Light Modulation of Human Clocks, Wake, and Sleep




Abstract
1. Light Modulation of Non-Image Forming Functions
1.1. The Eye, Vision, and Non-Image Forming Functions
1.2. Acute and Circadian Effects of Light
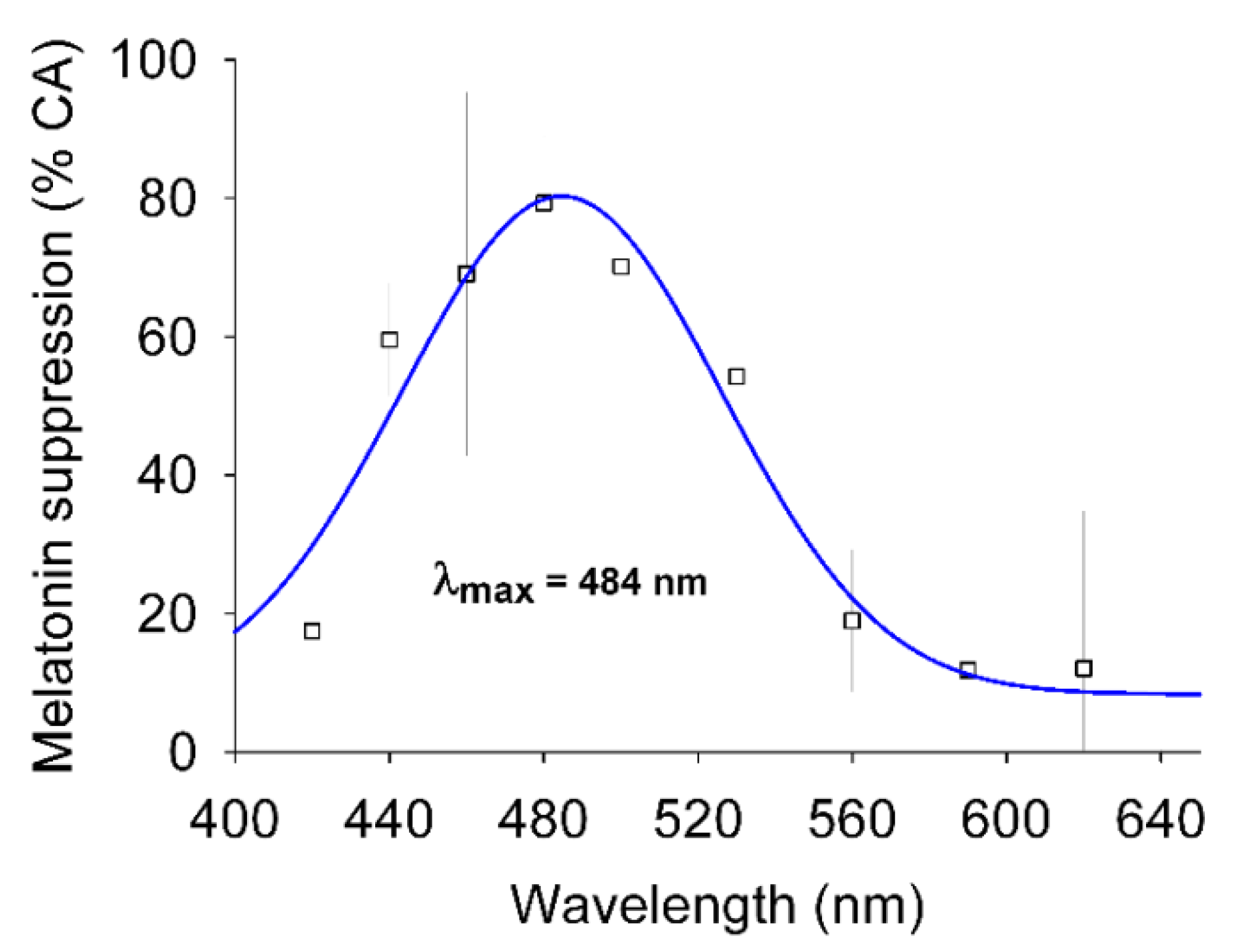
1.2.1. Differences in Spectral Sensitivity to Light
1.2.2. Impact of Duration of Light Exposure
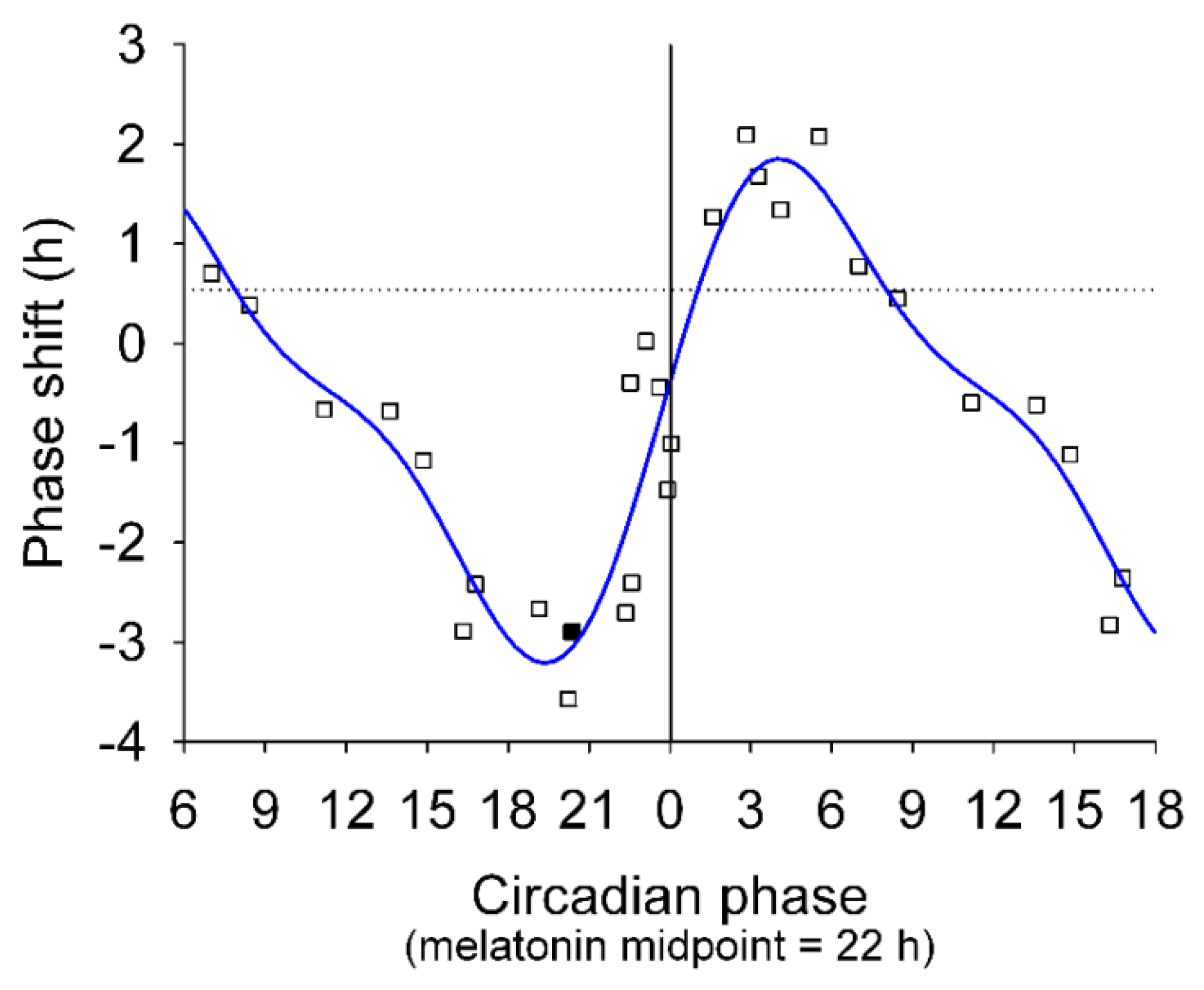
1.2.3. Effects of Timing of Light Exposure
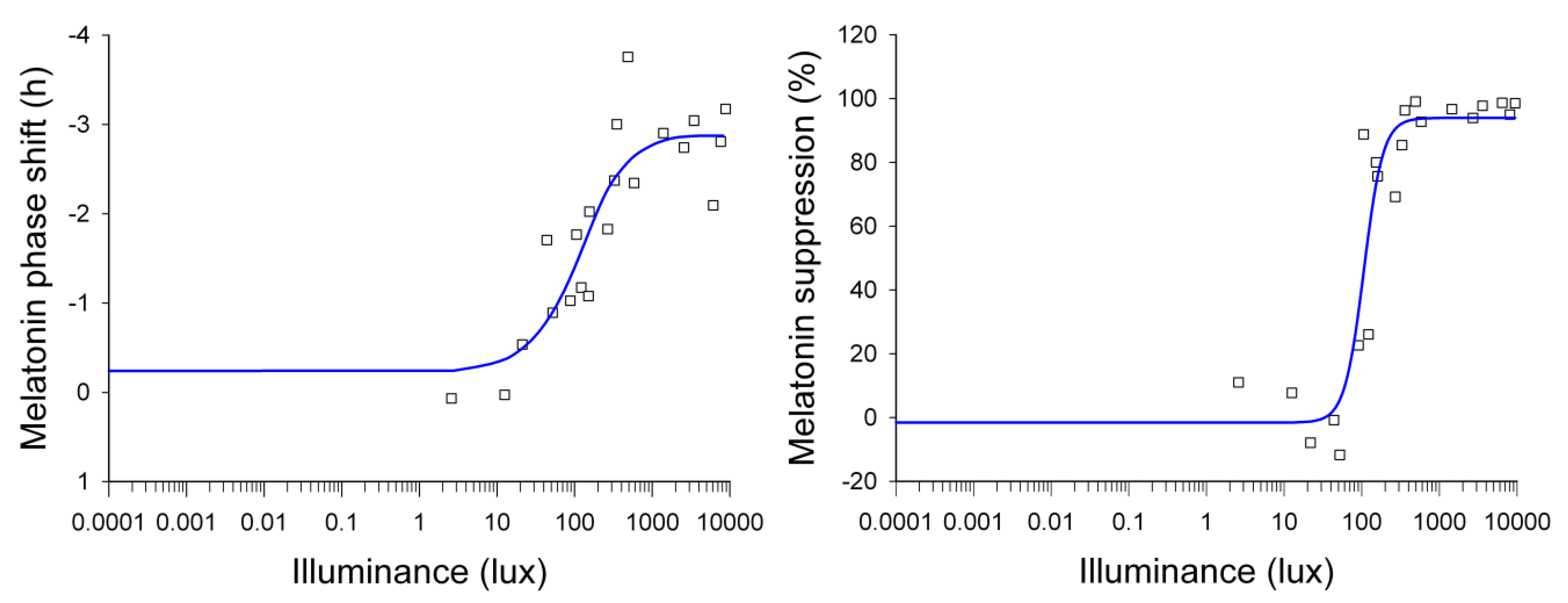
1.2.4. Effects of Light Intensity
1.2.5. Effects of Temporal Patterns of Light Exposure
1.2.6. Impact of Spatial Distribution of Light Exposure
1.2.7. Effects of Prior Light History
2. Impact of Light on Cortical Activity during Wakefulness
3. Impact of Light on Sleep
3.1. Impact of Timing and Intensity of Light Exposure on Sleep
3.2. Impact of Spectral Composition of Light Exposure on Sleep
4. Conclusions
5. Outlook and Future Directions
Funding
Conflicts of Interest
References
- Gooley, J.J.; Lu, J.; Chou, T.C.; Scammell, T.E.; Saper, C.B. Melanopsin in cells of origin of the retinohypothalamic tract. Nat. Neurosci. 2001, 4, 1165. [Google Scholar] [CrossRef] [PubMed]
- Hannibal, J.; Fahrenkrug, J. Melanopsin: A novel photopigment involved in the photoentrainment of the brain’s biological clock? Ann. Med. 2002, 34, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Buijs, R.M.; Scheer, F.A.; Kreier, F.; Yi, C.; Bos, N.; Goncharuk, V.D.; Kalsbeek, A. Organization of circadian functions: Interaction with the body. Prog. Brain Res. 2006, 153, 341–360. [Google Scholar] [PubMed]
- Hannibal, J.; Kankipati, L.; Strang, C.E.; Peterson, B.B.; Dacey, D.; Gamlin, P.D. Central projections of intrinsically photosensitive retinal ganglion cells in the macaque monkey. J. Comp. Neurol. 2014, 522, 2231–2248. [Google Scholar] [CrossRef]
- Hattar, S.; Kumar, M.; Park, A.; Tong, P.; Tung, J.; Yau, K.-W.; Berson, D.M. Central projections of melanopsin-expressing retinal ganglion cells in the mouse. J. Comp. Neurol. 2006, 497, 326–349. [Google Scholar] [CrossRef]
- Khalsa, S.B.S.; Jewett, M.E.; Cajochen, C.; Czeisler, C.A. A phase response curve to single bright light pulses in human subjects. J. Physiol. 2003, 549, 945–952. [Google Scholar] [CrossRef]
- Mure, L.S.; Cornut, P.-L.; Rieux, C.; Drouyer, E.; Denis, P.; Gronfier, C.; Cooper, H.M. Melanopsin bistability: A fly’s eye technology in the human retina. PLoS ONE 2009, 4, e5991. [Google Scholar] [CrossRef]
- Cajochen, C.; Münch, M.; Kobialka, S.; Kräuchi, K.; Steiner, R.; Oelhafen, P.; Orgül, S.; Wirz-Justice, A. High sensitivity of human melatonin, alertness, thermoregulation, and heart rate to short wavelength light. J. Clin. Endocrinol. Metab. 2005, 90, 1311–1316. [Google Scholar] [CrossRef]
- Rüger, M.; Gordijn, M.C.M.; Beersma, D.G.M.; de Vries, B.; Daan, S. Time-of-day-dependent effects of bright light exposure on human psychophysiology: Comparison of daytime and nighttime exposure. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2005, 290, R1413–R1420. [Google Scholar] [CrossRef]
- Dijk, D.-J.; Cajochen, C.; Borbély, A.A. Effect of a single 3-hour exposure to bright light on core body temperature and sleep in humans. Neurosci. Lett. 1991, 121, 59–62. [Google Scholar] [CrossRef]
- Rahman, S.A.; Flynn-Evans, E.E.; Aeschbach, D.; Brainard, G.C.; Czeisler, C.A.; Lockley, S.W. Diurnal spectral sensitivity of the acute alerting effects of light. Sleep 2014, 37, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Badia, P.; Myers, B.; Boecker, M.; Culpepper, J.; Harsh, J.R. Bright light effects on body Temperature, alertness, EEG and behavior. Physiol. Behav. 1991, 50, 583–588. [Google Scholar] [CrossRef]
