Sedative–Hypnotic Activity of the Water Extracts of Coptidis Rhizoma in Rodents

 , ,
, ,
Abstract
:1. Introduction
2. Results
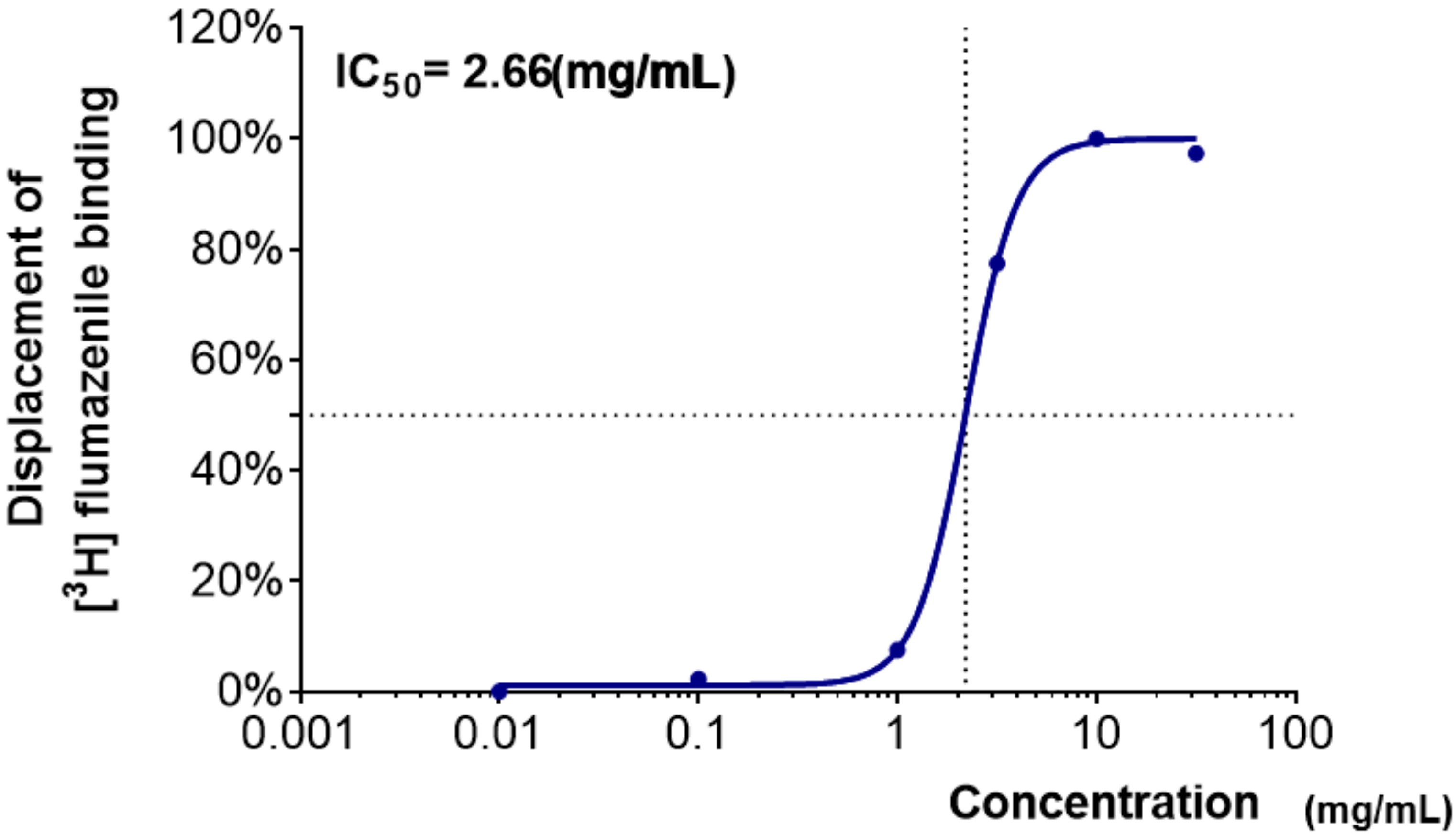
2.1. Binding Affinity of Medicinal Plants Extracts to the GABAA–BZD Receptors
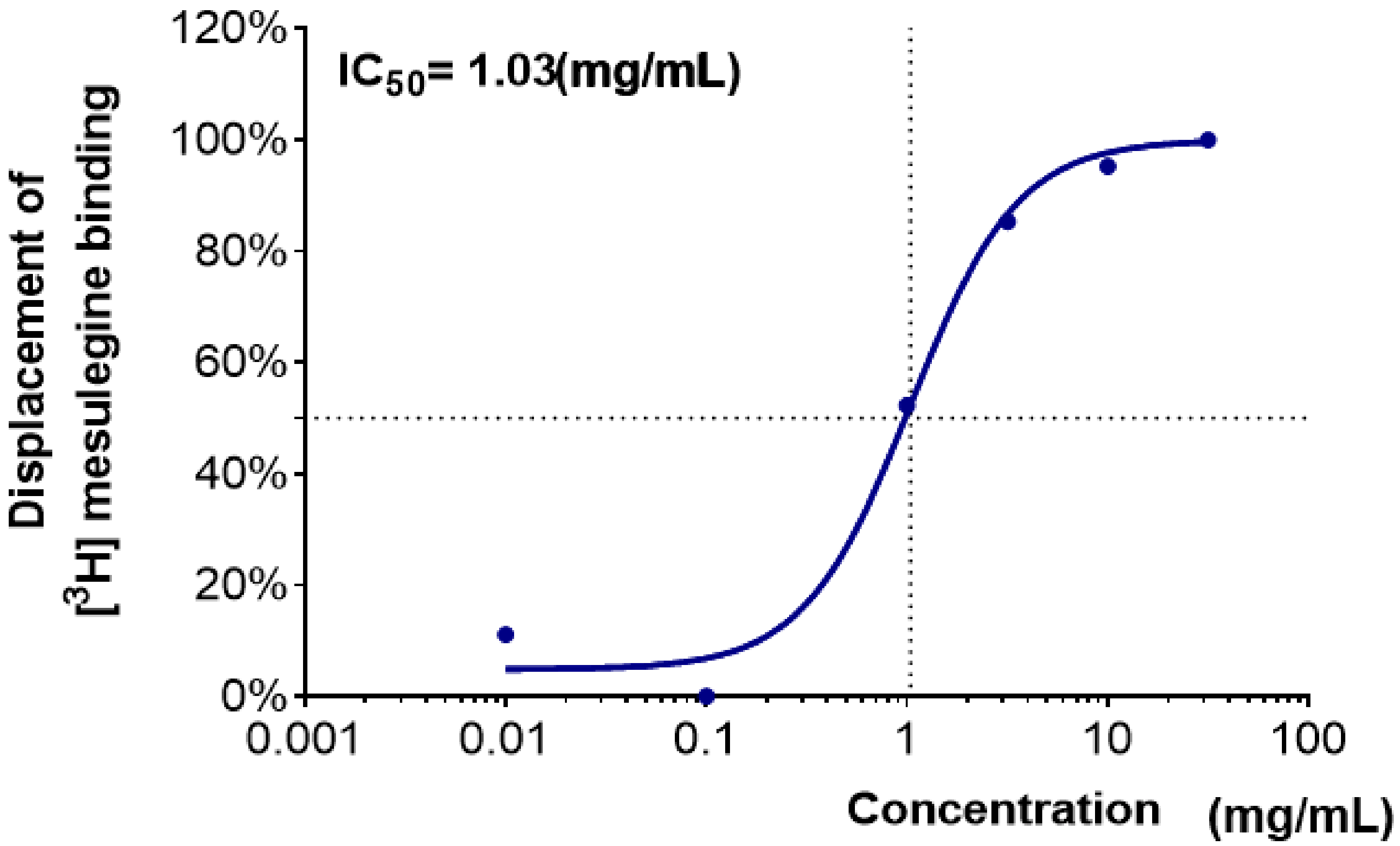
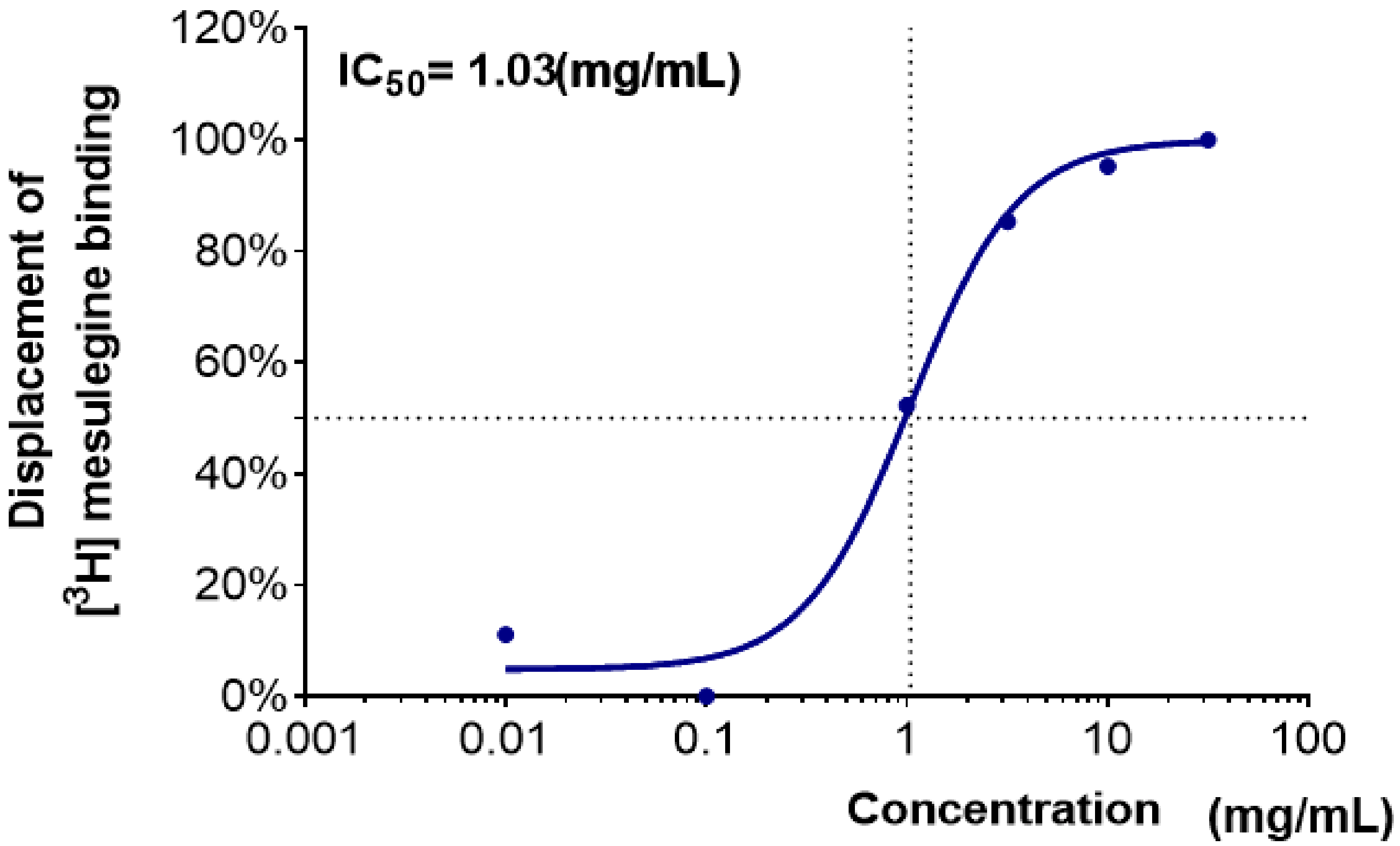
2.2. Binding Affinity of Plant Extracts to the 5-HT2C Receptors
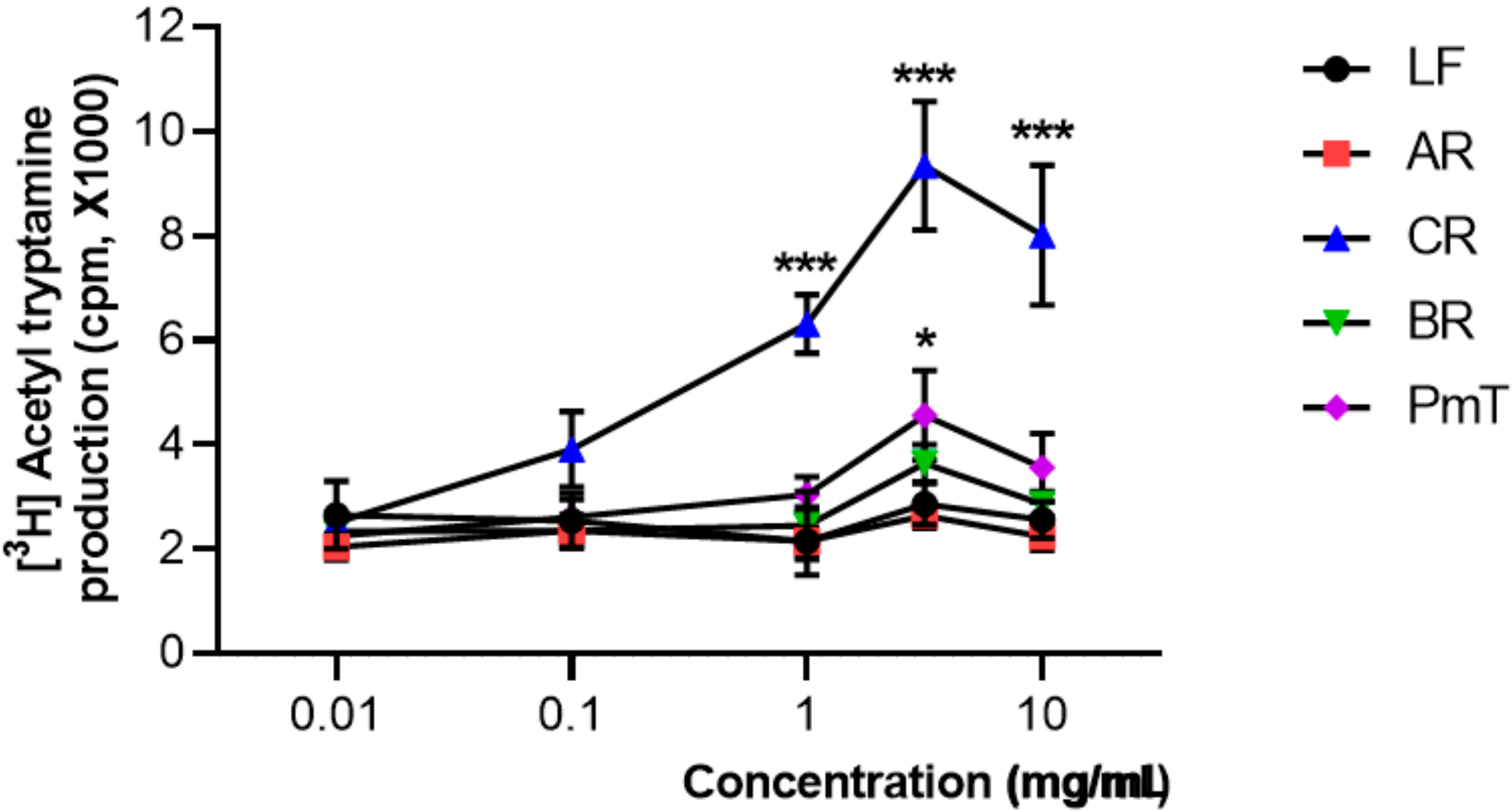
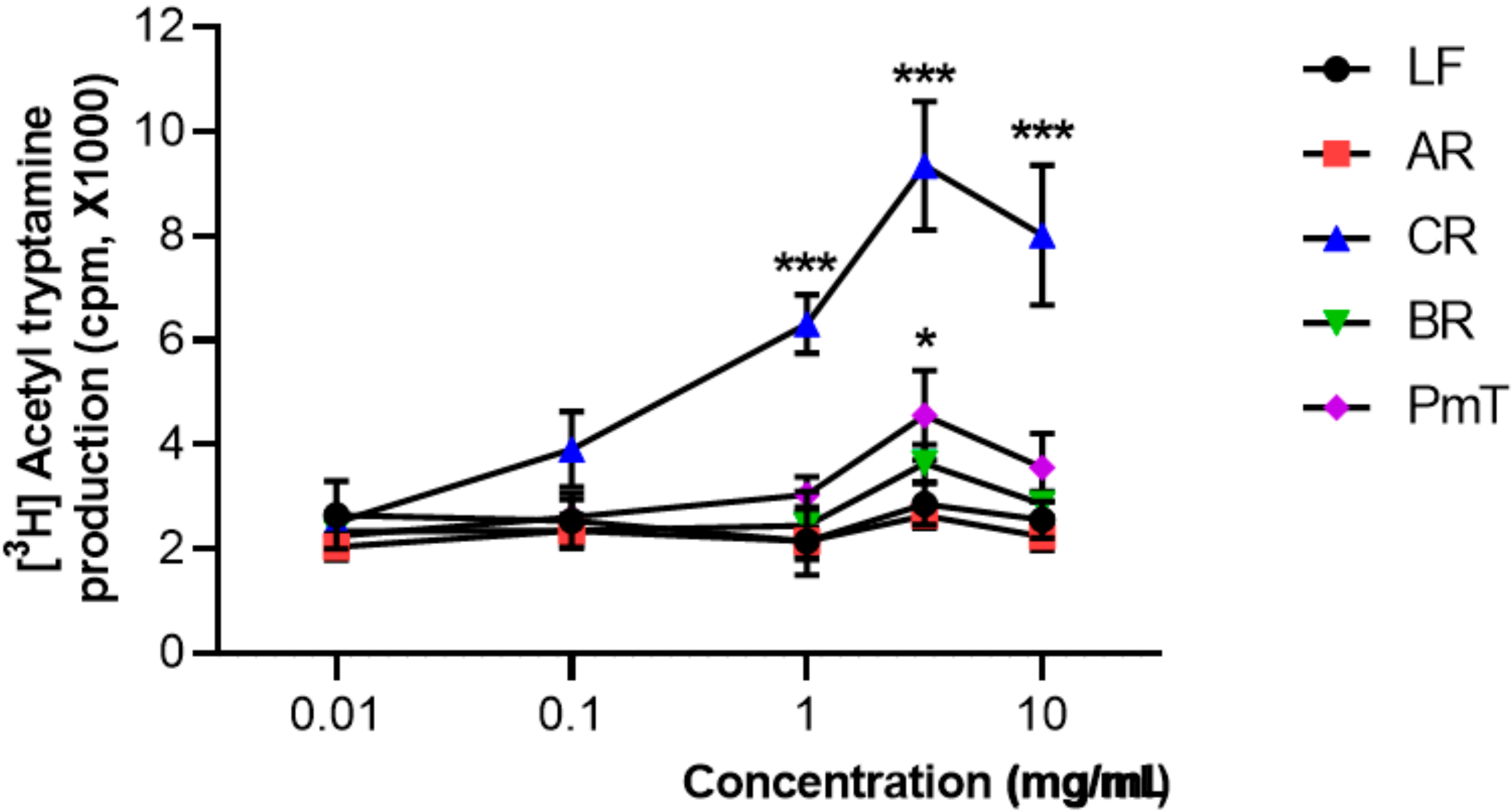
2.3. AANAT Assay
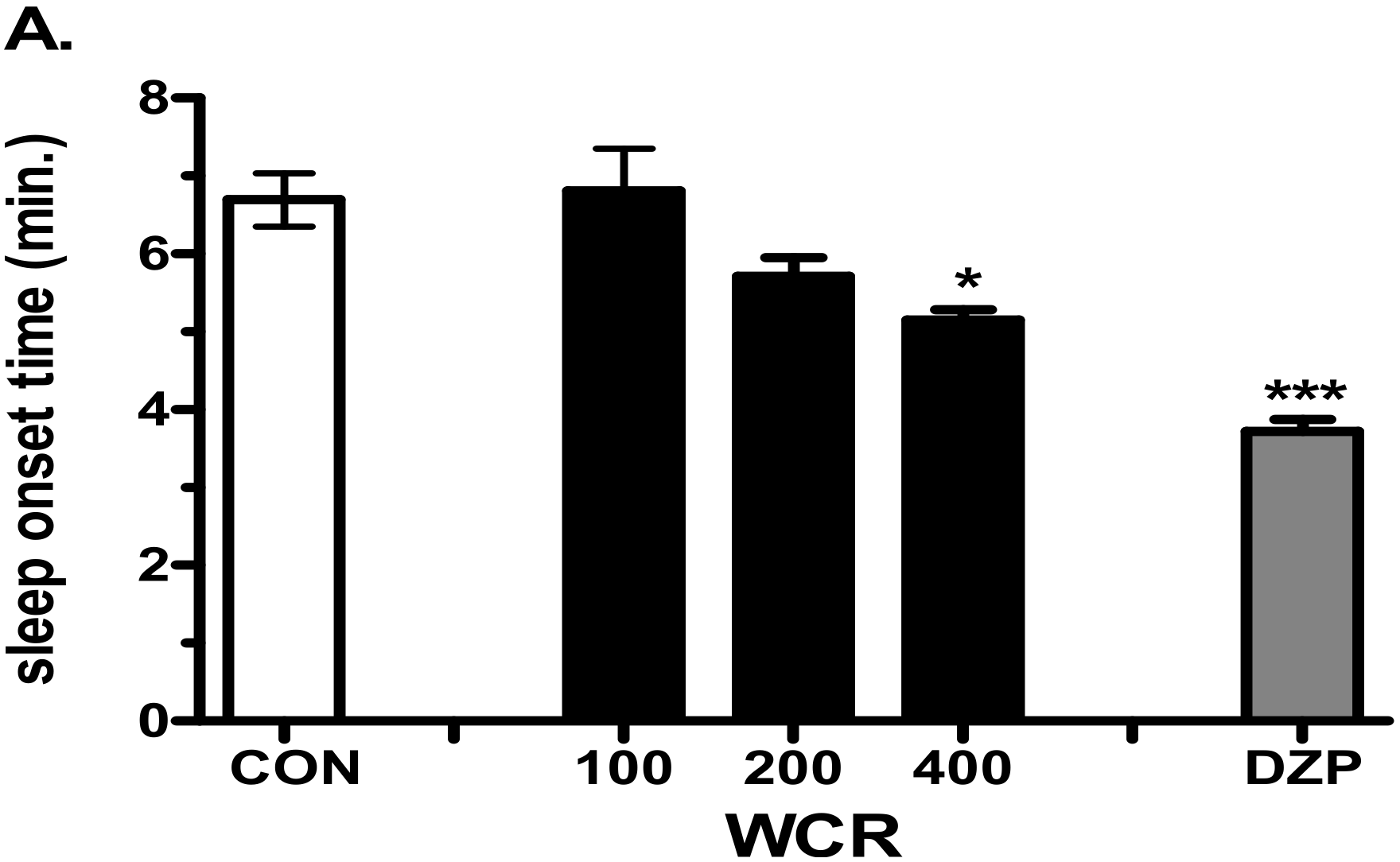
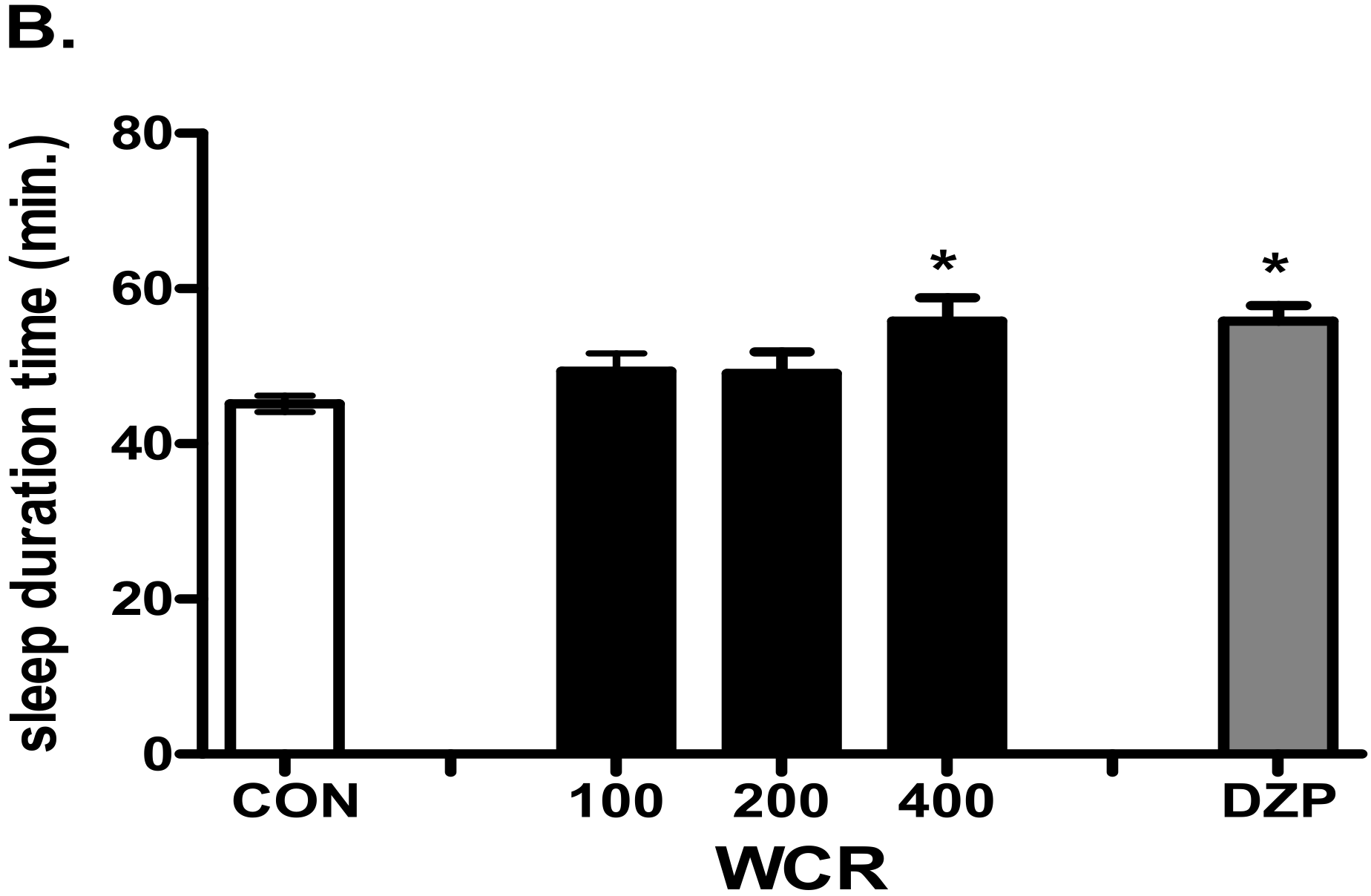
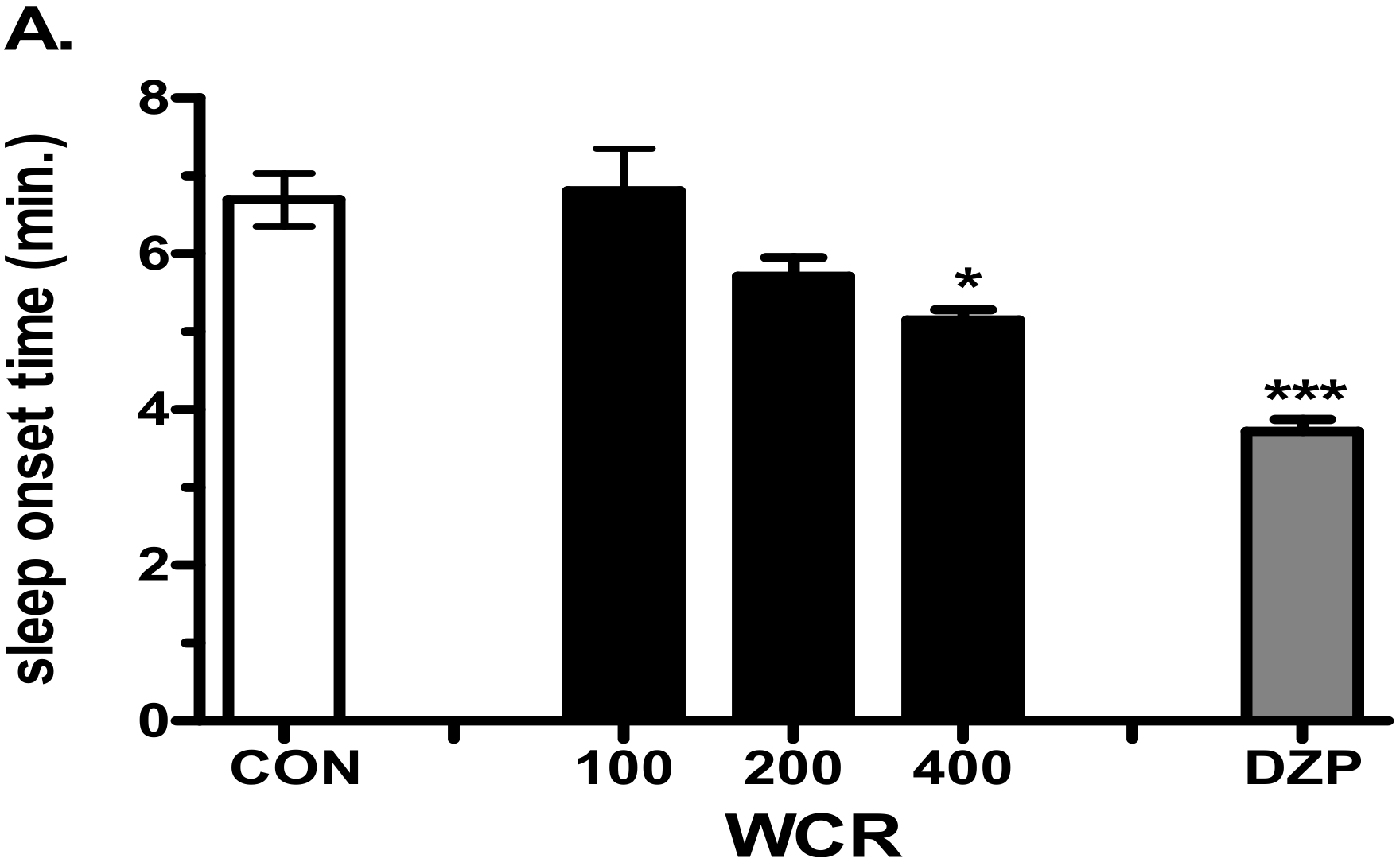
2.4. The Effect of WCR on Pentobarbital-Induced Sleep in Mice
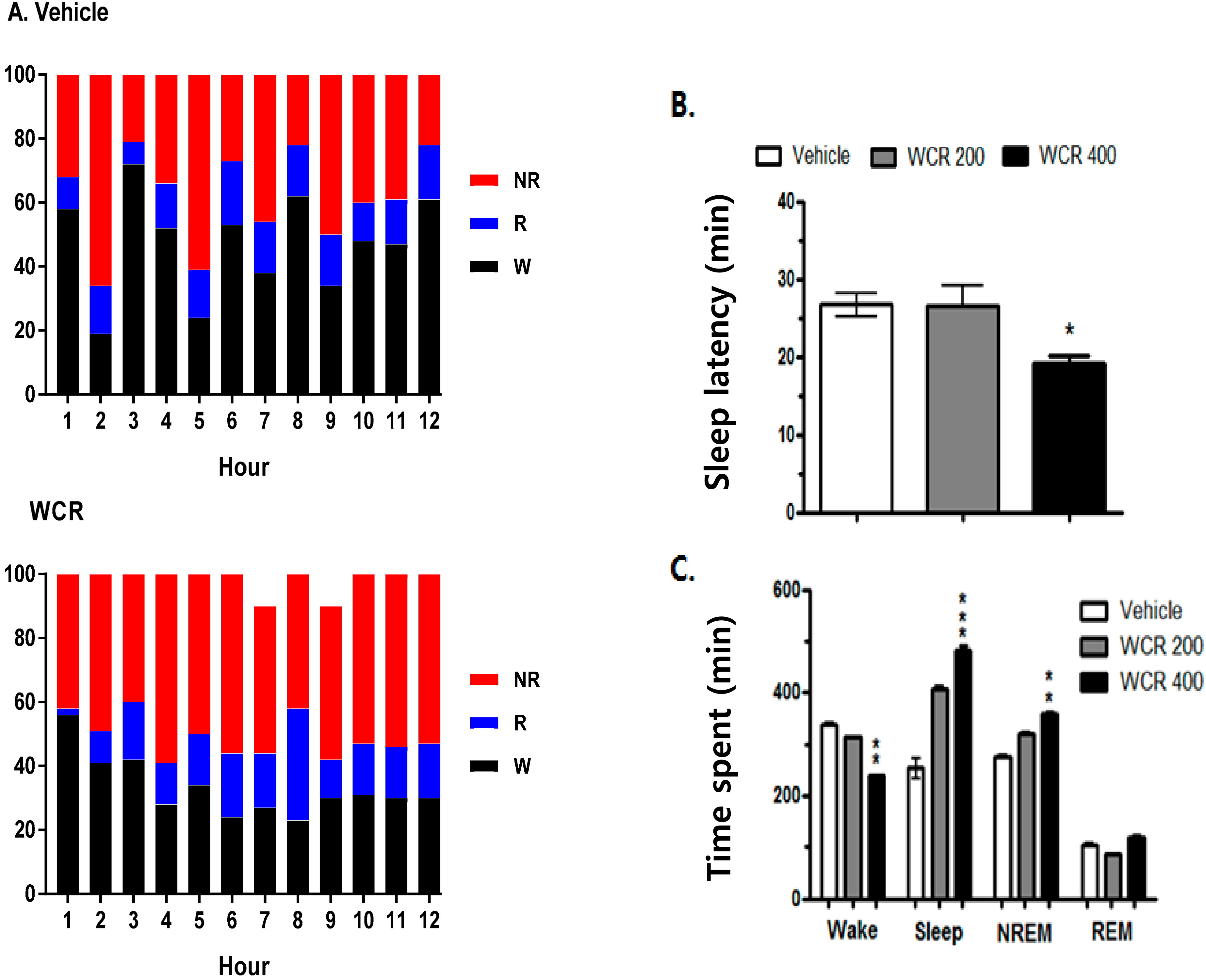
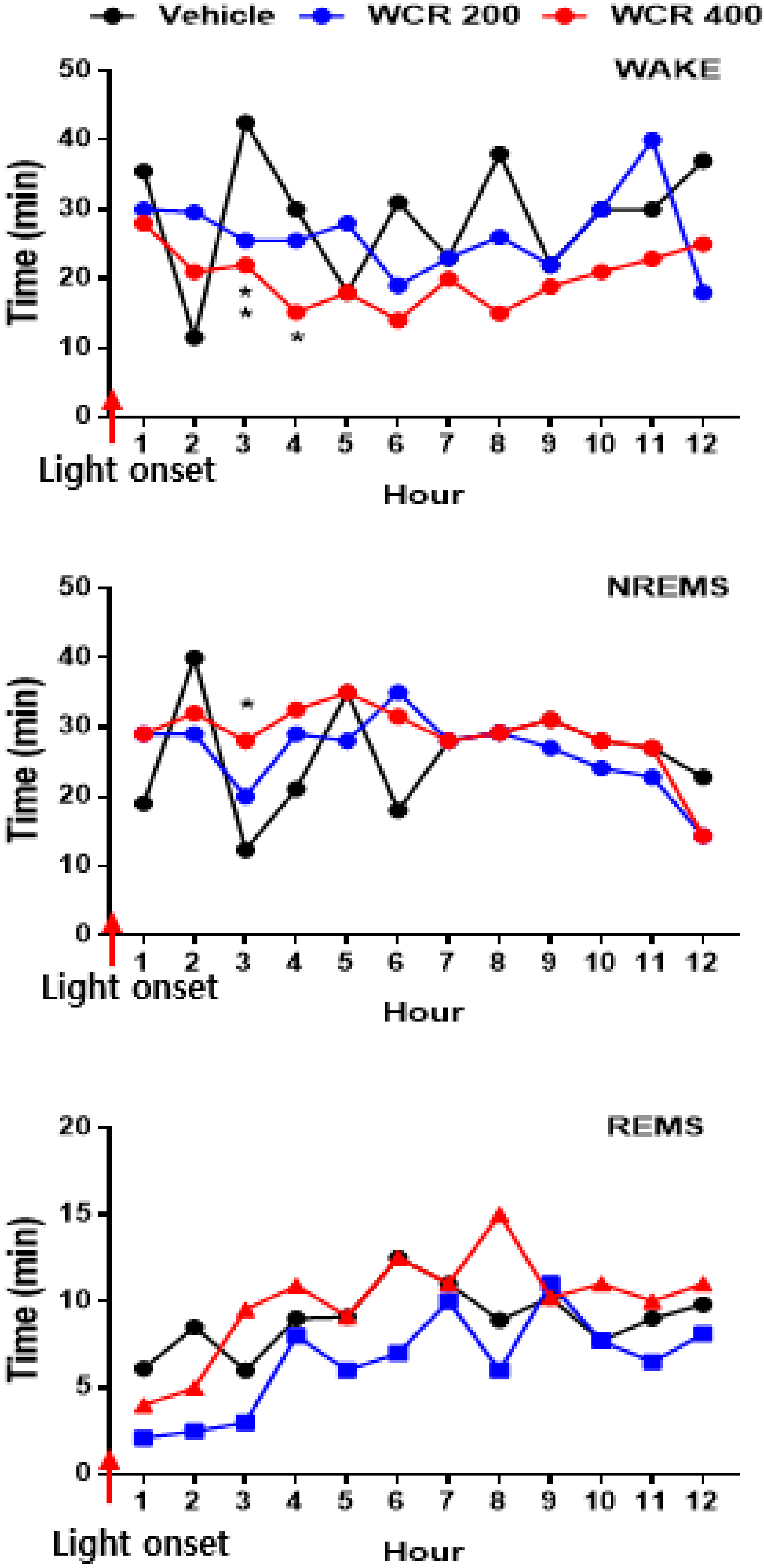
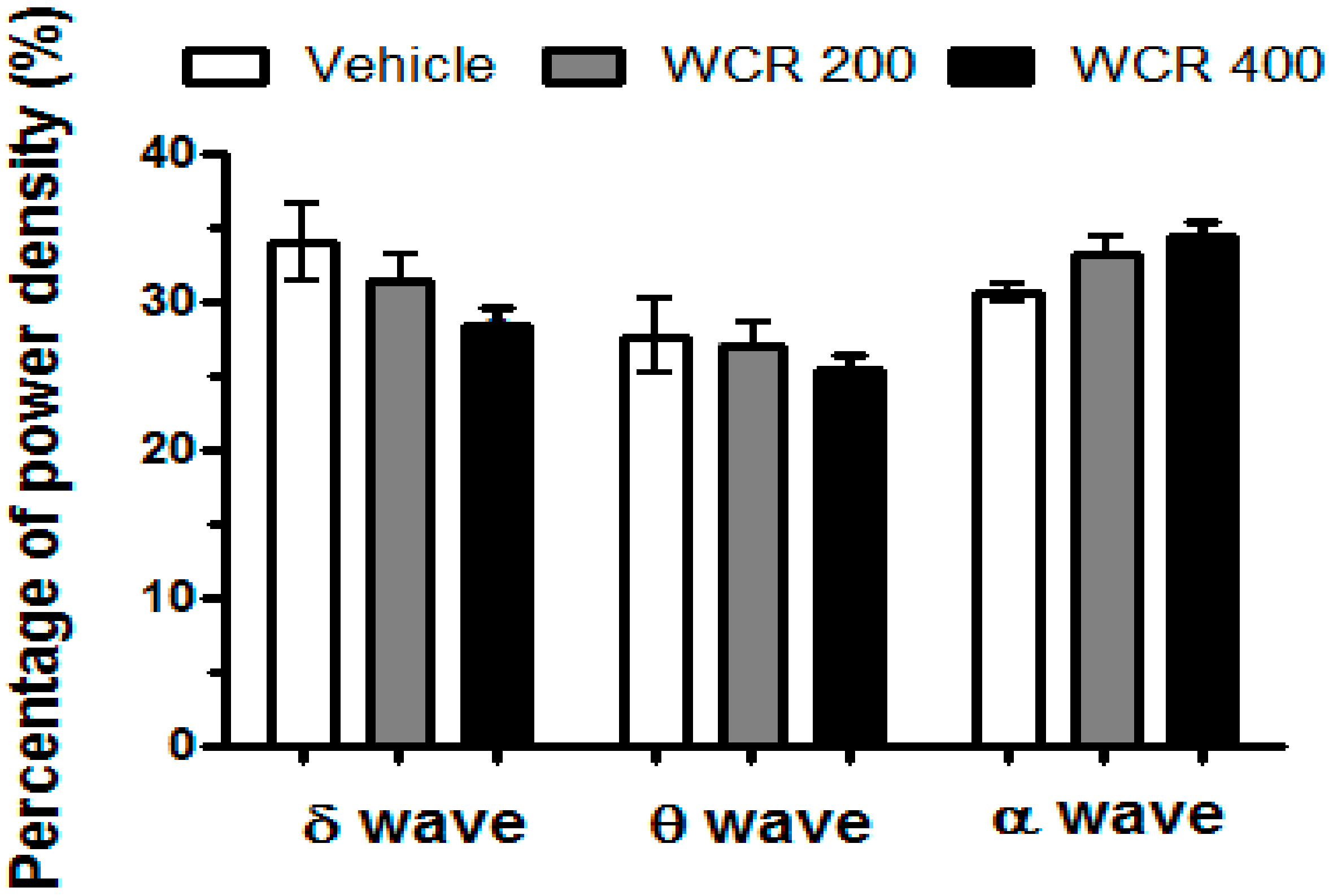
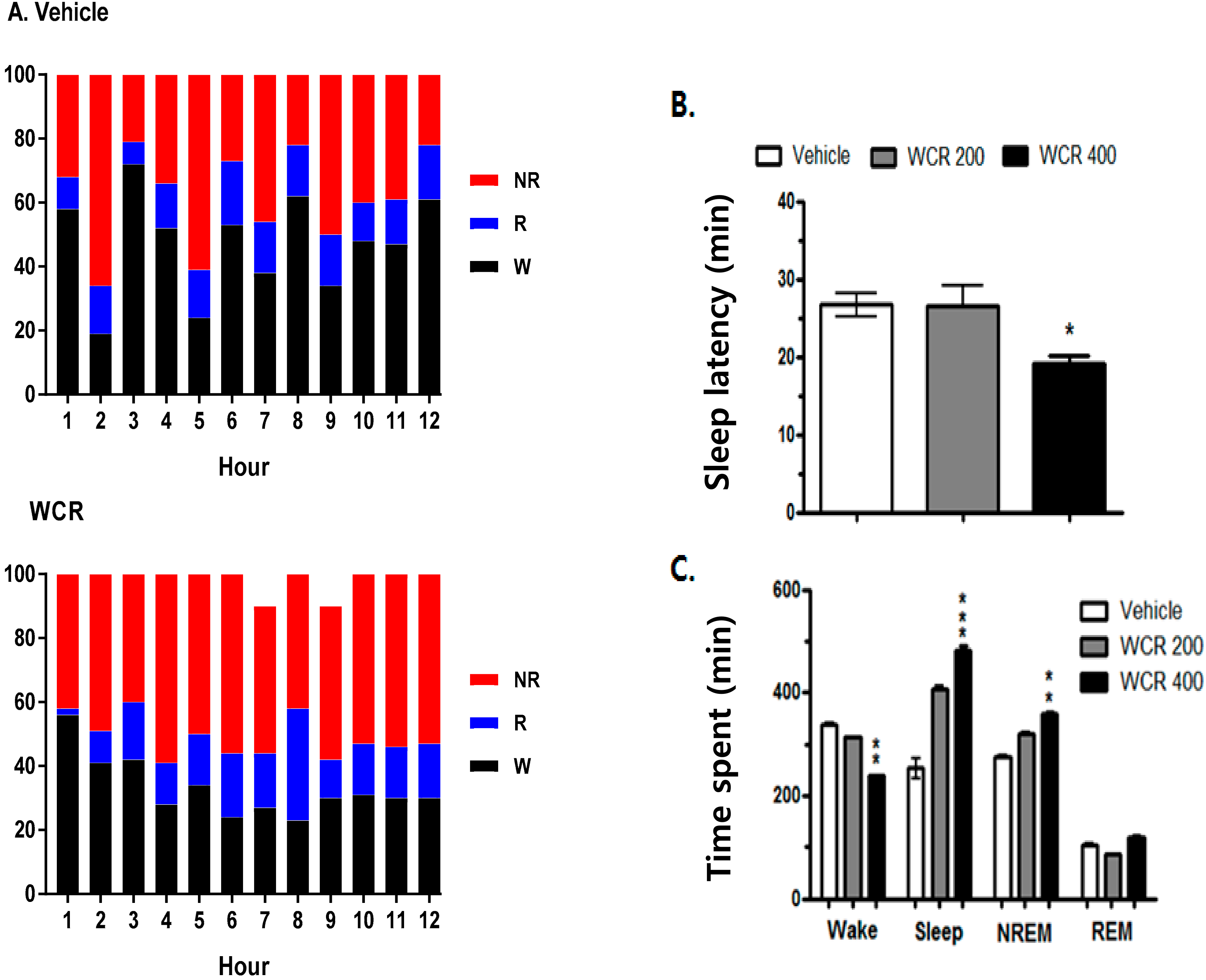
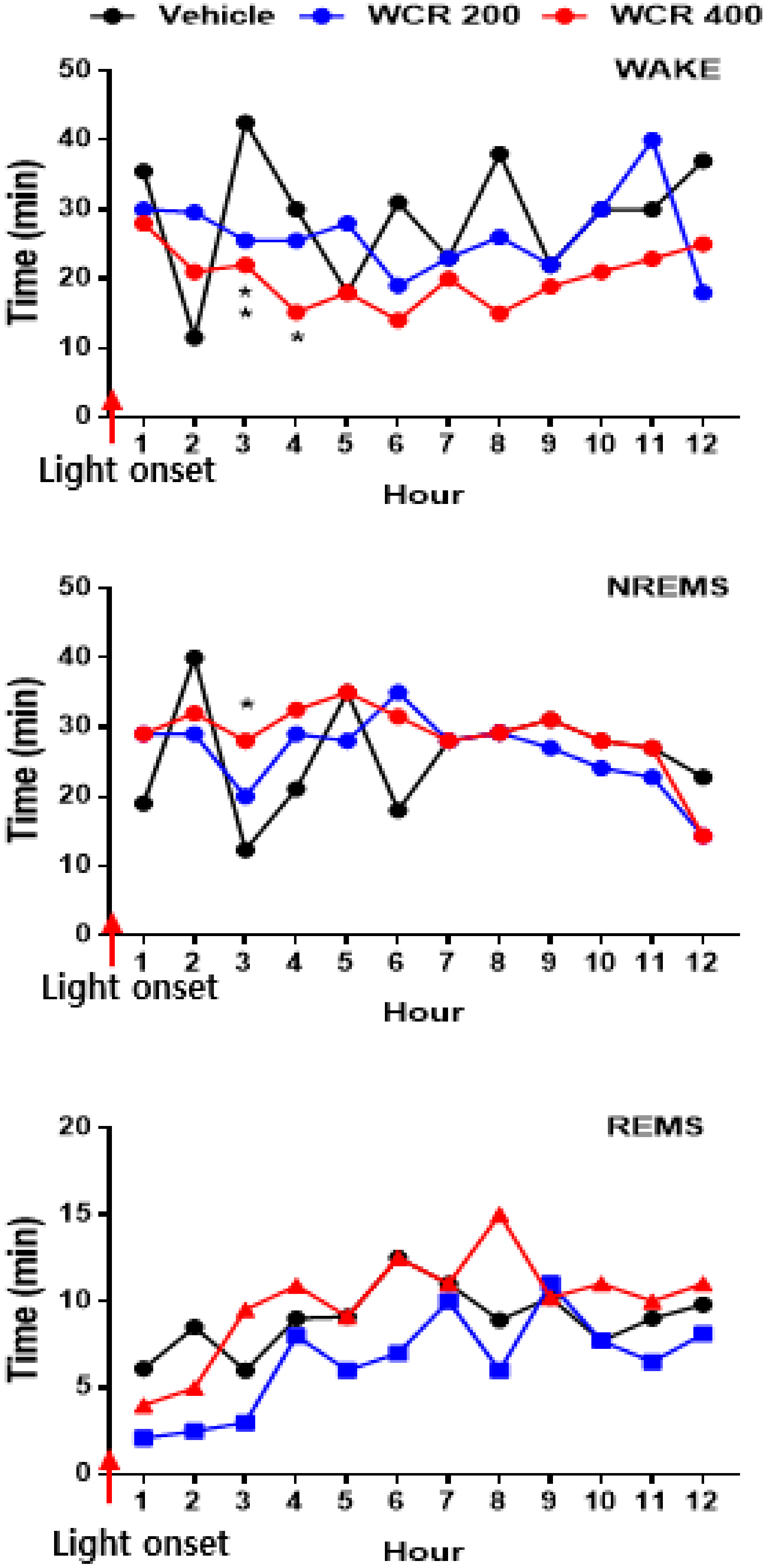
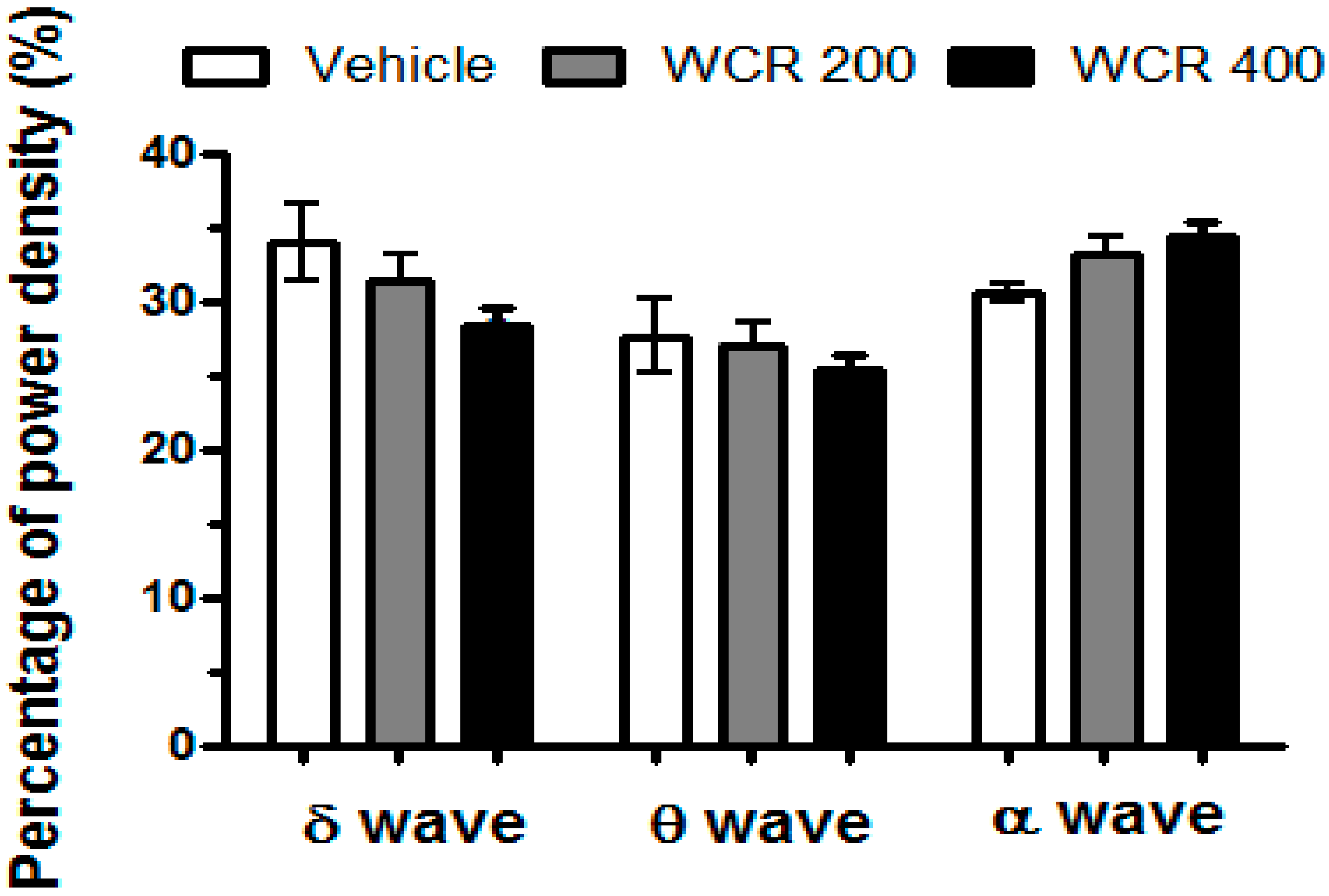
2.5. Effect of WCR on the EEG Sleep Architecture in Rats
3. Discussion
4. Materials and Methods
4.1. Materials Extracts
4.2. Animals
4.3. [3H]-Flumazenil Binding Assay
4.4. [3H]-Mesulergine Binding Assay
4.5. Pineal Gland Dissociation Culture
4.6. AANAT Assay
4.7. Measurement of Pentobarbital-Induced Anesthesia
4.8. EEG Surgery
4.9. Methodology of EEG and EMG Recording
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhaskar, S.; Hemavathy, D.; Prasad, S. Prevalence of chronic insomnia in adult patients and its correlation with medical comorbidities. J. Fam. Med. Prim. Care 2016, 5, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Attele, A.S.; Xie, J.T.; Yuan, C.S. Treatment of insomnia: An alternative approach. Altern. Med. Rev. 2000, 5, 249–259. [Google Scholar] [PubMed]
