossRef]
- Shimizu, M.; Shirakami, Y.; Iwasa, J.; Shiraki, M.; Yasuda, Y.; Hata, K.; Hirose, Y.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. Supplementation with branched-chain amino acids inhibits azoxymethane-induced colonic preneoplastic lesions in male C57BL/KsJ-db/db mice. Clin. Cancer Res. 2009, 15. [Google Scholar] [CrossRef]
- Yasuda, Y.; Shimizu, M.; Shirakami, Y.; Sakai, H.; Kubota, M.; Hata, K.; Hirose, Y.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. Pitavastatin inhibits azoxymethane-induced colonic preneoplastic lesions in C57BL/KsJ-db/db obese mice. Cancer Sci. 2010, 101, 1701–1707. [Google Scholar] [CrossRef]
- Kubota, M.; Shimizu, M.; Sakai, H.; Yasuda, Y.; Ohno, T.; Kochi, T.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. Renin-angiotensin system inhibitors suppress azoxymethane-induced colonic preneoplastic lesions in C57BL/KsJ-db/db obese mice. Biochem. Biophys. Res. Commun. 2011, 410, 108–113. [Google Scholar] [CrossRef]
- Shirakami, Y.; Shimizu, M.; Kubota, M.; Ohno, T.; Kochi, T.; Nakamura, N.; Sumi, T.; Tanaka, T.; Moriwaki, H.; Seishima, M. Pentoxifylline prevents nonalcoholic steatohepatitis-related liver pre-neoplasms by inhibiting hepatic inflammation and lipogenesis. Eur. J. Cancer Prev. Off. J. Eur. Cancer Prev. Organ 2016, 25, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Shirakami, Y.; Gottesman, M.E.; Blaner, W.S. Diethylnitrosamine-induced hepatocarcinogenesis is suppressed in lecithin: Retinol acyltransferase-deficient mice primarily through retinoid actions immediately after carcinogen administration. Carcinogenesis 2012, 33. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).