- Münch, M.; Léon, L.; Crippa, S.V.; Kawasaki, A. Circadian and wake-dependent effects on the pupil light reflex in response to narrow-bandwidth light pulses. Investig. Opthalmol. Vis. Sci. 2012, 53, 4546. [Google Scholar] [CrossRef] [PubMed]
- Zele, A.J.; Adhikari, P.; Feigl, B.; Cao, D. Cone and melanopsin contributions to human brightness estimation. J. Opt. Soc. Am. A 2018, 35, B19–B25. [Google Scholar] [CrossRef]
- Zele, A.J.; Feigl, B.; Adhikari, P.; Maynard, M.L.; Cao, D. Melanopsin photoreception contributes to human visual detection, temporal and colour processing. Sci. Rep. 2018, 8, 3842. [Google Scholar] [CrossRef]
- Spitschan, M.; Datta, R.; Stern, A.M.; Brainard, D.H.; Aguirre, G.K. Human visual cortex responses to rapid cone and melanopsin-directed flicker. J. Neurosci. 2016, 36, 1471–1482. [Google Scholar] [CrossRef] [PubMed]
- Spitschan, M.; Bock, A.S.; Ryan, J.; Frazzetta, G.; Brainard, D.H.; Aguirre, G.K. The human visual cortex response to melanopsin-directed stimulation is accompanied by a distinct perceptual experience. Proc. Natl. Acad. Sci. USA 2017, 114, 12291–12296. [Google Scholar] [CrossRef]
- Zaidi, F.H.; Hull, J.T.; Peirson, S.N.; Wulff, K.; Aeschbach, D.; Gooley, J.J.; Brainard, G.C.; Gregory-Evans, K.; Rizzo, J.F.; Czeisler, C.A.; et al. Short-wavelength light sensitivity of circadian, pupillary, and visual awareness in humans lacking an outer retina. Curr. Biol. 2007, 17, 2122–2128. [Google Scholar] [CrossRef]
- Brown, T.M.; Tsujimura, S.; Allen, A.E.; Wynne, J.; Bedford, R.; Vickery, G.; Vugler, A.; Lucas, R.J. Melanopsin-based brightness discrimination in mice and humans. Curr. Biol. 2012, 22, 1134–1141. [Google Scholar] [CrossRef]
- Hannibal, J.; Christiansen, A.T.; Heegaard, S.; Fahrenkrug, J.; Kiilgaard, J.F. Melanopsin expressing human retinal ganglion cells: Subtypes, distribution, and intraretinal connectivity. J. Comp. Neurol. 2017, 525, 1934–1961. [Google Scholar] [CrossRef]
- Schmidt, T.M.; Chen, S.-K.; Hattar, S. Intrinsically photosensitive retinal ganglion cells: Many subtypes, diverse functions. Trends Neurosci. 2011, 34, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.Y. CIRCADIAN RHYTHMS: Basic neurobiology and clinical applications. Annu. Rev. Med. 1997, 48, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, D.C.; Fogerson, P.M.; Lazzerini Ospri, L.; Thomsen, M.B.; Layne, R.M.; Severin, D.; Zhan, J.; Singer, J.H.; Kirkwood, A.; Zhao, H.; et al. Light affects mood and learning through distinct retina-brain pathways. Cell 2018, 175, 71–84. [Google Scholar] [CrossRef]
- Chellappa, S.L.; Lasauskaite, R.; Cajochen, C. In a heartbeat: Light and cardiovascular physiology. Front. Neurol. 2017, 8, 541. [Google Scholar] [CrossRef]
- Chellappa, S.L.; Gordijn, M.C.M.; Cajochen, C. Can light make us bright? Effects of light on cognition and sleep. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2011; Volume 190, pp. 119–133. [Google Scholar]
- Chang, A.-M.; Scheer, F.A.J.L.; Czeisler, C.A.; Aeschbach, D. Direct effects of light on alertness, vigilance, and the waking electroencephalogram in humans depend on prior light history. Sleep 2013, 36, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Brainard, G.C.; Hanifin, J.P.; Greeson, J.M.; Byrne, B.; Glickman, G.; Gerner, E.; Rollag, M.D. Action spectrum for melatonin regulation in humans: Evidence for a novel circadian photoreceptor. J. Neurosci. 2001, 21, 6405–6412. [Google Scholar] [CrossRef] [PubMed]