- Kroll, D.S.; Nieva, H.R.; Barsky, A.J.; Linder, J.A. Benzodiazepines are Prescribed More Frequently to Patients Already at Risk for Benzodiazepine-Related Adverse Events in Primary Care. J. Gen. Intern. Med. 2016, 31, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- Pariente, A.; de Gage, S.B.; Moore, N.; Begaud, B. The Benzodiazepine-Dementia Disorders Link: Current State of Knowledge. CNS Drugs 2016, 30, 1–7. [Google Scholar] [CrossRef]
- Borja, N.L.; Daniel, K.L. Ramelteon for the treatment of insomnia. Clin. Ther. 2006, 28, 1540–1555. [Google Scholar] [CrossRef]
- Ramar, K.; Olson, E.J. Management of common sleep disorders. Am. Fam. Physician 2013, 88, 231–238. [Google Scholar]
- Min, S.K. Korean traditional medicines as a source of novel drugs for neuropsychiatric disorders. Int. J. Neuropsychoph. 2006, 9, S67. [Google Scholar]
- Chung, I.W.; Kim, Y.S.; Ahn, J.S.; Lee, H.S.; Chen, G.; Manji, H.K.; Potter, W.Z.; Pickar, D. Pharmacologic profile of natural products used to treat psychotic illnesses. Psychopharmacol. Bull. 1995, 31, 139–145. [Google Scholar]
- Ernst, E. Herbal medications for common ailments in the elderly. Drugs Aging 1999, 15, 423–428. [Google Scholar] [CrossRef]
- Min, S.K.; Hong, C.H. Korean Traditional Medicines as Novel Drugs for Neuropsychiatric Disorders. Korean J. Psychopharmacol. 2007, 18, 5–17. [Google Scholar]
- Huang, F.; Xiong, Y.; Xu, L.; Ma, S.; Dou, C. Sedative and hypnotic activities of the ethanol fraction from Fructus Schisandrae in mice and rats. J. Ethnopharmacol. 2007, 110, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ma, H.; Eun, J.S.; Nam, S.Y.; Kim, Y.B.; Hong, J.T.; Lee, M.K.; Oh, K.W. Methanol extract of Longanae Arillus augments pentobarbital-induced sleep behaviors through the modification of GABAergic systems. J. Ethnopharmacol. 