- Thapan, K.; Arendt, J.; Skene, D.J. An action spectrum for melatonin suppression: Evidence for a novel non-rod, non-cone photoreceptor system in humans. J. Physiol. 2001, 535, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Najjar, R.P.; Chiquet, C.; Teikari, P.; Cornut, P.-L.; Claustrat, B.; Denis, P.; Cooper, H.M.; Gronfier, C. Aging of non-visual spectral sensitivity to light in humans: Compensatory mechanisms? PLoS ONE 2014, 9, e85837. [Google Scholar] [CrossRef] [PubMed]
- Lockley, S.W.; Brainard, G.C.; Czeisler, C.A. High sensitivity of the human circadian melatonin rhythm to resetting by short wavelength light. J. Clin. Endocrinol. Metab. 2003, 88, 4502–4505. [Google Scholar] [CrossRef]
- Revell, V.L.; Arendt, J.; Terman, M.; Skene, D.J. Short-wavelength sensitivity of the human circadian system to phase-advancing light. J. Biol. Rhythms 2005, 20, 270–272. [Google Scholar] [CrossRef]
- Chang, A.-M.; Santhi, N.; St Hilaire, M.; Gronfier, C.; Bradstreet, D.S.; Duffy, J.F.; Lockley, S.W.; Kronauer, R.E.; Czeisler, C.A. Human responses to bright light of different durations: Light DRC in humans. J. Physiol. 2012, 590, 3103–3112. [Google Scholar] [CrossRef] [PubMed]
- Dewan, K.; Benloucif, S.; Reid, K.; Wolfe, L.F.; Zee, P.C. Light-induced changes of the circadian clock of humans: Increasing duration is more effective than increasing light intensity. Sleep 2011, 34, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Prayag, A.S.; Jost, S.; Avouac, P.; Dumortier, D.; Gronfier, C. Dynamics of non-visual responses in humans: As fast as lightning? Front. Neurosci. 2019, 13, 126. [Google Scholar] [CrossRef]
- Rüger, M.; St Hilaire, M.A.; Brainard, G.C.; Khalsa, S.-B.S.; Kronauer, R.E.; Czeisler, C.A.; Lockley, S.W. Human phase response curve to a single 6.5 h pulse of short-wavelength light: Blue light phase response curve. J. Physiol. 2013, 591, 353–363. [Google Scholar] [CrossRef] [PubMed]
- St Hilaire, M.A.; Gooley, J.J.; Khalsa, S.B.S.; Kronauer, R.E.; Czeisler, C.A.; Lockley, S.W. Human phase response curve to a 1 h pulse of bright white light: PRC to 1 h light pulses in humans. J. Physiol. 2012, 590, 3035–3045. [Google Scholar] [CrossRef] [PubMed]
- Cajochen, C.; Zeitzer, J.M.; Czeisler, C.A.; Dijk, D.J. Dose-response relationship for light intensity and ocular and electroencephalographic correlates of human alertness. Behav. Brain Res. 2000, 115, 75–83. [Google Scholar] [CrossRef]
- Zeitzer, J.M.; Dijk, D.-J.; Kronauer, R.E.; Brown, E.N.; Czeisler, C.A. Sensitivity of the human circadian pacemaker to nocturnal light: Melatonin phase resetting and suppression. J. Physiol. 2000, 526, 695–702. [Google Scholar] [CrossRef]
- Nowozin, C.; Wahnschaffe, A.; Rodenbeck, A.; de Zeeuw, J.; Hädel, S.; Kozakov, R.; Schöpp, H.; Münch, M.; Kunz, D. Applying melanopic lux to measure biological light effects on melatonin suppression and subjective sleepiness. Curr. Alzheimer Res. 2017, 14, 1042–1052. [Google Scholar] [CrossRef]
- Prayag, A.S.; Najjar, R.P.; Gronfier, C. Melatonin suppression is exquisitely sensitive to light and primarily driven by melanopsin in humans. J. Pineal Res. 2019. [Google Scholar] [CrossRef]
- Rimmer, D.W.; Boivin, D.B.; Shanahan, T.L.; Kronauer, R.E.; Duffy, J.F.; Czeisler, C.A. Dynamic resetting of the human circadian pacemaker by intermittent bright light. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2000, 279, R1574–R1579. [Google Scholar] [CrossRef]
- Gronfier, C.; Wright, K.P.; Kronauer, R.E.; Jewett, M.E.; Czeisler, C.A. Efficacy of a single sequence of intermittent bright light pulses for delaying circadian phase in humans. Am. J. Physiol.-Endocrinol. Metab. 2004, 287, E174–E181. [Google Scholar] [CrossRef] [PubMed]