2009, 122, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.J.; Simpson, P.B. Methodological approaches for the study of GABA(A) receptor pharmacology and functional responses. Anal. Bioanal. Chem. 2003, 377, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Abourashed, E.A.; Koetter, U.; Brattstrom, A. In vitro binding experiments with a Valerian, hops and their fixed combination extract (Ze91019) to selected central nervous system receptors. Phytomedicine 2004, 11, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.M.; Shimizu, M.; Lee, C.J.; Han, D.S.; Jung, C.K.; Jo, J.H.; Kim, Y.M. Hypnotic effects and binding studies for GABA(A) and 5-HT(2C) receptors of traditional medicinal plants used in Asia for insomnia. J. Ethnopharmacol. 2010, 132, 225–232. [Google Scholar] [CrossRef]
- Monti, J.M. Serotonin control of sleep-wake behavior. Sleep Med. Rev. 2011, 15, 269–281. [Google Scholar] [CrossRef]
- Calabrese, E.J. An assessment of anxiolytic drug screening tests: Hormetic dose responses predominate. Crit. Rev. Toxicol. 2008, 38, 489–542. [Google Scholar] [CrossRef]
- Feng, J.; Cai, X.; Zhao, J.; Yan, Z. Serotonin receptors modulate GABA(A) receptor channels through activation of anchored protein kinase C in prefrontal cortical neurons. J. Neurosci. 2001, 21, 6502–6511. [Google Scholar] [CrossRef] [Green Version]
- Huidobro-Toro, J.P.; Valenzuela, C.F.; Harris, R.A. Modulation of GABAA receptor function by G protein-coupled 5-HT2C receptors. Neuropharmacology 1996, 35, 1355–1363. [Google Scholar] [CrossRef]
- Itoh, M.T.; Ishizuka, B.; Kuribayashi, Y.; Amemiya, A.; Sumi, Y. Melatonin, its precursors, and synthesizing enzyme activities in the human ovary. Mol. Hum. Reprod. 1999, 5, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Borjigin, J. N-acetyltransferase is not the rate-limiting enzyme of melatonin synthesis at night. J. Pineal Res. 2005, 39, 91–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altun, A.; Ugur-Altun, B. Melatonin: Therapeutic and clinical utilization. Int. J. Clin. Pr. 2007, 61, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Pandi-Perumal, S.R.; Trahkt, I.; Spence, D.W.; Poeggeler, B.; Hardeland, R.; Cardinali, D.P. Melatonin and melatonergic drugs on sleep: Possible mechanisms of action. Int. J. Neurosci. 2009, 119, 821–846. [Google Scholar] [CrossRef] [PubMed]