- Zeitzer, J.M.; Ruby, N.F.; Fisicaro, R.A.; Heller, H.C. Response of the human circadian system to millisecond flashes of light. PLoS ONE 2011, 6, e22078. [Google Scholar] [CrossRef]
- Najjar, R.P.; Zeitzer, J.M. Temporal integration of light flashes by the human circadian system. J. Clin. Invest. 2016, 126, 938–947. [Google Scholar] [CrossRef] [PubMed]
- Visser, E.K.; Beersma, D.G.M.; Daan, S. Melatonin suppression by light in humans is maximal when the nasal part of the retina is illuminated. J. Biol. Rhythms 1999, 14, 116–121. [Google Scholar] [CrossRef]
- Glickman, G.; Hanifin, J.P.; Rollag, M.D.; Wang, J.; Cooper, H.; Brainard, G.C. Inferior retinal light exposure is more effective than superior retinal exposure in suppressing melatonin in humans. J. Biol. Rhythms 2003, 18, 71–79. [Google Scholar] [CrossRef]
- Rüger, M.; Gordijn, M.C.M.; Beersma, D.G.M.; de Vries, B.; Daan, S. Nasal versus temporal illumination of the human retina: Effects on core body temperature, melatonin, and circadian phase. J. Biol. Rhythms 2005, 20, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Joyce, D.S.; Feigl, B.; Zele, A.J. Melanopsin-mediated post-illumination pupil response in the peripheral retina. J. Vis. 2016, 16, 5. [Google Scholar] [CrossRef] [PubMed]
- Hébert, M.; Martin, S.K.; Lee, C.; Eastman, C.I. The effects of prior light history on the suppression of melatonin by light in humans. J. Pineal Res. 2002, 33, 198–203. [Google Scholar] [CrossRef]
- Smith, K.A.; Schoen, M.W.; Czeisler, C.A. Adaptation of human pineal melatonin suppression by recent photic history. J. Clin. Endocrinol. Metab. 2004, 89, 3610–3614. [Google Scholar] [CrossRef]
- Jasser, S.A. Dim light adaptation attenuates acute melatonin suppression in humans. J. Biol. Rhythms 2006, 21, 394–404. [Google Scholar] [CrossRef]
- Chang, A.-M.; Scheer, F.A.J.L.; Czeisler, C.A. The human circadian system adapts to prior photic history. J. Physiol. 2011, 589, 1095–1102. [Google Scholar] [CrossRef]
- Chellappa, S.L.; Ly, J.Q.M.; Meyer, C.; Balteau, E.; Degueldre, C.; Luxen, A.; Phillips, C.; Cooper, H.M.; Vandewalle, G. Photic memory for executive brain responses. Proc. Natl. Acad. Sci. USA 2014, 111, 6087–6091. [Google Scholar] [CrossRef]
- Münch, M.; Nowozin, C.; Regente, J.; Bes, F.; De Zeeuw, J.; Hädel, S.; Wahnschaffe, A.; Kunz, D. Blue-enriched morning light as a countermeasure to light at the wrong time: Effects on cognition, sleepiness, sleep, and circadian phase. Neuropsychobiology 2016, 74, 207–218. [Google Scholar] [CrossRef]
- Lockley, S.W.; Evans, E.E.; Scheer, F.; Brainard, G.C.; Czeisler, C.A.; Aeschbach, D. Short-wavelength sensitivity for the direct effects of light on alertness, vigilance, and the waking electroencephalogram in humans. Sleep 2006, 29, 161–168. [Google Scholar] [PubMed]
- Phipps-Nelson, J.; Redman, J.R.; Schlangen, L.J.M.; Rajaratnam, S.M.W. Blue light exposure reduces objective measures of sleepiness during prolonged nighttime performance testing. Chronobiol. Int. 2009, 26, 891–912. [Google Scholar] [CrossRef] [PubMed]
- Figueiro, M.G.; Bierman, A.; Plitnick, B.; Rea, M.S. Preliminary evidence that both blue and red light can induce alertness at night. BMC Neurosci. 2009, 10, 105. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.A.; St. Hilaire, M.A.; Lockley, S.W. The effects of spectral tuning of evening ambient light on melatonin suppression, alertness and sleep. Physiol. Behav. 2017, 177, 221–229. [Google Scholar] [CrossRef]
- Wams, E.J.; Woelders, T.; Marring, I.; van Rosmalen, L.; Beersma, D.G.M.; Gordijn, M.C.M.; Hut, R.A. Linking light exposure and subsequent sleep: A field polysomnography study in humans. Sleep 2017, 40, zsx165. [Google Scholar] [CrossRef]
- Nowozin, C.; Wahnschaffe, A.; De Zeeuw, J.; Papakonstantinou, A.; Bes, F.; Münch, M.; Kunz, D. Dynamics of daytime light exposure impacts on sleep architecture in a naturalistic setting. In Chronobiology: Society for Light Treatment and Biological Rhythms, Proceedings of the 30th Annual Meeting, Groningen, The Netherlands, 21–24 June 2018; Neuropsychobiology; Karger: Basel, Switzerland, 2017; Volume 76, p. 25. [Google Scholar]
- van der Lely, S.; Frey, S.; Garbazza, C.; Wirz-Justice, A.; Jenni, O.G.; Steiner, R.; Wolf, S.; Cajochen, C.; Bromundt, V.; Schmidt, C. Blue blocker glasses as a countermeasure for alerting effects of evening light-emitting diode screen exposure in male teenagers. J. Adolesc. Health 2015, 56, 113–119. [Google Scholar] [CrossRef]
- Chang, A.-M.; Aeschbach, D.; Duffy, J.F.; Czeisler, C.A. Evening use of light-emitting eReaders negatively affects sleep, circadian timing, and next-morning alertness. Proc. Natl. Acad. Sci. USA 2015, 112, 1232–1237. [Google Scholar] [CrossRef]
- Münch, M.; Kobialka, S.; Steiner, R.; Oelhafen, P.; Wirz-Justice, A.; Cajochen, C. Wavelength-dependent effects of evening light exposure on sleep architecture and sleep EEG power density in men. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2006, 290, R1421–R1428. [Google Scholar] [CrossRef] [PubMed]
- Chellappa, S.L.; Steiner, R.; Oelhafen, P.; Lang, D.; Götz, T.; Krebs, J.; Cajochen, C. Acute exposure to evening blue-enriched light impacts on human sleep. J. Sleep Res. 2013, 22, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Ostrin, L.A.; Abbott, K.S.; Queener, H.M. Attenuation of short wavelengths alters sleep and the ipRGC pupil response. Ophthalmic Physiol. Opt. 2017, 37, 440–450. [Google Scholar] [CrossRef] [PubMed]
- van der Meijden, W.P.; te Lindert, B.H.W.; Ramautar, J.R.; Wei, Y.; Coppens, J.E.; Kamermans, M.; Cajochen, C.; Bourgin, P.; Van Someren, E.J.W. Sustained effects of prior red light on pupil diameter and vigilance during subsequent darkness. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180989. [Google Scholar] [CrossRef]
- Obayashi, K.; Saeki, K.; Kurumatani, N. Bedroom light exposure at night and the incidence of depressive symptoms: A longitudinal study of the HEIJO-KYO cohort. Am. J. Epidemiol. 2018, 187, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Zeitzer, J.M.; Fisicaro, R.A.; Ruby, N.F.; Heller, H.C. Millisecond flashes of light phase delay the human circadian clock during sleep. J. Biol. Rhythms 2014, 29, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.R.; Joo, E.Y.; Koo, D.L.; Hong, S.B. Let there be no light: The effect of bedside light on sleep quality and background electroencephalographic rhythms. Sleep Med. 2013, 14, 1422–1425. [Google Scholar] [CrossRef] [PubMed]
- Arendt, J. Melatonin and the pineal gland: Influence on mammalian seasonal and circadian physiology. Rev. Reprod. 1998, 3, 13–22. [Google Scholar] [CrossRef]
- Dacey, D.M.; Liao, H.-W.; Peterson, B.B.; Robinson, F.R.; Smith, V.C.; Pokorny, J.; Yau, K.-W.; Gamlin, P.D. Melanopsin-expressiong ganglion cells in primate retina signal colour and irradiance and project to the LGN. Nature 2005, 433, 749–754. [Google Scholar] [CrossRef]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef]
- Boivin, D.B.; Duffy, J.F.; Kronauer, R.E.; Czeisler, C.A. Dose-response relationships for resetting of human circadian clock by light. Nature 1996, 379, 540. [Google Scholar] [CrossRef]