- Fornaro, M.; Prestia, D.; Colicchio, S.; Perugi, G. A systematic, updated review on the antidepressant agomelatine focusing on its melatonergic modulation. Curr. Neuropharmacol. 2010, 8, 287–304. [Google Scholar] [CrossRef] [Green Version]
- Turek, F.W.; Gillette, M.U. Melatonin, sleep, and circadian rhythms: Rationale for development of specific melatonin agonists. Sleep Med. 2004, 5, 523–532. [Google Scholar] [CrossRef]
- Joung, H.Y.; Kang, Y.M.; Lee, B.J.; Chung, S.Y.; Kim, K.S.; Shim, I. Sedative-Hypnotic and Receptor Binding Studies of Fermented Marine Organisms. Biomol. Ther. (Seoul) 2015, 23, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Lancel, M. Role of GABAA receptors in the regulation of sleep: Initial sleep responses to peripherally administered modulators and agonists. Sleep 1999, 22, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.K.; Dhir, A. On the mechanism of antidepressant-like action of berberine chloride. Eur. J. Pharm. 2008, 589, 163–172. [Google Scholar] [CrossRef]
- Meng, F.C.; Wu, Z.F.; Yin, Z.Q.; Lin, L.G.; Wang, R.; Zhang, Q.W. Coptidis rhizoma and its main bioactive components: Recent advances in chemical investigation, quality evaluation and pharmacological activity. Chin. Med. 2018, 13, 13. [Google Scholar] [CrossRef]
- Wang, N.; Tan, H.Y.; Li, L.; Yuen, M.F.; Feng, Y. Berberine and Coptidis Rhizoma as potential anticancer agents: Recent updates and future perspectives. J. Ethnopharmacol. 2015, 176, 35–48. [Google Scholar] [CrossRef]