- Brainard, G.C.; Sliney, D.; Hanifin, J.P.; Glickman, G.; Byrne, B.; Greeson, J.M.; Jasser, S.; Gerner, E.; Rollag, M.D. Sensitivity of the human Circadian system to short-wavelength (420-nm) Light. J. Biol. Rhythms 2008, 23, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Lucas, R.J.; Peirson, S.N.; Berson, D.M.; Brown, T.M.; Cooper, H.M.; Czeisler, C.A.; Figueiro, M.G.; Gamlin, P.D.; Lockley, S.W.; O’Hagan, J.B.; et al. Measuring and using light in the melanopsin age. Trends Neurosci. 2014, 37, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Commission International de l’Eclairage. CIE System for Metrology of Optical Radiation for ipRGC-Influenced Responses to Light; CIE S 026/E:2018; CIE Central Bureau: Vienna, Austria, 2018. [Google Scholar]
- Gronfier, C.; Wright, K.P.; Kronauer, R.E.; Czeisler, C.A. Entrainment of the human circadian pacemaker to longer-than-24-h days. Proc. Natl. Acad. Sci. USA 2007, 104, 9081–9086. [Google Scholar] [CrossRef]
- Revell, V.L.; Molina, T.A.; Eastman, C.I. Human phase response curve to intermittent blue light using a commercially available device: Human blue light phase response curve. J. Physiol. 2012, 590, 4859–4868. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.Y.; Dunn, F.A.; Berson, D.M. Photoreceptor adaptation in intrinsically photosensitive retinal ganglion cells. Neuron 2005, 48, 1001–1010. [Google Scholar] [CrossRef]
- Segal, A.Y.; Sletten, T.L.; Flynn-Evans, E.E.; Lockley, S.W.; Rajaratnam, S.M. Daytime exposure to short-and medium-wavelength light did not improve alertness and neurobehavioral performance. J. Biol. Rhythms 2016, 31, 470–482. [Google Scholar] [CrossRef]
- Lupi, D.; Oster, H.; Thompson, S.; Foster, R.G. The acute light-induction of sleep is mediated by OPN4-based photoreception. Nat. Neurosci. 2008, 11, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.W.; Hannibal, J.; Hagiwara, G.; Colas, D.; Ruppert, E.; Ruby, N.F.; Heller, H.C.; Franken, P.; Bourgin, P. Melanopsin as a sleep modulator: Circadian gating of the direct effects of light on sleep and altered sleep homeostasis in Opn4−/− mice. PLoS Biol. 2009, 7, e1000125. [Google Scholar] [CrossRef]
- Rahman, S.A.; Shapiro, C.M.; Wang, F.; Ainlay, H.; Kazmi, S.; Brown, T.J.; Casper, R.F. Effects of filtering visual short wavelengths during nocturnal shiftwork on sleep and performance. Chronobiol. Int. 2013, 30, 951–962. [Google Scholar] [CrossRef]
- Dijk, D.J.; Visscher, C.; Bloem, G.; Beersma, D.; Daan, S. Reduction of human sleep duration after bright light exposure in the morning. Neurosci. Lett. 1987, 73, 181–186. [Google Scholar] [CrossRef]
- Kobayashi, R.; Kohsaka, M.; Fukuda, N.; Sakakibara, S.; Honma, H.; Koyama, T. Effects of morning bright light on sleep in healthy elderly women. Psychiatry Clin. Neurosci. 1999, 53, 237–238. [Google Scholar] [CrossRef] [PubMed]
- Sack, R.L.; Lewy, A.; Miller, L.; Singer, C. Effects of morning versus evening bright light exposure on REM latency. Biol. Psychiatry 1986, 21, 410–413. [Google Scholar] [CrossRef]
- Dijk, D.J.; Beersma, D.G.; Daan, S.; Lewy, A.J. Bright morning light advances the human circadian system without affecting NREM sleep homeostasis. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1989, 256, R106–R111. [Google Scholar] [CrossRef] [PubMed]
- Carrier, J.; Dumont, M. Sleep propensity and sleep architecture after bright light exposure at three different times of day. J. Sleep Res. 1995, 4, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Najjar, R.P.; Wolf, L.; Taillard, J.; Schlangen, L.J.M.; Salam, A.; Cajochen, C.; Gronfier, C. Chronic artificial blue-enriched white light is an effective countermeasure to delayed circadian phase and neurobehavioral decrements. PLoS ONE 2014, 9, e102827. [Google Scholar] [CrossRef]