- Kong, J.C.; Lee, M.S.; Shin, B.C. Randomized clinical trials on acupuncture in korean literature: A systematic review. Evid. Based Complement. Altern. Med. 2009, 6, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, L.P.; Sochilina, E.E.; Faddeeva, M.D.; Iagodina, O.V. Effect of some isoquinoline alkaloids on enzymatic activity of acetylcholinesterase and monoamine oxidase. Ukr. Biokhimichnyi Zhurnal 2005, 77, 147–153. [Google Scholar]
- Fan, J.; Li, B.; Ge, T.; Zhang, Z.; Lv, J.; Zhao, J.; Wang, P.; Liu, W.; Wang, X.; Mlyniec, K. Berberine produces antidepressant-like effects in ovariectomized mice. Sci. Rep. 2017, 7, 1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dmitrzak-Weglarz, M.; Banach, E.; Bilska, K.; Narozna, B.; Szczepankiewicz, A.; Reszka, E.; Jablonska, E.; Kapelski, P.; Skibinska, M.; Pawlak, J. Molecular Regulation of the Melatonin Biosynthesis Pathway in Unipolar and Bipolar Depression. Front. Pharm. 2021, 12, 666541. [Google Scholar] [CrossRef] [PubMed]
- De Berardis, D.; Marini, S.; Fornaro, M.; Srinivasan, V.; Iasevoli, F.; Tomasetti, C.; Valchera, A.; Perna, G.; Quera-Salva, M.A.; Martinotti, G. The melatonergic system in mood and anxiety disorders and the role of agomelatine: Implications for clinical practice. Int. J. Mol. Sci. 2013, 14, 12458–12483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonneaux, V.; Ribelayga, C. Generation of the melatonin endocrine message in mammals: A review of the complex regulation of melatonin synthesis by norepinephrine, peptides, and other pineal transmitters. Pharm. Rev. 2003, 55, 325–395. [Google Scholar] [CrossRef]
- Hartley, S.W.; Coon, S.L.; Savastano, L.E.; Mullikin, J.C.; Program, N.C.S.; Fu, C.; Klein, D.C. Neurotranscriptomics: The Effects of Neonatal Stimulus Deprivation on the Rat Pineal Transcriptome. PLoS ONE 2015, 10, e0137548. [Google Scholar] [CrossRef] [Green Version]