- Gordijn, M.C.M.; Beersma, D.G.M.; Korte, H.J.; Hoofdakker, R.H. Effects of light exposure and sleep displacement on dim light melatonin onset. J. Sleep Res. 1999, 8, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Komada, Y.; Tanaka, H.; Yamamoto, Y.; Shirakawa, S.; Yamazaki, K. Effects of bright light pre-exposure on sleep onset process. Psychiatry Clin. Neurosci. 2000, 54, 365–366. [Google Scholar] [CrossRef]
- Santhi, N.; Thorne, H.C.; van der Veen, D.R.; Johnsen, S.; Mills, S.L.; Hommes, V.; Schlangen, L.J.M.; Archer, S.N.; Dijk, D.-J. The spectral composition of evening light and individual differences in the suppression of melatonin and delay of sleep in humans: Artificial evening light suppresses melatonin and delays sleep. J. Pineal Res. 2012, 53, 47–59. [Google Scholar] [CrossRef]
- Touitou, Y.; Touitou, D.; Reinberg, A. Disruption of adolescents’ circadian clock: The vicious circle of media use, exposure to light at night, sleep loss and risk behaviors. J. Physiol.-Paris 2016, 110, 467–479. [Google Scholar] [CrossRef]
- Figueiro, M.; Rea, M. Office lighting and personal light exposures in two seasons: Impact on sleep and mood. Light. Res. Technol. 2016, 48, 352–364. [Google Scholar] [CrossRef]
- Figueiro, M.G.; Wood, B.; Plitnick, B.; Rea, M.S. The impact of light from computer monitors on melatonin levels in college students. Biog. Amines 2011, 25, 106–116. [Google Scholar]
- Cajochen, C.; Frey, S.; Anders, D.; Spati, J.; Bues, M.; Pross, A.; Mager, R.; Wirz-Justice, A.; Stefani, O. Evening exposure to a light-emitting diodes (LED)-backlit computer screen affects circadian physiology and cognitive performance. J. Appl. Physiol. 2011, 110, 1432–1438. [Google Scholar] [CrossRef] [PubMed]
- Hatori, M.; Gronfier, C.; Van Gelder, R.N.; Bernstein, P.S.; Carreras, J.; Panda, S.; Marks, F.; Sliney, D.; Hunt, C.E.; Hirota, T.; et al. Global rise of potential health hazards caused by blue light-induced circadian disruption in modern aging societies. NPJ Aging Mech. Dis. 2017, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Hätönen, T.; Alila-Johansson, A.; Mustanoja, S.; Laakso, M.-L. Suppression of melatonin by 2000-lux light in humans with closed eyelids. Biol. Psychiatry 1999, 46, 827–831. [Google Scholar] [CrossRef]
- Akacem, L.D.; Wright, K.P.; LeBourgeois, M.K. Sensitivity of the circadian system to evening bright light in preschool-age children. Physiol. Rep. 2018, 6, e13617. [Google Scholar] [CrossRef]
- Pilorz, V.; Tam, S.K.E.; Hughes, S.; Pothecary, C.A.; Jagannath, A.; Hankins, M.W.; Bannerman, D.M.; Lightman, S.L.; Vyazovskiy, V.V.; Nolan, P.M.; et al. melanopsin regulates both sleep-promoting and arousal-promoting responses to light. PLoS Biol. 2016, 14, e1002482. [Google Scholar] [CrossRef]
- Allen, A.E.; Hazelhoff, E.M.; Martial, F.P.; Cajochen, C.; Lucas, R.J. Exploiting metamerism to regulate the impact of a visual display on alertness and melatonin suppression independent of visual appearance. Sleep 2018, 41, zsy100. [Google Scholar] [CrossRef]
- Cajochen, C.; Chellappa, S.L. Commentary: Melanopsin regulates both sleep-promoting and arousal-promoting responses to light. Front. Neural Circuits 2016, 10, 94. [Google Scholar] [CrossRef]
- Rahman, S.A.; St Hilaire, M.A.; Gronfier, C.; Chang, A.-M.; Santhi, N.; Czeisler, C.A.; Klerman, E.B.; Lockley, S.W. Functional decoupling of melatonin suppression and circadian phase resetting in humans. J. Physiol. 2018, 596, 2147–2157. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Section | Key References |
|---|---|
| Differences in spectral sensitivity to light | Spectral sensitivity of (i) melatonin suppression by light (Brainard et al. [27]; Thapan et al. [28]; Cajochen et al. [8]; Najjar et al. [29]) (ii) circadian phase-shifting (Lockley et al. [30]; Revell et al. [31]) |
| Impact of duration of light exposure | Duration response curves for (i) melatonin suppression and circadian phase-shifting (Chang et al. [32]; Dewan et al. [33]) (ii) EEG, distal to proximal temperature gradient (DPG), heart rate (Prayag et al. [34]) |
| Effects of timing of light exposure | Phase response curves (Khalsa et al. [6]; Rüger et al. [35]; St Hilaire et al. [36]) |
| Effects of light intensity/irradiance | Illuminance response curves for (i) subjective alertness (KSS), EEG (5–9 Hz), slow-eye movements (Cajochen et al. [37]) (ii) melatonin suppression and circadian phase-shifting (Zeitzer et al. [38]) (iii) melatonin suppression as a function of melanopic illuminance (Nowozin et al. [39]; Prayag et al. [40]) |
| Effects of patterns of light exposure | (i) Intermittent bright light and circadian phase-shifting (Rimmer et al. [41]; Gronfier et al. [42]) (ii) Response to milliseconds flashes of light (Zeitzer et al. [43]; Zeitzer & Najjar, [44]) |
| Impact of spatial distribution of light exposure | (i) melatonin suppression (Visser et al. [45]; Glickman et al. [46]; Rüger et al. [47]) (ii) pupillary light reflex (Joyce et al. [48]) |
| Effects of prior light history | Influence of light history on non-visual responses (Hébert et al. [49]; Smith et al. [50]; Jasser et al. [51]; Chang et al. [52]; Chellappa et al. [53]; Münch et al. [54]) |
| Impact of light on cortical activity during wakefulness | Influence of bright light and short-wavelength light on the waking EEG (Badia et al. [12]; Lockley et al. [55]; Phipps-Nelson et al. [56]; Figueiro et al. [57]; Rahman et al. [11,58]) |
| Impact of light on sleep | (i) Impact of brighter light during daytime increases SWS accumulation during the following night (Wams et al. [59]; Nowozin et al. [60]) (ii) Effects of short-wavelength light, including from LED devices, before bedtime on sleepiness (e.g., van der Lely et al. [61]), sleep onset (e.g., Chang et al. [62]), REM sleep, SWS and sleep architecture (Münch et al. [63]; Chellappa et al. [64]) (iii) Impact of longer wavelengths light prior to sleep on sleep promotion (Münch et al. [54]; Ostrin et al. [65]; van der Meijden et al [66]) (iv) Impact of light during sleep on melatonin suppression (Obayashi et al. [67]; Zeitzer et al. [68]) and SWS (Cho et al. [69]) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prayag, A.S.; Münch, M.; Aeschbach, D.; Chellappa, S.L.; Gronfier, C. Light Modulation of Human Clocks, Wake, and Sleep. Clocks & Sleep 2019, 1, 193-208. https://doi.org/10.3390/clockssleep1010017
Prayag AS, Münch M, Aeschbach D, Chellappa SL, Gronfier C. Light Modulation of Human Clocks, Wake, and Sleep. Clocks & Sleep. 2019; 1(1):193-208. https://doi.org/10.3390/clockssleep1010017
Chicago/Turabian StylePrayag, Abhishek S., Mirjam Münch, Daniel Aeschbach, Sarah L. Chellappa, and Claude Gronfier. 2019. "Light Modulation of Human Clocks, Wake, and Sleep" Clocks & Sleep 1, no. 1: 193-208. https://doi.org/10.3390/clockssleep1010017
APA StylePrayag, A. S., Münch, M., Aeschbach, D., Chellappa, S. L., & Gronfier, C. (2019). Light Modulation of Human Clocks, Wake, and Sleep. Clocks & Sleep, 1(1), 193-208. https://doi.org/10.3390/clockssleep1010017