- Klein, D.C.; Coon, S.L.; Roseboom, P.H.; Weller, J.L.; Bernard, M.; Gastel, J.A.; Zatz, M.; Iuvone, P.M.; Rodriguez, I.R.; Begay, V. The melatonin rhythm-generating enzyme: Molecular regulation of serotonin N-acetyltransferase in the pineal gland. Recent Prog. Horm. Res. 1997, 52, 307–357. [Google Scholar]
- Yu, H.; Benitez, S.G.; Jung, S.R.; Farias Altamirano, L.E.; Kruse, M.; Seo, J.B.; Koh, D.S.; Munoz, E.M.; Hille, B. GABAergic signaling in the rat pineal gland. J. Pineal Res. 2016, 61, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Rosenstein, R.E.; Chuluyan, H.E.; Diaz, M.C.; Cardinali, D.P. GABA as a presumptive paracrine signal in the pineal gland. Evidence on an intrapineal GABAergic system. Brain Res. Bull. 1990, 25, 339–344. [Google Scholar] [CrossRef]
- Risa, J.; Risa, A.; Adsersen, A.; Gauguin, B.; Stafford, G.I.; van Staden, J.; Jager, A.K. Screening of plants used in southern Africa for epilepsy and convulsions in the GABA(A)-benzodiazepine receptor assay. J. Ethnopharmacol. 2004, 93, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Wang, C.; Yang, W. Role of berberine in Alzheimer’s disease. Neuropsychiatr. Dis. Treat. 2016, 12, 2509–2520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, S.; Hobson, J.A. The rat as an experimental model for sleep neurophysiology. Behav. Neurosci. 2000, 114, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- Borbely, A.A.; Baumann, F.; Brandeis, D.; Strauch, I.; Lehmann, D. Sleep deprivation: Effect on sleep stages and EEG power density in man. Electroencephalogr. Clin. Neurophysiol. 1981, 51, 483–495. [Google Scholar] [CrossRef]
- Borbely, A.A. A two process model of sleep regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar]
- Dijk, D.J.; Brunner, D.P.; Borbely, A.A. Time course of EEG power density during long sleep in humans. Am. J. Physiol. 1990, 258, R650–R661. [Google Scholar] [CrossRef]
- Kim, B.; Hwang, E.; Strecker, R.E.; Choi, J.H.; Kim, Y. Differential modulation of NREM sleep regulation and EEG topography by chronic sleep restriction in mice. Sci. Rep. 2020, 10, 18. [Google Scholar]
- Bernard, M.; Klein, D.C.; Zatz, M. Chick pineal clock regulates serotonin N-acetyltransferase mRNA rhythm in culture. Proc. Natl. Acad. Sci. USA 1997, 94, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Chae, H.D.; Park, T.J.; Lee, Y.K.; Lee, T.G.; Kim, K.T. Rapid and simple measurement of serotonin N-acetyltransferase activity by liquid biphasic diffusion assay. Neurochem. Int. 1999, 35, 447–451. [Google Scholar] [CrossRef]
- Rakhshandah, H.; Shakeri, M.T.; Ghasemzadeh, M.R. Comparative hypnotic effect of Rosa damascena fractions and diazepam in mice. Iran. J. Pharm. Res. 2007, 6, 193–197. [Google Scholar]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, Compact 6th ed.; Academic: London, UK, 2009. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Displacement (%) of [3H]-Flumazenil Binding at Different Concentrations (mg/mL) | |||||
|---|---|---|---|---|---|---|
| Scientific Name | 0.01 | 0.1 | 1 | 5 | 10 | 20 |
| Lycii Fructus | −2.1 ± 0.7 | −2.1 ± 4.7 | −14.6 ± 1.4 | −8.6 ± 0.6 | −1.5 ± 3.7 | 10.5 ± 4.6 |
| Angelicae sinensis Radix | 3.2 ± 1.7 | 7.5 ± 2.7 | −0.8 ± 4.2 | 5.28 ± 2.3 | 4.3 ± 1.2 | 14.5 ± 1.5 |
| Coptidis Rhizoma | −6.5 ± 6.2 | −4.4 ± 1.2 | 0.4 ± 1.6 | 64.2 ± 1.0 | 84.8 ± 0.6 | 82.4 ± 0.8 |
| Bupleuri Radix | 25.0 ± 1.5 | 26.0 ± 3.8 | 31.5 ± 2.1 | 55.3 ± 0.9 | 62.8 ± 0.5 | 78.2 ± 0.6 |
| Polygonum multiflorum Thunberg | −16.8 ± 0.5 | −18.9 ± 0.8 | −17.9 ± 1.5 | −10.0 ± 1.5 | 0.1 ± 2.5 | 22.2 ± 1.9 |
| Species | Displacement (%) of [3H]-Mesulergine Binding at Different Plant Concentrations (mg/mL) | |||||
|---|---|---|---|---|---|---|
| Scientific Name | 0.01 | 0.1 | 1 | 5 | 10 | 20 |
| Lycii Fructus | 22.4 ± 0.2 | 15.2 ± 3.1 | 13.6 ± 5.4 | 12.2 ± 4.0 | 9.3 ± 4.1 | 13.4 ± 0.8 |
| Angelicae sinensis Radix | 15.9 ± 4.5 | 17.8 ± 3.1 | 8.6 ± 5.4 | −2.0 ± 3.0 | 5.0 ± 11.3 | 2.2 ± 12.1 |
| Coptidis Rhizoma | 31.3 ± 3.3 | 22.8 ± 0.3 | 62.7 ± 0.2 | 87.9 ± 1.4 | 95.5 ± 0.4 | 99.1 ± 0.2 |
| Bupleuri Radix | 15.5 ± 4.7 | 12.8 ± 1.5 | 18.7 ± 4.7 | 19.6 ± 0.4 | 0.4 ± 3.1 | −5.7 ± 1.8 |
| Polygonum multiflorum Thunberg | 16.3 ± 2.0 | 15.6 ± 2.6 | 14.2 ± 0.9 | 36.8 ± 1.3 | 59.2 ± 2.3 | 66.1 ± 0.4 |
| Species | Plant Part Analyzed | Traditional Usage for Neuropsychology | |
|---|---|---|---|
| Name | Scientific Name | ||
| Gouqi | Lycii Fructus | Fruit | Inhibition of CNS, sedation |
| Danghui | Angelicae sinensis Radix | Root | Relaxation, sedation |
| Barberry root | Coptidis Rhizoma | Root | Relaxation, treatment of anxiety symptoms |
| Chai Hu | Bupleuri Radix | Root | Sedation, analgesic activity |
| Huang bai | Polygonum multiflorumThunberg | Bark | Anti-inflammation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joung, H.-Y.; Ye, M.; Lee, M.; Hong, Y.; Kim, M.; Kim, K.S.; Shim, I. Sedative–Hypnotic Activity of the Water Extracts of Coptidis Rhizoma in Rodents. Clocks & Sleep 2022, 4, 145-159. https://doi.org/10.3390/clockssleep4010014
Joung H-Y, Ye M, Lee M, Hong Y, Kim M, Kim KS, Shim I. Sedative–Hypnotic Activity of the Water Extracts of Coptidis Rhizoma in Rodents. Clocks & Sleep. 2022; 4(1):145-159. https://doi.org/10.3390/clockssleep4010014
Chicago/Turabian StyleJoung, Hye-Young, Minsook Ye, Miyoung Lee, Yunki Hong, Minji Kim, Kyung Soo Kim, and Insop Shim. 2022. "Sedative–Hypnotic Activity of the Water Extracts of Coptidis Rhizoma in Rodents" Clocks & Sleep 4, no. 1: 145-159. https://doi.org/10.3390/clockssleep4010014
APA StyleJoung, H.-Y., Ye, M., Lee, M., Hong, Y., Kim, M., Kim, K. S., & Shim, I. (2022). Sedative–Hypnotic Activity of the Water Extracts of Coptidis Rhizoma in Rodents. Clocks & Sleep, 4(1), 145-159. https://doi.org/10.3390/clockssleep4010014